A Follow-Up to the Geographical Distribution of Anopheles Species in Malaria-Endemic and Non-Endemic Areas of Honduras

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
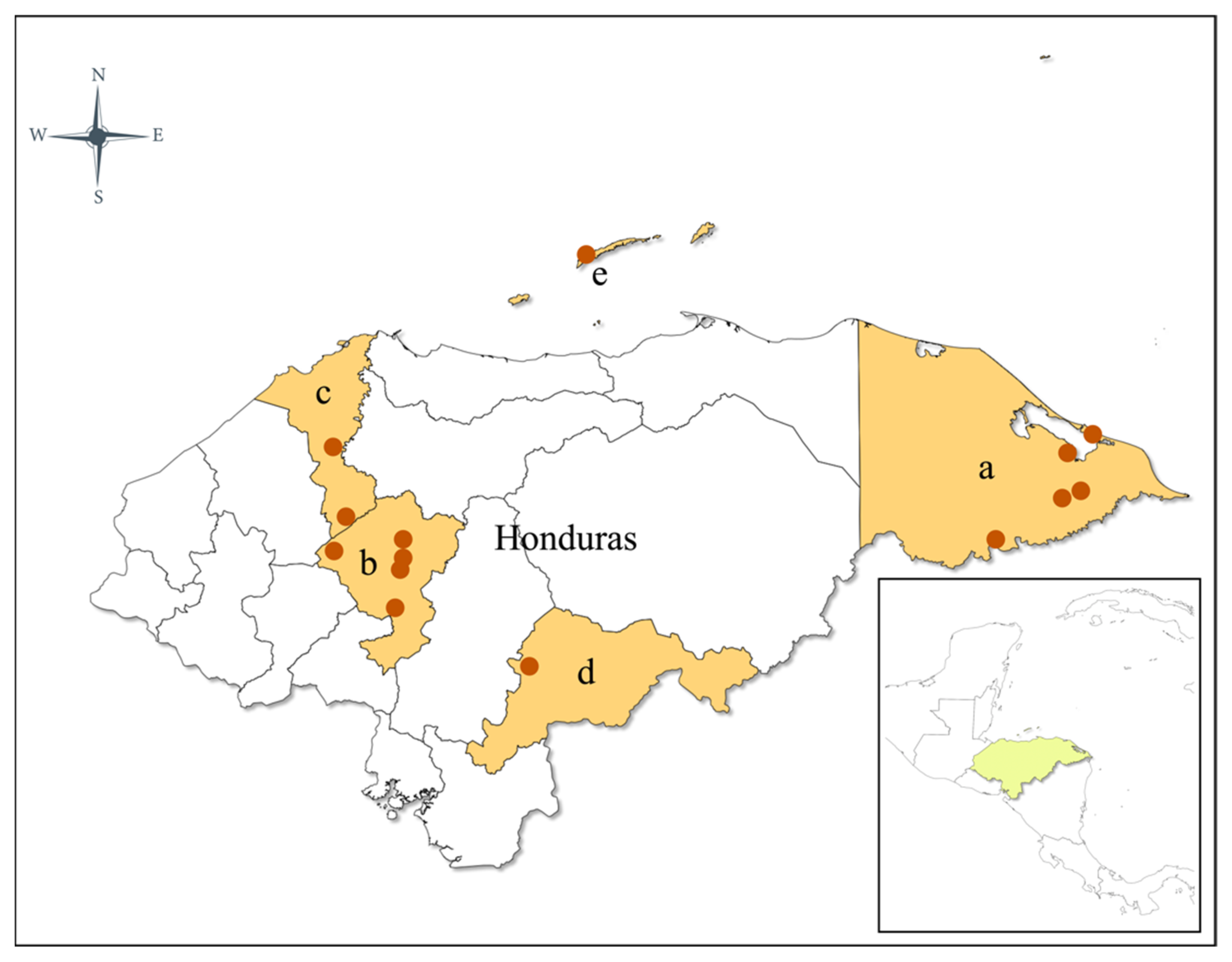
2.1. Study Sites, Mosquito Collection, and Morphological Identification
2.2. DNA Extraction, COI Gene Amplification, and Sequencing
2.3. Sequence Analyses
2.4. cox1 Gene PCR for Plasmodium Sporozoite Detection
3. Results
3.1. Distribution of Anopheles Species
3.2. Nucleotide Sequences and Diversity
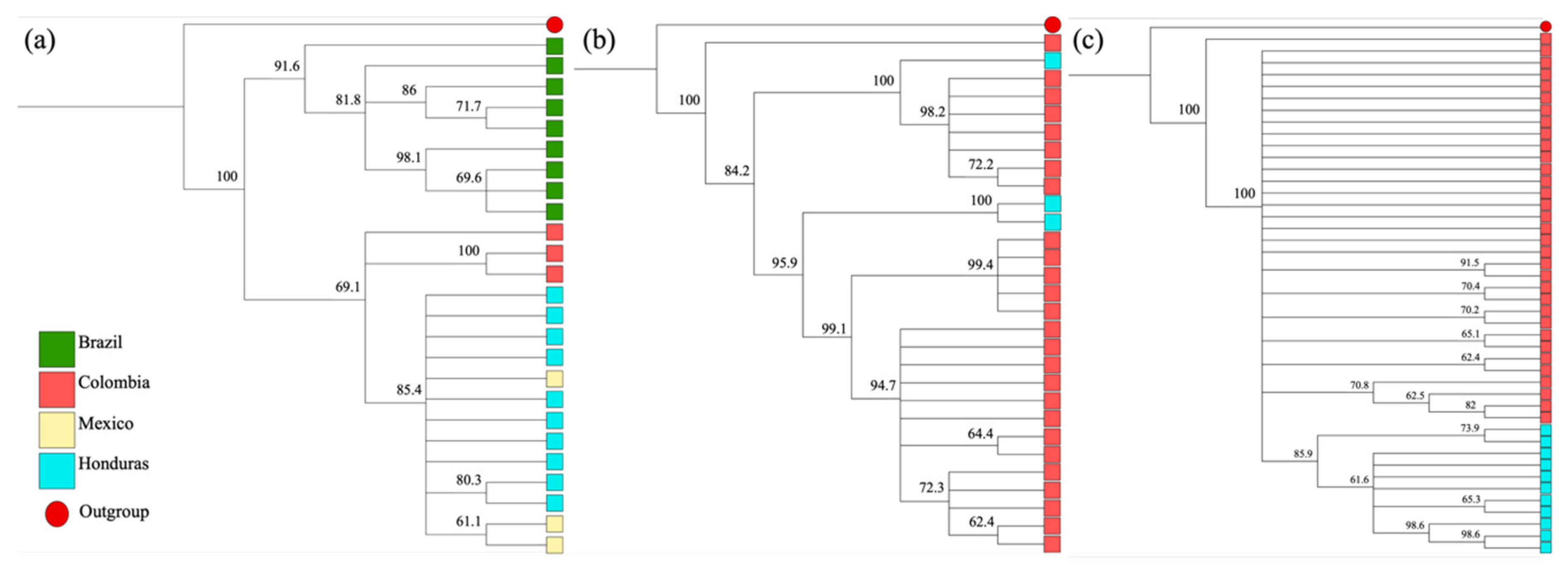
3.3. Phylogenetic Analysis
3.4. Plasmodium spp. DNA Detection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
Copyright Statement
References
- World Health Organization. World Malaria Report 2021; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- World Health Organization. Global Technical Strategy for Malaria 2016–2030, 2021 Update; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Escobar, D.; Ascencio, K.; Ortiz, A.; Palma, A.; Fontecha, G. Distribution and phylogenetic diversity of Anopheles species in malaria endemic areas of Honduras in an elimination setting. Parasit. Vectors 2020, 13, 333. [Google Scholar] [CrossRef]
- Ghassemi-Khademi, T.; Oshaghi, M.A.; Vatandoost, H.; Madjdzadeh, S.M.; Gorouhi, M.A. Utility of Complete Mitochondrial Genomes in Phylogenetic Classification of the Species of Anopheles (Culicidae: Anophelinae). J. Arthropod. Borne Dis. 2021, 15, 1–20. [Google Scholar] [CrossRef]
- Sumruayphol, S.; Chaiphongpachara, T.; Samung, Y.; Ruangsittichai, J.; Cui, L.; Zhong, D.; Sattabongkot, J.; Sriwichai, P. Seasonal dynamics and molecular differentiation of three natural Anopheles species (Diptera: Culicidae) of the Maculatus group (Neocellia series) in malaria hotspot villages of Thailand. Parasit. Vectors 2020, 13, 574. [Google Scholar] [CrossRef]
- Massey, N.C.; Garrod, G.; Wiebe, A.; Henry, A.J.; Huang, Z.; Moyes, C.L.; Sinka, M.E. A global bionomic database for the dominant vectors of human malaria. Sci. Data 2016, 3, 160014. [Google Scholar] [CrossRef]
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Rubio-Palis, Y.; Chareonviriyaphap, T.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; et al. A global map of dominant malaria vectors. Parasit. Vectors 2012, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Sinka, M.E.; Rubio-Palis, Y.; Manguin, S.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Van Boeckel, T.; Kabaria, C.W.; Harbach, R.E.; Hay, S.I. The dominant Anopheles vectors of human malaria in the Americas: Occurrence data, distribution maps and bionomic precis. Parasit. Vectors 2010, 3, 72. [Google Scholar] [CrossRef]
- Ambrose, L.; Ortiz-Barrientos, D.; Cooper, R.D.; Lobo, N.F.; Burkot, T.R.; Russell, T.L.; Beebe, N.W. Gene flow between island populations of the malaria mosquito, Anopheles hinesorum, may have contributed to the spread of divergent host preference phenotypes. Evol. Appl. 2021, 14, 2244–2257. [Google Scholar] [CrossRef]
- Escobar, D.; Ascencio, K.; Ortiz, A.; Palma, A.; Sanchez, A.; Fontecha, G. Blood Meal Sources of Anopheles spp. in Malaria Endemic Areas of Honduras. Insects 2020, 11, 450. [Google Scholar] [CrossRef]
- Stoops, C.A.; Rusmiarto, S.; Susapto, D.; Munif, A.; Andris, H.; Barbara, K.A.; Sukowati, S. Bionomics of Anopheles spp. (Diptera: Culicidae) in a malaria endemic region of Sukabumi, West Java, Indonesia. J. Vector. Ecol. 2009, 34, 200–207. [Google Scholar] [CrossRef]
- Avila, M.I.; Vajda, E.A.; Gutierrez, E.J.; Gibson, D.A.; Renteria, M.M.; Presley, N.; O’Reilly, D.; Burton, T.A.; Tatarsky, A.; Lobo, N.F. Anopheles drivers of persisting malaria transmission in Guna Yala, Panama: An operational investigation. Malar. J. 2021, 20, 443. [Google Scholar] [CrossRef]
- Hinne, I.A.; Attah, S.K.; Mensah, B.A.; Forson, A.O.; Afrane, Y.A. Larval habitat diversity and Anopheles mosquito species distribution in different ecological zones in Ghana. Parasit Vectors 2021, 14, 193. [Google Scholar] [CrossRef] [PubMed]
- Fuller, D.O.; Ahumada, M.L.; Quinones, M.L.; Herrera, S.; Beier, J.C. Near-present and future distribution of Anopheles albimanus in Mesoamerica and the Caribbean Basin modeled with climate and topographic data. Int. J. Health Geogr. 2012, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Palis, Y.; Zimmerman, R.H. Ecoregional classification of malaria vectors in the neotropics. J. Med. Entomol. 1997, 34, 499–510. [Google Scholar] [CrossRef]
- Valderrama, L.; Ayala, S.; Reyes, C.; Gonzalez, C.R. Modeling the Potential Distribution of the Malaria Vector Anopheles (Ano.) pseudopunctipennis Theobald (Diptera: Culicidae) in Arid Regions of Northern Chile. Front. Public Health 2021, 9, 611152. [Google Scholar] [CrossRef]
- Hertig, E. Distribution of Anopheles vectors and potential malaria transmission stability in Europe and the Mediterranean area under future climate change. Parasit. Vectors 2019, 12, 18. [Google Scholar] [CrossRef] [Green Version]
- Collins, F.H.; Paskewitz, S.M. A review of the use of ribosomal DNA (rDNA) to differentiate among cryptic Anopheles species. Insect Mol. Biol. 1996, 5, 1–9. [Google Scholar] [CrossRef]
- Smitz, N.; De Wolf, K.; Gheysen, A.; Deblauwe, I.; Vanslembrouck, A.; Meganck, K.; De Witte, J.; Schneider, A.; Verle, I.; Dekoninck, W.; et al. DNA identification of species of the Anopheles maculipennis complex and first record of An. daciae in Belgium. Med. Vet. Entomol. 2021, 35, 442–450. [Google Scholar] [CrossRef]
- Singh, T.; Hussain, S.S.A.; Rawat, M.; Kar, N.P.; Pasi, S.; Dhiman, R.C. Prevalence of sibling-species of Anopheles (Cellia) fluviatilis complex in Himachal Pradesh, India. J. Vector. Borne Dis. 2021, 58, 85–89. [Google Scholar] [CrossRef]
- Murugan, K.; Vadivalagan, C.; Karthika, P.; Panneerselvam, C.; Paulpandi, M.; Subramaniam, J.; Wei, H.; Aziz, A.T.; Alsalhi, M.S.; Devanesan, S.; et al. DNA barcoding and molecular evolution of mosquito vectors of medical and veterinary importance. Parasitol. Res. 2016, 115, 107–121. [Google Scholar] [CrossRef]
- Motoki, M.T.; Linton, Y.M.; Conn, J.E.; Ruiz-Lopez, F.; Wilkerson, R.C. Phylogenetic Network of Mitochondrial COI Gene Sequences Distinguishes 10 Taxa Within the Neotropical Albitarsis Group (Diptera: Culicidae), Confirming the Separate Species Status of Anopheles albitarsis H (Diptera: Culicidae) and Revealing a Novel Lineage, Anopheles albitarsis J. J. Med. Entomol. 2021, 58, 599–607. [Google Scholar] [CrossRef]
- Fang, Y.; Shi, W.Q.; Zhang, Y. Molecular phylogeny of Anopheles hyrcanus group (Diptera: Culicidae) based on mtDNA COI. Infect. Dis. Poverty 2017, 6, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkerson, R.C.; Strickman, D.; Litwak, T.R. Illustrated key to the female anopheline mosquitoes of Central America and Mexico. J. Am. Mosq. Control Assoc. 1990, 6, 7–34. [Google Scholar] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Kumar, N.P.; Rajavel, A.R.; Natarajan, R.; Jambulingam, P. DNA barcodes can distinguish species of Indian mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2007, 44, 1–7. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Echeverry, D.F.; Deason, N.A.; Makuru, V.; Davidson, J.; Xiao, H.; Niedbalski, J.; Yu, X.; Stevenson, J.C.; Bugoro, H.; Aparaimo, A.; et al. Fast and robust single PCR for Plasmodium sporozoite detection in mosquitoes using the cytochrome oxidase I gene. Malar. J. 2017, 16, 230. [Google Scholar] [CrossRef]
- Vidhya, P.T.; Sunish, I.P.; Maile, A.; Zahid, A.K. Anopheles sundaicus Mosquitoes as Vector for Plasmodium knowlesi, Andaman and Nicobar Islands, India. Emerg. Infect. Dis. 2019, 25, 817–820. [Google Scholar] [CrossRef] [Green Version]
- Villarreal-Trevino, C.; Rios-Delgado, J.C.; Penilla-Navarro, R.P.; Rodriguez, A.D.; Lopez, J.H.; Nettel-Cruz, J.A.; Moo-Llanes, D.A.; Fuentes-Maldonado, G. Composition and abundance of anopheline species according to habitat diversity in Mexico. Salud. Publica Mex. 2020, 62, 388–401. [Google Scholar] [CrossRef]
- Bond, J.G.; Casas-Martinez, M.; Quiroz-Martinez, H.; Novelo-Gutierrez, R.; Marina, C.F.; Ulloa, A.; Orozco-Bonilla, A.; Munoz, M.; Williams, T. Diversity of mosquitoes and the aquatic insects associated with their oviposition sites along the Pacific coast of Mexico. Parasit. Vectors 2014, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Torres-Cosme, R.; Rigg, C.; Santamaria, A.M.; Vasquez, V.; Victoria, C.; Ramirez, J.L.; Calzada, J.E.; Caceres Carrera, L. Natural malaria infection in anophelines vectors and their incrimination in local malaria transmission in Darien, Panama. PLoS ONE 2021, 16, e0250059. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, L.A.; Rigg, C.A.; Calzada, J.E.; Dutary, S.; Bernal, D.; Koo, S.I.; Chaves, L.F. Population Dynamics of Anopheles albimanus (Diptera: Culicidae) at Ipeti-Guna, a Village in a Region Targeted for Malaria Elimination in Panama. Insects 2018, 9, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla-Rodriguez, J.C.; Olivera, M.J.; Ahumada-Franco, M.L.; Paredes-Medina, A.E. Malaria risk stratification in Colombia 2010 to 2019. PLoS ONE 2021, 16, e0247811. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Velez, Y.; Altamiranda-Saavedra, M.; Correa, M.M. Potential distribution of main malaria vector species in the endemic Colombian Pacific region. Trop. Med. Int. Health 2020, 25, 861–873. [Google Scholar] [CrossRef]
- Project, T.M.A. The Malaria Atlas Project. Available online: https://malariaatlas.org/bionomics/ (accessed on 23 March 2022).
- Marteis, L.S.; Sallum, M.A.; Natal, D.; Oliveira, T.M.; Gama, R.A.; Dolabella, S.S.; Santos, R.L. First Record of Anopheles oryzalimnetes, Anopheles argyritarsis, and Anopheles sawyeri (Diptera: Culicidae) in the Caatinga Biome, Semiarid Scrubland of Sergipe State, Brazil. J. Med. Entomol. 2015, 52, 858–865. [Google Scholar] [CrossRef]
- Jimenez, P.; Conn, J.E.; Wirtz, R.; Brochero, H. Anopheles (Diptera: Culicidae) vectors of malaria in Puerto Carreno municipality, Vichada, Colombia. Biomedica 2012, 32 (Suppl. S1), 13–21. [Google Scholar] [CrossRef] [Green Version]
- Dantur Juri, M.J.; Claps, G.L.; Santana, M.; Zaidenberg, M.; Almiron, W.R. Abundance patterns of Anopheles pseudopunctipennis and Anopheles argyritarsis in northwestern Argentina. Acta Trop. 2010, 115, 234–241. [Google Scholar] [CrossRef]
- Silva Jdos, S.; Acel, A.M.; Guimaraes, A.E.; Alencar, J. Anopheles (Nyssorhynchus) argyritarsis larvae found in artificial breeding sites in the State of Mato Grosso. Rev. Soc. Bras. Med. Trop. 2008, 41, 313–314. [Google Scholar] [CrossRef] [Green Version]
- Lardeux, F.; Aliaga, C.; Tejerina, R.; Torrez, L. Comparison of transmission parameters between Anopheles argyritarsis and Anopheles pseudopunctipennis in two ecologically different localities of Bolivia. Malar. J. 2013, 12, 282. [Google Scholar] [CrossRef]
- Chan-Chable, R.J.; Martinez-Arce, A.; Mis-Avila, P.C.; Ortega-Morales, A.I. Confirmation of occurrence of Anopheles (Anopheles) veruslanei Vargas in Quintana Roo, Mexico using morphology and DNA barcodes. Acta Trop. 2018, 188, 138–141. [Google Scholar] [CrossRef]
- Dix, M.; Darsie, R.F., Jr.; Molina, P.A.; Rodriguez, L.; Padilla, N.; Juarez, J. Anopheles neomaculipalpus, first record for Guatemala. J. Am. Mosq. Control Assoc. 1991, 7, 330–331. [Google Scholar] [PubMed]
- Roberts, D.R.; Chan, O.; Pecor, J.; Rejmankova, E.; Manguin, S.; Polanco, J.; Legters, L.J. Preliminary observations on the changing roles of malaria vectors in southern Belize. J. Am. Mosq. Control Assoc. 1993, 9, 456–459. [Google Scholar] [PubMed]
- Alvarez, N.; Gomez, G.F.; Naranjo-Diaz, N.; Correa, M.M. Discrimination of Anopheles species of the Arribalzagia Series in Colombia using a multilocus approach. Infect. Genet. Evol. 2018, 64, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Ahumada, M.L.; Orjuela, L.I.; Pareja, P.X.; Conde, M.; Cabarcas, D.M.; Cubillos, E.F.; Lopez, J.A.; Beier, J.C.; Herrera, S.; Quinones, M.L. Spatial distributions of Anopheles species in relation to malaria incidence at 70 localities in the highly endemic Northwest and South Pacific coast regions of Colombia. Malar. J. 2016, 15, 407. [Google Scholar] [CrossRef] [Green Version]
- Gomez, G.F.; Bickersmith, S.A.; Gonzalez, R.; Conn, J.E.; Correa, M.M. Molecular taxonomy provides new insights into anopheles species of the neotropical arribalzagia series. PLoS ONE 2015, 10, e0119488. [Google Scholar] [CrossRef]
- Morales Viteri, D.; Herrera-Varela, M.; Albuja, M.; Quiroga, C.; Diaz, G.; Del Aguila Morante, C.; Ramirez, D.; Vinetz, J.M.; Bickersmith, S.A.; Conn, J.E. New Records of Anopheles benarrochi B (Diptera: Culicidae) in Malaria Hotspots in the Amazon Regions of Ecuador and Peru. J. Med. Entomol. 2021, 58, 1234–1240. [Google Scholar] [CrossRef]
- Schiemann, D.J.; Pinzon, M.L.; Hankeln, T. Anthropophilic Anopheles species composition and malaria in Tierradentro, Cordoba, Colombia. Mem. Inst. Oswaldo Cruz 2014, 109, 384–387. [Google Scholar] [CrossRef] [Green Version]
- Neves, A.; Urbinatti, P.R.; Malafronte Rdos, S.; Fernandes, A.; Paganini Wda, S.; Natal, D. Malaria outside the Amazon region: Natural Plasmodium infection in anophelines collected near an indigenous village in the Vale do Rio Branco, Itanhaem, SP, Brazil. Acta Trop. 2013, 125, 102–106. [Google Scholar] [CrossRef]
- Rondon, S.; Leon, C.; Link, A.; Gonzalez, C. Prevalence of Plasmodium parasites in non-human primates and mosquitoes in areas with different degrees of fragmentation in Colombia. Malar. J. 2019, 18, 276. [Google Scholar] [CrossRef]
- Gonzalez, C.; Molina, A.G.; Leon, C.; Salcedo, N.; Rondon, S.; Paz, A.; Atencia, M.C.; Tovar, C.; Ortiz, M. Entomological characterization of malaria in northern Colombia through vector and parasite species identification, and analyses of spatial distribution and infection rates. Malar. J. 2017, 16, 431. [Google Scholar] [CrossRef] [Green Version]
- Stein, M.; Luduena-Almeida, F.; Willener, J.A.; Almiron, W.R. Classification of immature mosquito species according to characteristics of the larval habitat in the subtropical province of Chaco, Argentina. Mem. Inst. Oswaldo Cruz 2011, 106, 400–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, J.E.; Rubio-Palis, Y.; Paez, E.; Perez, E.; Sanchez, V.; Vaccari, E. Anopheles (Anopheles) neomaculipalpus: A new malaria vector in the Amazon basin? Med. Vet. Entomol. 2005, 19, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Molina-Cruz, A.; de Merida, A.M.; Mills, K.; Rodriguez, F.; Schoua, C.; Yurrita, M.M.; Molina, E.; Palmieri, M.; Black, W.C.t. Gene flow among Anopheles albimanus populations in Central America, South America, and the Caribbean assessed by microsatellites and mitochondrial DNA. Am. J. Trop. Med. Hyg. 2004, 71, 350–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loaiza, J.R.; Scott, M.E.; Bermingham, E.; Rovira, J.; Conn, J.E. Evidence for pleistocene population divergence and expansion of Anopheles albimanus in Southern Central America. Am. J. Trop. Med. Hyg. 2010, 82, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Hendershot, A.L.; Esayas, E.; Sutcliffe, A.C.; Irish, S.R.; Gadisa, E.; Tadesse, F.G.; Lobo, N.F. A comparison of PCR and ELISA methods to detect different stages of Plasmodium vivax in Anopheles arabiensis. Parasit. Vectors 2021, 14, 473. [Google Scholar] [CrossRef]
- Finney, M.; McKenzie, B.A.; Rabaovola, B.; Sutcliffe, A.; Dotson, E.; Zohdy, S. Widespread zoophagy and detection of Plasmodium spp. in Anopheles mosquitoes in southeastern Madagascar. Malar. J. 2021, 20, 25. [Google Scholar] [CrossRef]
- Zhong, D.; Hemming-Schroeder, E.; Wang, X.; Kibret, S.; Zhou, G.; Atieli, H.; Lee, M.C.; Afrane, Y.A.; Githeko, A.K.; Yan, G. Extensive new Anopheles cryptic species involved in human malaria transmission in western Kenya. Sci. Rep. 2020, 10, 16139. [Google Scholar] [CrossRef]
- Ant, T.; Foley, E.; Tytheridge, S.; Johnston, C.; Goncalves, A.; Ceesay, S.; Ndiath, M.O.; Affara, M.; Martinez, J.; Pretorius, E.; et al. A survey of Anopheles species composition and insecticide resistance on the island of Bubaque, Bijagos Archipelago, Guinea-Bissau. Malar. J. 2020, 19, 27. [Google Scholar] [CrossRef]
- Zogo, B.; Soma, D.D.; Tchiekoi, B.N.; Some, A.; Ahoua Alou, L.P.; Koffi, A.A.; Fournet, F.; Dahounto, A.; Coulibaly, B.; Kande, S.; et al. Anopheles bionomics, insecticide resistance mechanisms, and malaria transmission in the Korhogo area, northern Cote d’Ivoire: A pre-intervention study. Parasite 2019, 26, 40. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Department | Municipality | Coordinates (Latitude and Longitude) | Altitude (masl) | Number of Specimens Collected | Date of Collection |
|---|---|---|---|---|---|
| Gracias a Dios | Puerto Lempira | 14.93567985, −83.84507528/14.94412734, −83.82883771/15.25098087, −83.77352977/15.31331402, −83.5747129/14.700630, −84.335100 | 7–35 | 211 | July to September 2021 |
| Bay Islands | Roatán | 16.323647, −86.563377 | 62 | 32 | September 2021 |
| Comayagua | Comayagua | 14.650778, −87.608472/14.627806, −87.605806/14.650778, −87.608472/14.651083, −87.609444/14.650333, −87.607472 | 430–640 | 23 | June 2018 to May 2021 |
| Comayagua | La Libertad | 14.759750, −87.614861 | 392 | March 2019 | |
| Comayagua | San José | 14.735295, −88.029439 | 701 | July 2021 | |
| El Paraíso | Morocelí | 14.103917, −86.918417/14.104639, −86.919111/14.102944, −86.917694 | 605 | 19 | August 2019 |
| Cortés | Pimienta | 15.289617, −88.029439/15.289617, −87.977116 | 47–237 | 3 | August 2021 |
| Cortés | Santa Cruz de Yojoa | 14.855360, −87.929549 | 742 | June 2021 |
| Department | Municipality | An. (Nyssorhyncus) albimanus | An. (Nyssorhyncus) argyritarsis | An. (Anopheles) pseudopunctipennis | An. (Anopheles) apicimacula | An. (Anopheles) neomaculipalpus | An. (Anopheles) punctimacula | An. (Anopheles) crucians | An. (Anopheles) vestitipennis | Total (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| Gracias a Dios | Puerto Lempira | 168 | 4 | - | 3 | 11 | 2 | 13 | 10 | 211 (73.3%) |
| Bay Islands | Roatán | 32 | - | - | - | - | - | - | - | 32 (11.1%) |
| Comayagua | Comayagua | 7 | - | 7 | - | - | - | - | - | 14 (4.9%) |
| Comayagua | La Libertad | - | - | 2 | - | - | - | - | - | 2 (0.69%) |
| Comayagua | San José | - | 7 | - | - | - | - | - | - | 7 (2.4%) |
| El Paraíso | Morocelí | 19 | - | - | - | - | - | - | - | 19 (6.6%) |
| Cortés | Pimienta | 2 | - | - | - | - | - | - | - | 2 (0.69%) |
| Cortés | Santa Cruz de Yojoa | 1 | - | - | - | - | - | - | - | 1 (0.35%) |
| Total (%) | 229 (79.51%) | 11 (3.82%) | 9 (3.13%) | 3 (1.04%) | 11 (3.82%) | 2 (0.69%) | 13 (4.51%) | 10 (3.47%) | 288 (100%) |
| Feature | An. (Nys.) albimanus | An. (Nys.) argyritarsis | An. (An.) pseudopunctipennis | An. (An.) apicimacula | An. (An.) neomaculipalpus | An. (An.) punctimacula | An. (An.) crucians | An. (An.) vestitipennis |
|---|---|---|---|---|---|---|---|---|
| Nucleotide sequence length | 581 | 596 | 598 | 652 | 572 | 530 | 611 | 615 |
| Number of sequences analysed | 89 | 16 | 14 | 7 | 21 | 2 | 20 | 6 |
| Identical sites | 544 | 589 | 583 | 612 | 567 | - | 584 | 607 |
| Identical sites (%) | 93.6% | 98.8% | 97.5% | 94.0% | 99.1% | - | 95.6% | 98.9% |
| Pairwise % identity | 99.1% | 99.7% | 99.3% | 96.0% | 99.7% | - | 98.2% | 99.4% |
| π | 0.01 | 0.01 | 0.01 | 0.04 | 0.00 | - | 0.02 | 0.01 |
| Haplotype | ||||||||
| Number of sequences analysed | 80 | 10 | 8 | 3 | 11 | 2 | 8 | 5 |
| Nº of haplotypes (nucleotide) | 35 | 6 | 5 | 3 | 4 | - | 8 | 3 |
| Haplotypes/N | 0.44 | 0.6 | 0.62 | 1 | 0.4 | - | 1 | 0.6 |
| Haplotype diversity | 0.94 | 0.84 | 0.79 | 1 | 0.8 | - | 1 | 0.7 |
| Aminoacid sequence length | 209 | 198 | 198 | 216 | 190 | 176 | 203 | 204 |
| Nº of haplotypes (amino acid) | 5 | 2 | 1 | 1 | 1 | - | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escobar, D.; Archaga, O.; Reyes, A.; Palma, A.; Larson, R.T.; Vásquez, G.M.; Fontecha, G. A Follow-Up to the Geographical Distribution of Anopheles Species in Malaria-Endemic and Non-Endemic Areas of Honduras. Insects 2022, 13, 548. https://doi.org/10.3390/insects13060548
Escobar D, Archaga O, Reyes A, Palma A, Larson RT, Vásquez GM, Fontecha G. A Follow-Up to the Geographical Distribution of Anopheles Species in Malaria-Endemic and Non-Endemic Areas of Honduras. Insects. 2022; 13(6):548. https://doi.org/10.3390/insects13060548
Chicago/Turabian StyleEscobar, Denis, Osman Archaga, Allan Reyes, Adalid Palma, Ryan T. Larson, Gissella M. Vásquez, and Gustavo Fontecha. 2022. "A Follow-Up to the Geographical Distribution of Anopheles Species in Malaria-Endemic and Non-Endemic Areas of Honduras" Insects 13, no. 6: 548. https://doi.org/10.3390/insects13060548
APA StyleEscobar, D., Archaga, O., Reyes, A., Palma, A., Larson, R. T., Vásquez, G. M., & Fontecha, G. (2022). A Follow-Up to the Geographical Distribution of Anopheles Species in Malaria-Endemic and Non-Endemic Areas of Honduras. Insects, 13(6), 548. https://doi.org/10.3390/insects13060548