
Sf-FGFR and Sf-SR-C Are Not the Receptors for Vip3Aa to Exert Insecticidal Toxicity in Spodoptera frugiperda



 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
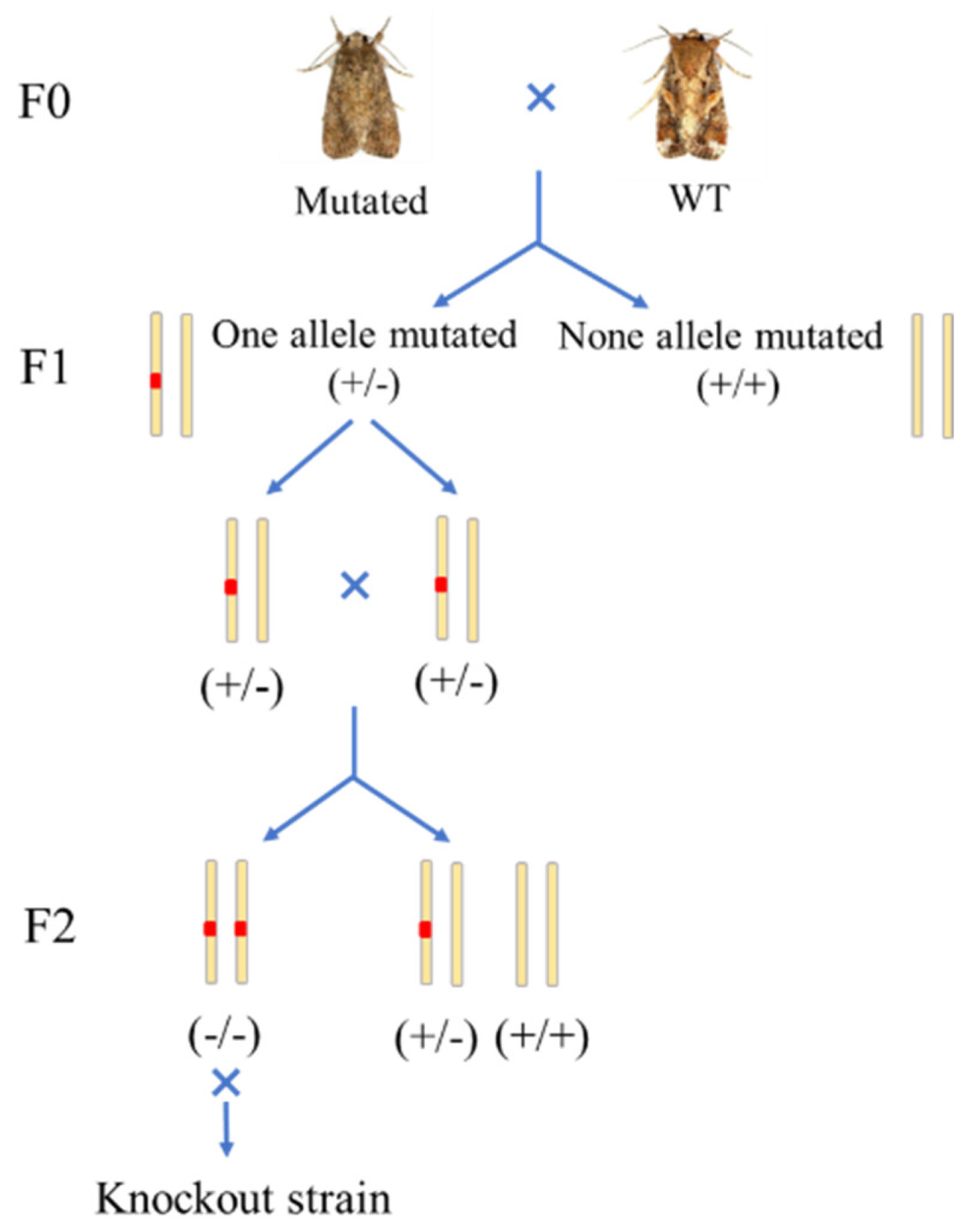
2.1. Insect Strains and Rearing
2.2. SgRNAs Design and Synthesis
2.3. Collection and Injection of S. frugiperda Eggs
2.4. Genomic DNA Extraction and Mutagenesis Detection
2.5. Bt Toxins and Bioassays
3. Results
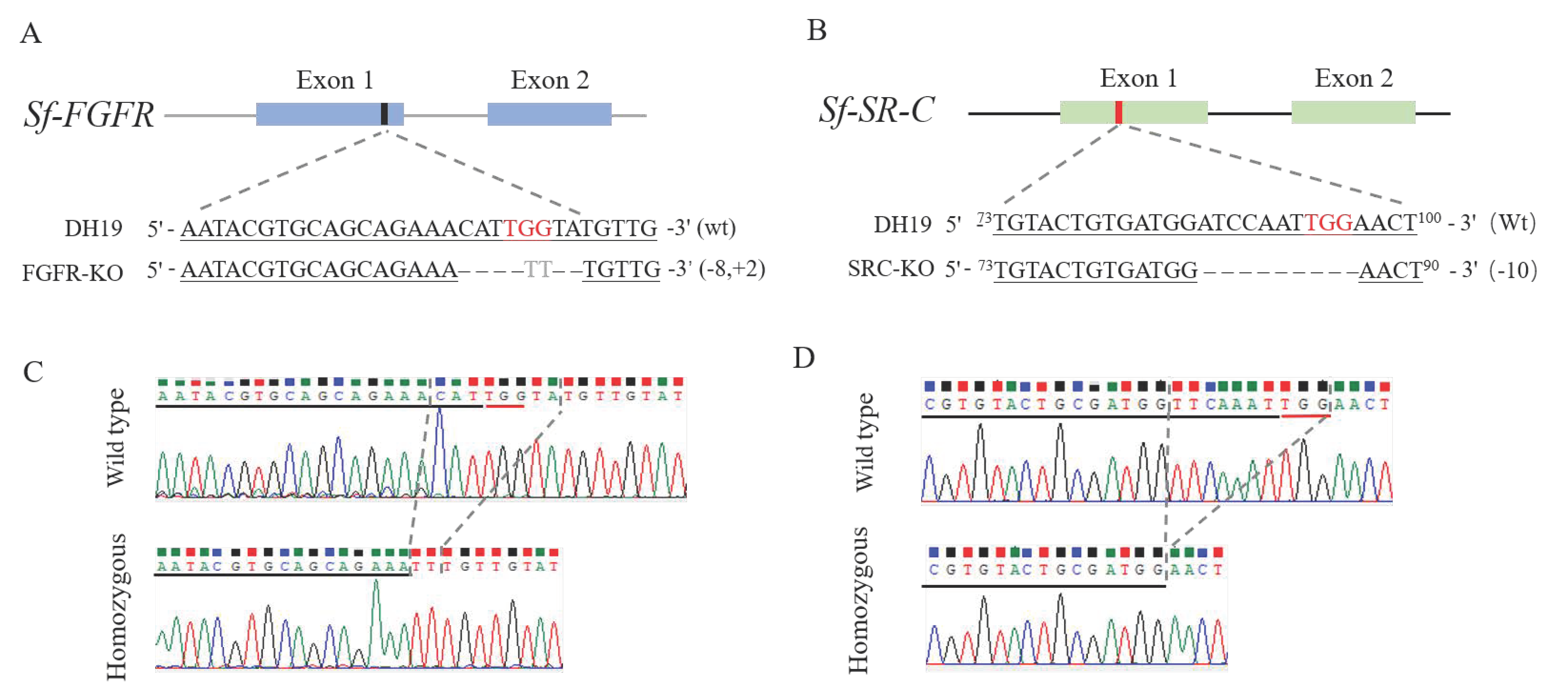
3.1. Establishment of the Sf-FGFR Knockout Strain and Its Mutation Types
3.2. Establishment of Sf-SR-C Knockout Strain and Its Mutation Types
3.3. Susceptibility to Vip3Aa Toxins in FGFR-KO and SRC-KO
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Hardke, J.T.; Lorenz, G.M.; Leonard, B.R. Fall Armyworm (Lepidoptera: Noctuidae) Ecology in Southeastern Cotton. Int. J. Pest Manag. 2015, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Sparks, A.N. A Review of the Biology of the Fall Armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a New Alien Invasive Pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Sisodiya, D.B.; Raghunandan, B.L.; Bhatt, N.A.; Verma, H.S.; Shewale, C.P.; Timbadiya, B.G.; Borad, P.K. The fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae); First report of new invasive pest in maize fields of Gujarat, India. J. Entomol. Zool. Stud. 2018, 6, 2089–2091. [Google Scholar]
- FAO. First Detection of All Armyworm on the Border of Thailand; FAO: Rome, Italy, 2018. [Google Scholar]
- FAO. First Detection Report of the Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) on Maize in MYANMAR; FAO: Rome, Italy, 2019. [Google Scholar]
- Guo, J.F.; He, K.L.; Wang, Z.Y. Biological characteristics, trend of fall armyworm Spodoptera frugiperda, and the strategy for management of the pest. Chin. J. Appl. Entomol. 2019, 56, 361–369. [Google Scholar]
- Jiang, Y.Y.; Liu, J.; Xie, M.C.; Li, Y.H.; Yang, J.J.; Zhang, M.L.; Qiu, K. Observation on law of diffusion damage of Spodoptera frugiperda in China in 2019. Plant Protect. 2019, 45, 10–19. [Google Scholar]
- Liao, Y.L.; Li, C.Y.; Huang, S.H.; Pan, Z.P.; Yang, B.; Chen, J.R.; Wang, L.J.; Liu, W.L.; Zhang, Y.P. Survey on the prevalence and damage of Spodoptera furgiperda first invasive in Guangdong. J. Econ. Entomol. 2019, 41, 497–502. [Google Scholar]
- Wu, Q.; Jiang, Y.Y.; Wu, K. Analysis of migration routes of the fall armyworm Spodoptera frugiperda (J. E. Smith) from Myanmar to China. Plant Protect. 2019, 45, 1–6. [Google Scholar]
- Stokstad, E. New crop pest takes Africa at lightning speed. Science 2017, 356, 473–474. [Google Scholar] [CrossRef]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2017: Biotech Crop Adoption Surges as Economic Benefits Accumulate in 22 Years; ISAAA Brief No. 53.; ISAAA: Ithaca, NY, USA, 2017. [Google Scholar]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Huang, F.; Qureshi, J.A.; Head, G.P.; Price, P.A.; Levy, R.; Yang, F.; Niu, Y. Frequency of Bacillus thuringiensis Cry1A.105 resistance alleles in field populations of the fall armyworm, Spodoptera frugiperda, in Louisiana and Florida. Crop Prot. 2016, 83, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Monnerat, R.; Martins, E.; Macedo, C.; Queiroz, P.; Praça, L.; Soares, C.M.; Moreira, H.; Grisi, I.; Silva, J.; Soberon, M.; et al. Evidence of field-evolved resistance of Spodoptera frugiperda to Bt corn expressing Cry1F in Brazil that is still sensitive to modified Bt toxins. PLoS ONE 2015, 10, e0119544. [Google Scholar] [CrossRef] [Green Version]
- Chandrasena, D.I.; Signorini, A.M.; Abratti, G.; Storer, N.P.; Olaciregui, M.L.; Alves, A.P.; Pilcher, C.D. Characterization of field-evolved resistance to Bacillus thuringiensis-derived Cry1F δ-endotoxin in Spodoptera frugiperda populations from Argentina. Pest Manag. Sci. 2018, 74, 746–754. [Google Scholar] [CrossRef] [Green Version]
- Núñez-Ramírez, R.; Huesa, J.; Bel, Y.; Ferré, J.; Casino, P.; Arias-Palomo, E. Molecular architecture and activation of the insecticidal protein Vip3Aa from Bacillus thuringiensis. Nat. Commun. 2020, 11, 3974. [Google Scholar] [CrossRef]
- Estruch, J.J.; Warren, G.W.; Mullins, M.A.; Nye, G.J.; Craig, J.A.; Koziel, M.G. Vip3A, a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects. Proc. Natl. Acad. Sci. USA 1996, 93, 5389–5394. [Google Scholar] [CrossRef] [Green Version]
- Sena, J.A.; Hernández-Rodríguez, C.S.; Ferré, J. Interaction of Bacillus thuringiensis Cry1 and Vip3A proteins with Spodoptera frugiperda midgut binding sites. Appl. Environ. Microbiol. 2018, 75, 2236–2237. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Huang, F.N.; Qureshi, J.A.; Leonard, B.R.; Niu, Y.; Zhang, L.P.; Wangila, D.S. Susceptibility of Louisiana and Florida populations of Spodoptera frugiperda (Lepidoptera:Noctuidae) to transgenic Agrisure (R) Viptera (TM) 3111 corn. Crop Prot. 2013, 50, 37–39. [Google Scholar] [CrossRef]
- Yang, F.; Kerns, D.L.; Head, G.; Brown, S.; Huang, F.N. Susceptibility of Cry1F-maize resistant, heterozygous, and susceptible Spodoptera frugiperda to Bt proteins used in the transgenic cotton. Crop Prot. 2017, 98, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Kerns, D.L.; Head, G.P.; Price, P.; Huang, F. Cross-resistance to purified Bt proteins, Bt corn and Bt cotton in a Cry2Ab2-corn resistant strain of Spodoptera frugiperda. Pest Manag. Sci. 2017, 73, 2495–2503. [Google Scholar] [CrossRef]
- Gomis-Cebolla, J.; Wang, Y.; Quan, Y.; He, K.; Walsh, T.; James, B.; Downes, S.; Kain, W.; Wang, P.; Leonard, K.; et al. Analysis of cross-resistance to Vip3 proteins in eight insect colonies, from four insect species, selected for resistance to Bacillus thuringiensis insecticidal proteins. J. Invertebr. Pathol. 2018, 155, 64–70. [Google Scholar] [CrossRef]
- Chakroun, M.; Banyuls, N.; Bel, Y.; Escriche, B.; Ferré, J. Bacterial Vegetative Insecticidal Proteins (Vip) from Entomopathogenic Bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 329–350. [Google Scholar] [CrossRef] [Green Version]
- Bernardi, O.; Bernardi, D.; Ribeiro, R.S.; Okuma, D.M.; Salmeron, E.; Fatoretto, J.; Medeiros, F.C.; Burd, T.; Omoto, C. Frequency of resistance to Vip3Aa20 toxin from Bacillus thuringiensis in Spodoptera frugiperda (Lepidoptera: Noctuidae) populations in Brazil. Crop Prot. 2015, 76, 7–14. [Google Scholar] [CrossRef]
- Yang, F.; Williams, J.; Porter, P.; Huang, F.; Kerns, D.L. F2 screen for resistance to Bacillus thuringiensis Vip3Aa51 protein in field populations of Spodoptera frugiperda (Lepidoptera: Noctuidae) from Texas, USA. Crop Prot. 2019, 126, 104915. [Google Scholar] [CrossRef]
- Fitt, G.P. An Australian approach to IPM in cotton: Integrating new technologies to minimise insecticide dependence. Crop Prot. 2000, 19, 793–800. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Mota-Sanchez, D.; Whalon, M.E.; Hollingworth, R.M.; Carrière, Y. Defning terms for proactive management of resistance to Bt crops and pesticides. J. Econ. Entomol. 2014, 107, 496–507. [Google Scholar] [CrossRef]
- Jiang, K.; Hou, X.Y.; Tan, T.T.; Cao, Z.L.; Mei, S.Q.; Yan, B.; Chang, J.; Han, L.; Zhao, D.; Cai, J. Scavenger receptor-C acts as a receptor for Bacillus thuringiensis vegetative insecticidal protein Vip3Aa and mediates the internalization of Vip3Aa via endocytosis. PLoS Pathog. 2018, 14, e1007347. [Google Scholar] [CrossRef]
- Jiang, K.; Hou, X.; Han, L.; Tan, T.; Cao, Z.; Cai, J. Fibroblast Growth Factor Receptor, a Novel Receptor for Vegetative Insecticidal Protein Vip3Aa. Toxins 2018, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Tao, J.; Li, Q.; Cheng, Y.; Sun, X.; Wu, K.; Xiao, Y. Genome editing of the SfABCC2 gene confers resistance to Cry1F toxin from Bacillus thuringiensis in Spodoptera frugiperda. J. Integr. Agric. 2019, 18, 2–7. [Google Scholar] [CrossRef]
- Li, G.P.; Sun, X.X.; Jiang, Y.Y.; Wu, K.M.; Feng, H.Q. Susceptibility evaluation of invaded Spodoptera frugiperda population in Yunan province to five Bt proteins. Plant Protect. 2019, 45, 15–20. [Google Scholar]
- Xie, S.; Shen, B.; Zhang, C.; Huang, X.; Zhang, Y. sgRNAcas9: A software package for designing CRISPR sgRNA and evaluating potential off-target cleavage sites. PLoS ONE 2014, 9, e100448. [Google Scholar] [CrossRef] [PubMed]
- Wesche, J.; Haglund, K.; Haugsten, E.M. Fibroblast growth factors and their receptors in cancer. Biochem. J. 2011, 437, 199–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, A.M. Scavenger receptors in innate immunity. Curr. Opin. Immunol. 1996, 8, 208. [Google Scholar] [CrossRef]
- Lee, M.K.; Walters, F.S.; Hart, H.; Palekar, N.; Chen, J.S. The mode of action of the Bacillus thuringiensis vegetative insecticidal protein Vip3A differs from that of Cry1Ab delta-endotoxin. Appl. Environ. Microbiol. 2003, 69, 4648–4657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.K.; Miles, P.; Chen, J.S. Brush border membrane binding properties of Bacillus thuringiensis Vip3A toxin to Heliothis virescens and Helicoverpa zea midguts. Biochem. Biophys. Res. Commun. 2006, 339, 1043–1047. [Google Scholar] [CrossRef] [PubMed]
- Syed, T.; Askari, M.; Meng, Z.; Li, Y.; Abid, M.A.; Wei, Y.; Guo, S.; Liang, C.; Zhang, R. Current Insights on Vegetative Insecticidal Proteins (Vip) as Next Generation Pest Killers. Toxins 2020, 12, 522. [Google Scholar] [CrossRef]
- Dai, S.; Zhou, Z.; Chen, Z.; Xu, G.; Chen, Y. Fibroblast Growth Factor Receptors (FGFRs): Structures and Small Molecule Inhibitors. Cells 2019, 8, 614. [Google Scholar] [CrossRef] [Green Version]
- Katoh, M. Therapeutics Targeting FGF Signaling Network in Human Diseases. Trends Pharmacol. Sci. 2016, 37, 1081–1096. [Google Scholar] [CrossRef]
- Turner, N.; Grose, R. Fibroblast growth factor signalling: From development to cancer. Nat. Rev. Cancer 2010, 10, 116–129. [Google Scholar] [CrossRef]
- Bel, Y.; Jakubowska, A.K.; Costa, J.; Herrero, S.; Escriche, B. Comprehensive analysis of gene expression profiles of the beet armyworm Spodoptera exigua larvae challenged with Bacillus thuringiensis Vip3Aa toxin. PLoS ONE 2013, 8, e81927. [Google Scholar] [CrossRef] [Green Version]
- PrabhuDas, M.R.; Baldwin, C.L.; Bollyky, P.L.; Bowdish, D.M.E.; Drickamer, K.; Febbraio, M.; Herz, J.; Kobzik, L.; Krieger, M.; Loike, J.; et al. A Consensus Definitive Classification of Scavenger Receptors and Their Roles in Health and Disease. J. Immunol. 2017, 198, 3775–3789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canton, J.; Neculai, D.; Grinstein, S. Scavenger receptors in homeostasis and immunity. Nat. Rev. Immunol. 2013, 13, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Pearson, A.; Lux, A.; Krieger, M. Expression cloning of dSR-CI, a class C macrophage-specific scavenger receptor from Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1995, 92, 4056–4060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.C.; Shi, X.Z.; Yang, H.T.; Sun, J.J.; Xu, L.; Wang, X.W.; Zhao, X.F.; Wang, J.X. Scavenger Receptor C Mediates Phagocytosis of White Spot Syndrome Virus and Restricts Virus Proliferation in Shrimp. PLoS Pathog. 2016, 12, e1006127. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Guo, C.; Fisher, P.B.; Subjeck, J.R.; Wang, X.Y. Scavenger Receptors: Emerging Roles in Cancer Biology and Immunology. Adv. Cancer Res. 2015, 128, 309–364. [Google Scholar]
- Zhu, X.D.; Zhuang, Y.; Ben, J.J.; Qian, L.L.; Huang, H.P.; Bai, H.; Sha, J.H.; He, Z.G.; Chen, Q. Caveolae-dependent endocytosis is required for class A macrophage scavenger receptor-mediated apoptosis in macrophages. J. Biol. Chem. 2011, 286, 8231–8239. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Yang, M.; Yuen, P.M.; Chik, K.W.; Li, C.K.; Shing, M.M.; Lam, H.K.; Fok, T.F. Thrombospondin-1 induces apoptosis in primary leukemia and cell lines mediated by CD36 and Caspase-3. Int. J. Mol. Med. 2003, 12, 995–1001. [Google Scholar] [CrossRef]
- Murphy, J.E.; Tacon, D.; Tedbury, P.R.; Hadden, J.M.; Knowling, S.; Sawamura, T.; Peckham, M.; Phillips, S.E.; Walker, J.H.; Ponnambalam, S. LOX-1 scavenger receptor mediates calcium-dependent recognition of phosphatidylserine and apoptotic cells. Biochem. J. 2006, 393, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Oka, K.; Sawamura, T.; Kikuta, K.; Itokawa, S.; Kume, N.; Kita, T.; Masaki, T. Lectin-like oxidized low-density lipoprotein receptor 1 mediates phagocytosis of aged/apoptotic cells in endothelial cells. Proc. Natl. Acad. Sci. USA 1998, 95, 9535–9540. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Mei, S.Q.; Wang, T.T.; Pan, J.H.; Chen, Y.H.; Cai, J. Vip3Aa induces apoptosis in cultured Spodoptera frugiperda (Sf9) cells. Toxicon 2016, 120, 49–56. [Google Scholar] [CrossRef]
- Hou, X.; Han, L.; An, B.; Zhang, Y.; Cao, Z.; Zhan, Y.; Cai, X.; Yan, B.; Cai, J. Mitochondria and Lysosomes Participate in Vip3Aa-Induced Spodoptera frugiperda Sf9 Cell Apoptosis. Toxins 2020, 12, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurat-Fuentes, J.L.; Heckel, D.G.; Ferré, J. Mechanisms of Resistance to Insecticidal Proteins from Bacillus thuringiensis. Annu. Rev. Entomol. 2021, 66, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Gahan, L.J.; Gould, F.; Heckel, D.G. Identification of a gene associated with Bt resistance in Heliothis virescens. Science 2001, 293, 857–860. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, H.; Wang, H.; Zhao, S.; Zuo, Y.; Yang, Y.; Wu, Y. Functional validation of cadherin as a receptor of Bt toxin Cry1Ac in Helicoverpa armigera utilizing the CRISPR/Cas9 system. Insect Biochem. Mol. Biol. 2016, 76, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wu, K.; Liang, G.; Guo, Y. Gene cloning and expression of cadherin in midgut of Helicoverpa armigera and its Cry1A binding region. Sci. China C Life Sci. 2005, 48, 346–356. [Google Scholar] [CrossRef]
- Qiu, L.; Hou, L.; Zhang, B.; Liu, L.; Li, B.; Deng, P.; Ma, W.; Wang, X.; Fabrick, J.A.; Chen, L.; et al. Cadherin is involved in the action of Bacillus thuringiensis toxins Cry1Ac and Cry2Aa in the beet armyworm, Spodoptera exigua. J. Invertebr. Pathol. 2015, 127, 47–53. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, J.; Xiao, Y.; Yang, Y.; Liu, C.; Peng, R.; Yang, Y.; Bravo, A.; Soberón, M.; Liu, K. The cadherin Cry1Ac binding-region is necessary for the cooperative effect with ABCC2 transporter enhancing insecticidal activity of Bacillus thuringiensis Cry1Ac toxin. Toxins 2019, 11, 538. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kain, W.; Wang, P. Bacillus thuringiensis Cry1A toxins exert toxicity by multiple pathways in insects. Insect Biochem. Mol. Biol. 2018, 102, 59–66. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
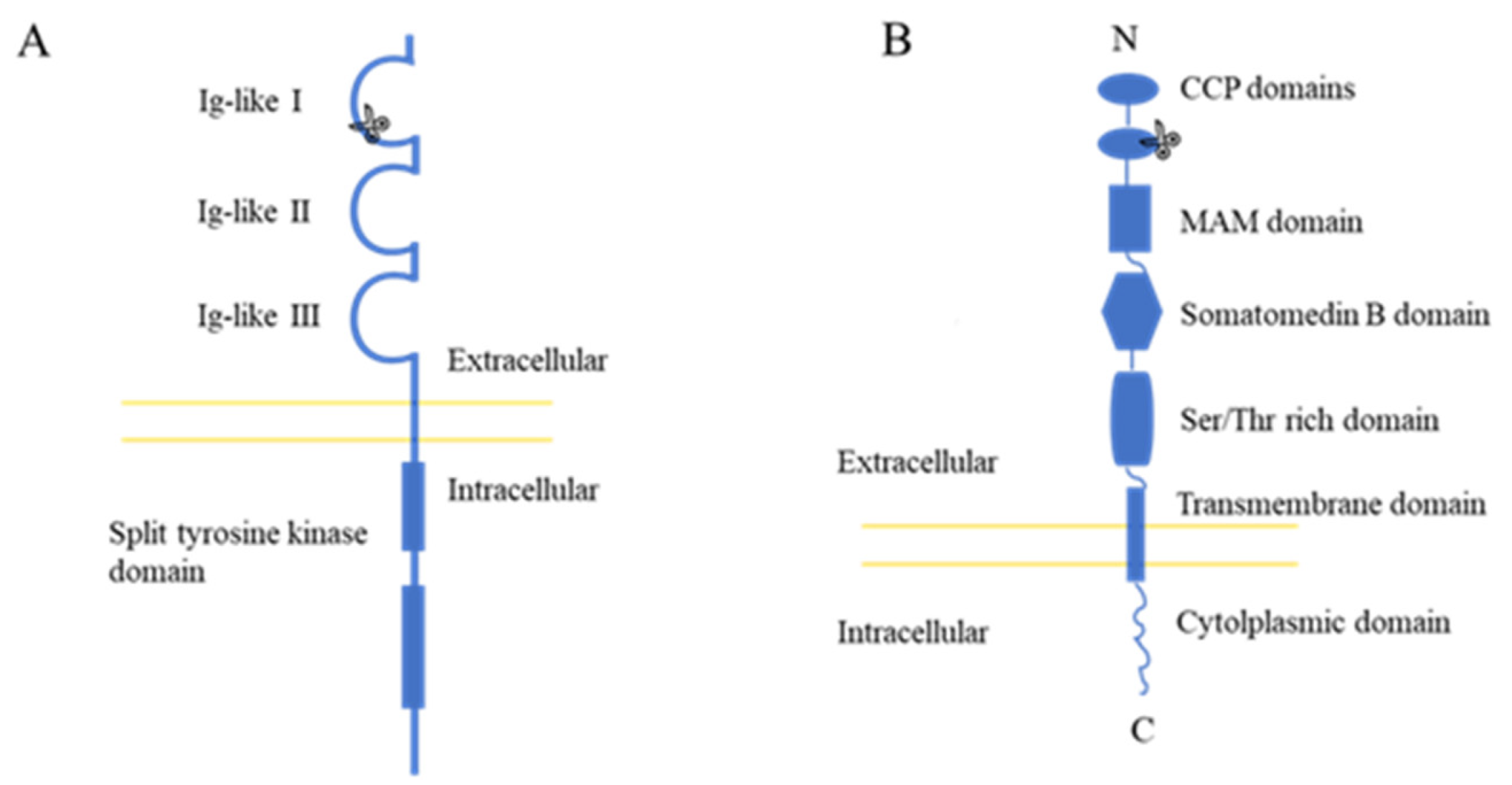{kind=link}
| Strain | N a | LC50 (ng/cm2) b | Slope ± SE c | Toxicity Ratio d |
|---|---|---|---|---|
| DH19 | 168 | 36.64 (22.52–59.61) | 4.46 ± 0.65 | 1 |
| FGFR-KO | 168 | 39.20 (22.61–67.99) | 4.44 ± 0.75 | 1.01 |
| SRC-KO | 168 | 51.85 (24.80–108.38) | 4.80 ± 1.29 | 1.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, Y.; Jin, M.; Chakrabarty, S.; Yang, B.; Li, Q.; Cheng, Y.; Zhang, L.; Xiao, Y. Sf-FGFR and Sf-SR-C Are Not the Receptors for Vip3Aa to Exert Insecticidal Toxicity in Spodoptera frugiperda. Insects 2022, 13, 547. https://doi.org/10.3390/insects13060547
Shan Y, Jin M, Chakrabarty S, Yang B, Li Q, Cheng Y, Zhang L, Xiao Y. Sf-FGFR and Sf-SR-C Are Not the Receptors for Vip3Aa to Exert Insecticidal Toxicity in Spodoptera frugiperda. Insects. 2022; 13(6):547. https://doi.org/10.3390/insects13060547
Chicago/Turabian StyleShan, Yinxue, Minghui Jin, Swapan Chakrabarty, Bo Yang, Qi Li, Ying Cheng, Lei Zhang, and Yutao Xiao. 2022. "Sf-FGFR and Sf-SR-C Are Not the Receptors for Vip3Aa to Exert Insecticidal Toxicity in Spodoptera frugiperda" Insects 13, no. 6: 547. https://doi.org/10.3390/insects13060547
APA StyleShan, Y., Jin, M., Chakrabarty, S., Yang, B., Li, Q., Cheng, Y., Zhang, L., & Xiao, Y. (2022). Sf-FGFR and Sf-SR-C Are Not the Receptors for Vip3Aa to Exert Insecticidal Toxicity in Spodoptera frugiperda. Insects, 13(6), 547. https://doi.org/10.3390/insects13060547