
The King’s Lace Bug Recaredus rex Distant, 1909 (Hemiptera: Heteroptera: Tingidae): Systematic Position, First Palaearctic and Afrotropical Records, and Ecological Niche Modelling



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens Examined
2.2. Photograph Preparation
2.3. Ecological Niche Modelling (ENM)
2.3.1. Occurrence Data
2.3.2. Environmental Variables
2.3.3. Model Optimisation and Maxent Settings
2.4. Output Visualisation and ENM Map Preparation
2.5. Tribal Classification of the Subfamily Tinginae
3. Results and Discussion
3.1. Material Studied, First Country Records and Geographic Coordinates
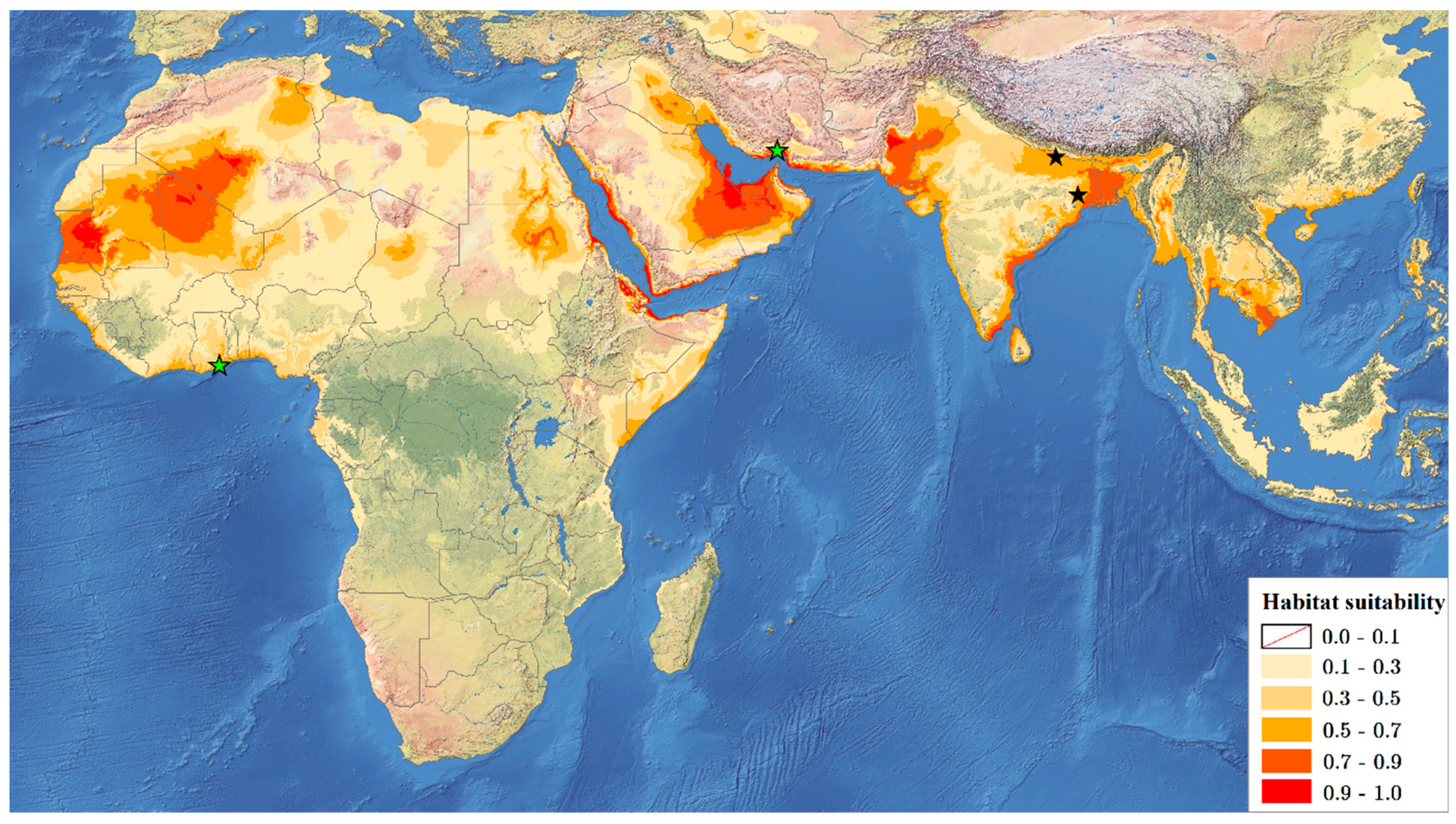
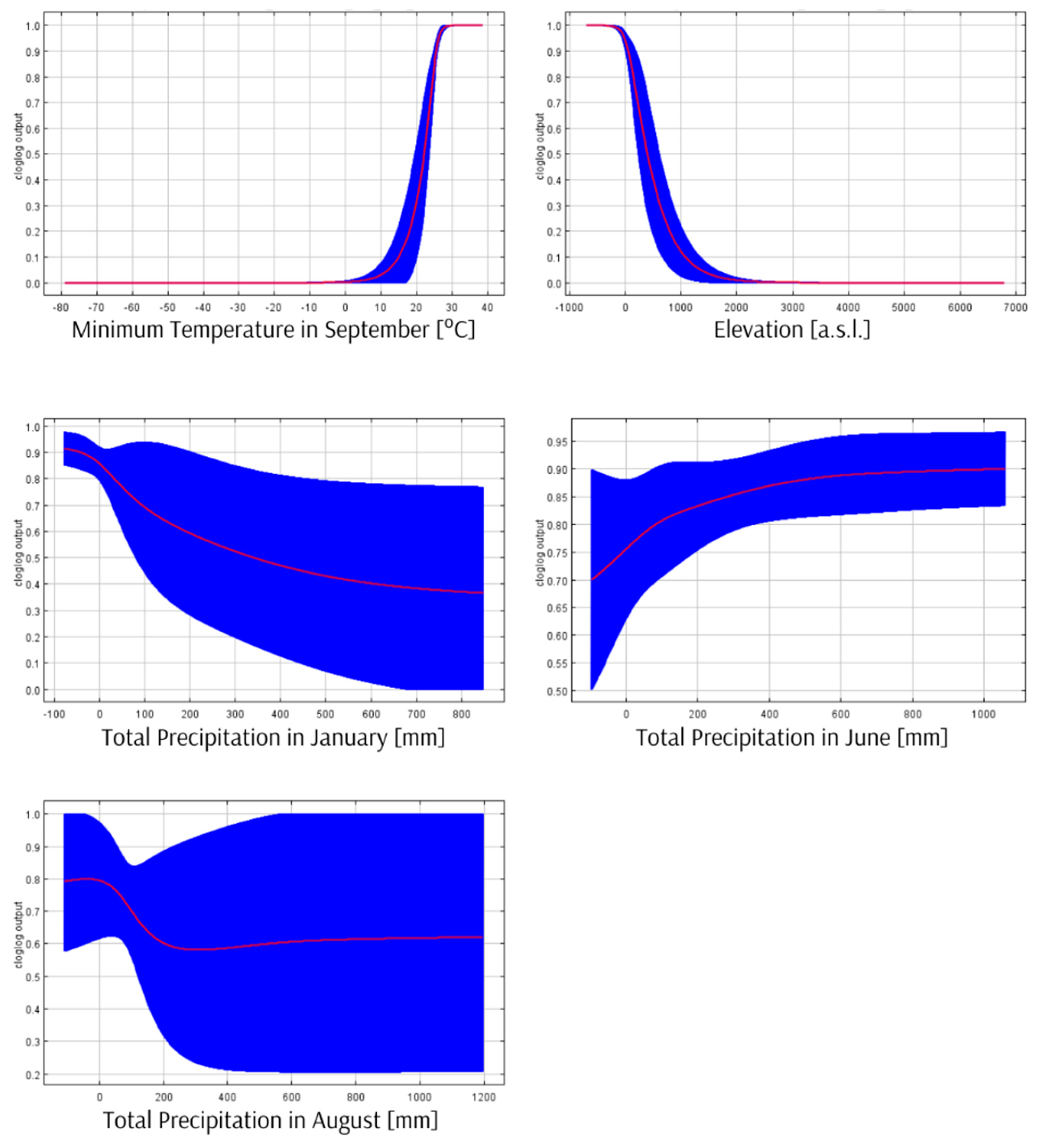
3.2. Ecological Niche Modelling
3.3. Systematic Position and New Tribal Assignment
3.4. Taxonomy
3.5. Key to the World Genera of the Tribe Acalyptaini
3.6. Distribution and Biology
4. Conclusions
- The lace bug genus Recaredus, based on the diagnostic characters provided for the tribe Acalyptaini and the structure of aedeagus, is transferred from the tribe Ypsotingini to the Acalyptaini.
- Recaredus rex, a lace bug species only known from localities distributed within the Oriental region, are recorded for the first time in the Palaearctic region and the Afrotropics.
- Ecological niche modelling results and new records of species suggest its possible Palaeotropical distribution.
- The areas with the best environmental conditions for R. rex indicated by Maxent can be a starting point for further searches for specimens of this species. It might help verify the hypothesis of its Palaeotropical distribution and its oligo- or polyphagy.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Distant, W.L. Rhynchota (Heteroptera) from British India. Ann. de La Sociètè Entomol. de Belg. 1909, 53, 360–376. [Google Scholar]
- Saint Isidore (of Seville). Isidore of Seville, Historia de regibus Gothorum, Vandalorum et Suevorum, chapter 54. In Isidore of Seville’s History of the Goths, Vandals, and Suevi, 2nd Revised ed.; E.J. Brill: Leiden, The Netherlands, 1970; p. 25. [Google Scholar]
- Christys, A. Christians in Al-Andalus, 711-1000; Curzon Press: London, UK, 2002; p. 37. [Google Scholar]
- Wolf, K.B. (Ed.) John of Biclaro, Chronicle, 91. In Conquerors and Chroniclers of Early Medieval Spain, 2nd ed.; University Press: Liverpool, The Netherlands, 1990; p. 74. [Google Scholar]
- Distant, W.L. Rhynchota–Vol. V. Heteroptera: Appendix. In The fauna of British India, including Ceylon and Burma; Taylor and Francis: London, UK, 1910; 362p. [Google Scholar]
- Imaginary Portrait of Reccared I by Dióscoro Puebla. Oil on Canvas (1857). Museo del Prado, Madrid, Spain. Public Domain. Available online: https://commons.wikimedia.org/w/index.php?curid=48073472 (accessed on 7 March 2022).
- Lis, B.; Parveen, S.; Ramamurthy, V.V. Redescription of the Oriental lace-bug Recaredus rex Distant, 1909 (Hemiptera: Tingidae: Tinginae), and its new tribal assignment, with a key to Ypsotingini. Zootaxa 2013, 3702, 597–600. [Google Scholar] [CrossRef] [Green Version]
- Monte, O. Gêneros e genótipos dos tingídeos do mundo. Papéis Avulsos De Zool. 1947, 8, 1–22. [Google Scholar]
- Drake, C.J. Concerning the Cantacaderinae of the World (Hemiptera: Tingidae). Arthropoda 1950, 1, 153–166. [Google Scholar]
- Drake, C.J.; Ruhoff, F.A. Lace-bug genera of the World (Hemiptera: Tingidae). Proc. United States Natl. Mus. 1960, 112, 1–105. [Google Scholar] [CrossRef] [Green Version]
- Drake, C.J.; Ruhoff, F.A. Lacebugs of the World: A Catalog (Hemiptera: Tingidae). USA Natl. Mus. Bull. 1965, 243, 1–634. [Google Scholar] [CrossRef]
- Froeschner, R.C. Lace Bug Genera of the World, II: Subfamily Tinginae: Tribes Litadeini and Ypsotingini (Heteroptera; Tingidae). Smithson. Contrib. Zool. 2001, 611, 1–28. [Google Scholar] [CrossRef]
- Golub, V.B.; Golub, N.V. On the status of the genera complex Acalypta, Dictyonota, Kalama and Derephysia (Heteroptera: Tingidae: Tinginae) having common morphological and karyological features. Zoosystematica Ross. 2019, 28, 228–237. [Google Scholar] [CrossRef]
- Gapon, D.A.; Golub, V.B.; Knudson, A.H. Case 3813—Acalyptini Thomson, 1859 (Hexapoda, Coleoptera) and Acalyptini Blatchley, 1926 (Hexapoda, Heteroptera): Proposed removal of homonymy by emendation of the latter name to Acalyptaini. Bull. Zool. Nomencl. 2019, 76, 175–178. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions (MPB-49) 2011; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Zielińska, A.; Lis, J.A. Can Nysius huttoni F.B. White, 1878 (Hemiptera: Heteroptera: Lygaeidae), a species alien to Europe, have in Poland conditions conducive to existence? Heteroptera Pol.-Acta Faun. 2020, 14, 189–192. [Google Scholar] [CrossRef]
- Zielińska, A.; Lis, B. Evaluation of the possibilities of potential expansion of the oak lace bug Corythucha arcuata (Say, 1832), an invasive species of Tingidae (Hemiptera: Heteroptera), into the territory of Poland. Heteroptera Pol.-Acta Faun. 2020, 14, 175–180. [Google Scholar] [CrossRef]
- Olivera, L.; Minghetti, E.; Montemayor, S.I. Ecological niche modeling (ENM) of Leptoglossus clypealis a new potential global invader: Following in the footsteps of Leptoglossus occidentalis? Bull. Entomol. Res. 2021, 111, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Streito, J.-C.; Chartois, M.; Pierre, É.; Dusoulier, F.; Armand, J.-M.; Gaudin, J.; Rossi, J.-P. Citizen science and niche modeling to track and forecast the expansion of the brown marmorated stinkbug Halyomorpha halys (Stål, 1855). Sci. Rep. 2021, 11, 11421. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.-P.; Ye, Z.; Du, J.; Zhang, D.-L.; Zhen, Y.-h.; Zheng, C.-g.; Zhao, L.; Li, M.; Bu, W.-J. Range wide molecular data and niche modeling revealed the Pleistocene history of a global invader (Halyomorpha halys). Sci. Rep. 2016, 6, 23192. [Google Scholar] [CrossRef] [Green Version]
- Solhjouy-Fard, S.; Sarafrazi, A.; Moeini, M.M.; Ahadiyat, A. Predicting habitat distribution of five heteropteran pest species in Iran. J. Insect Sci. 2013, 13, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montemayor, S.I.; Dellapé, P.M.; Melo, M.C. Predicting the potential invasion suitability of regions to cassava lacebug pests (Heteroptera: Tingidae: Vatiga spp.). Bull. Entomol. Res. 2015, 105, 173–181. [Google Scholar] [CrossRef]
- Zhu, G.-p.; Liu, G.-q.; Bu, W.-J.; Lis, J.A. Geographic distribution and niche divergence of two stinkbugs, Parastrachia japonensis and Parastrachia nagaensis. J. Insect Sci. 2013, 13, 102. [Google Scholar] [CrossRef] [Green Version]
- Chłond, D.; Bugaj-Nawrocka, A. Distribution Pattern and Climate Preferences of the Representatives of the Cosmopolitan Genus Sirthenea Spinola, 1840 (Heteroptera: Reduviidae: Peiratinae). PLoS ONE 2015, 10, e0140801. [Google Scholar] [CrossRef] [Green Version]
- Parra-Henao, G.; Suárez-Escudero, L.C.; González-Caro, S. Potential Distribution of Chagas Disease Vectors (Hemiptera, Reduviidae, Triatominae) in Colombia, Based on Ecological Niche Modeling. J. Trop. Med. 2016, 2016, 1439090. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Chen, D.-y.; Yuan, J.-J.; Zheng, C.-g.; Yang, X.; Wang, W.-w.; Zhang, Y.-y.; Wang, S.; Jiang, K.; Bu, W.-J. Are population isolations and declines a threat to island endemic water striders? A lesson from demographic and niche modelling of Metrocoris esakii (Hemiptera: Gerridae). Mol. Ecol. 2020, 29, 4573–4587. [Google Scholar] [CrossRef] [PubMed]
- Bugaj-Nawrocka, A.; Sawka-Gądek, N.; Chłond, D. Prediction of hybridisation zones of selected species of the genus Platymeris (Hemiptera: Reduviidae) supported by laboratory crossbreeding. Austral Entomol. 2020, 59, 323–336. [Google Scholar] [CrossRef]
- Minghetti, E.; Olivera, L.; Montemayor, S.I. Ecological niche modelling of Gargaphia decoris (Heteroptera), a biological control agent of the invasive tree Solanum mauritianum (Solanales: Solanaceae). Pest Manag. Sci. 2020, 76, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Lis, J.A.; Zielińska, A.; Lis, B. Ecological niche modelling and first records from Namibia and Zimbabwe validate the amphi-equatorial distribution of Byrsinus pseudosyriacus (Hemiptera: Heteroptera: Cydnidae). Afr. J. Ecol. 2022, 60, 100–104. [Google Scholar] [CrossRef]
- Fan, S.; Chen, C.; Zhao, Q.; Wei, J.; Zhang, H. Identifying Potentially Climatic Suitability Areas for Arma custos (Hemiptera: Pentatomidae) in China under Climate Change. Insects 2020, 11, 674. [Google Scholar] [CrossRef]
- Hoberlandt, L. Results of the Czechoslovak-Iranian entomological expeditions to Iran. Introduction to the Third expedition 1977. Acta Entomol. Musei Natl. Pragae 1983, 41, 5–24. [Google Scholar]
- Lis, B. Comparative studies on the ductus seminist of aedeagus in Tingoidea (Hemiptera: Heteroptera). Pol. J. Entomol. 2004, 73, 245–258. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Ortega-Huerta, M.A.; Peterson, A.T. Modeling ecological niches and predicting geographic distributions: A test of six presence-only methods. Rev. Mex. Biodivers. 2008, 79, 205–216. [Google Scholar]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions (Version 3.4.1); American Museum of Natural History: New York, NJ, USA, 2022; Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 3 March 2022).
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Townsend Peterson, A.; Graham, C.H.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Bean, W.T.; Stafford, R.; Brashares, J.S. The effects of small sample size and sample bias on threshold selection and accuracy assessment of species distribution models. Ecography 2012, 35, 250–258. [Google Scholar] [CrossRef]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Li, Y.; Ding, C. Effects of sample size, sample accuracy and environmental variables on predictive performance of MaxEnt model. Pol. J. Ecol. 2016, 64, 303–312. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Google Earth 9.158.0.0. Available online: http://Earth.Google.Com/Web/ (accessed on 3 March 2022).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Realease 10.7.1; Environmental Systems Research Institute Inc.: Redlands, CA, USA, 2022. [Google Scholar]
- Jueterbock, A.; Smolina, I.; Coyer, J.A.; Hoarau, G. The fate of the Arctic seaweed Fucus distichus under climate change: An ecological niche modeling approach. Ecol. Evol. 2016, 6, 1712–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Natural Earth. Natural Earth I with Shaded Relief, Water, and Drainages. Available online: https://www.naturalearthdata.com/downloads/10m-raster-data/10m-natural-earth-1/ (accessed on 3 March 2022).
- Open Data Soft. World Administrative Boundaries (by World Food Programme (UN Agency)). Available online: https://public.opendatasoft.com/explore/dataset/world-administrative-boundaries/information/ (accessed on 3 March 2022).
- Péricart, J. Hémiptères Tingidae euro-mediterranéens. In Faune de France 69; Fédération Française des Sociétés de Sciences Naturelles: Paris, France, 1983; 620p. [Google Scholar]
- Péricart, J.; Golub, V.B. Superfamily Tingoidea Laporte, 1832. In Catalogue of the Heteroptera of the Palaearctic Region. Cimicomorpha I; Netherlands Entomological Society: Wageningen, The Netherlands, 1996; Volume 2, pp. 3–78. [Google Scholar]
- Guilbert, E. Do larvae evolve the same way as adults in Tingidae (Insecta: Heteroptera)? Cladistics 2004, 20, 139–150. [Google Scholar] [CrossRef]
- Guilbert, E.; Damgaard, J.; D’Haese, C.A. Phylogeny of the lacebugs (Insecta: Heteroptera: Tingidae) using morphological and molecular data. Syst. Entomol. 2014, 39, 431–441. [Google Scholar] [CrossRef]
- Schuh, R.T.; Weirauch, C.H. True bugs of the World (Hemiptera: Heteroptera). In Classification and Natural History, 2nd ed.; Monograph Series; Siri Scientific Press: Manchester, UK, 2020; Volume 8, 767p, 32 plates. [Google Scholar]
- Wert Palaniswami, M.S.; Pillai, K.S. Biology of Cochlochila bullita a pest on Chinese potato. J. Root Crops 1983, 9, 59–62. [Google Scholar]
- Stonedahl, G.; Dolling, W.; DuHeaume, G. Identification guide to common tingid pests of the World (Heteroptera: Tingidae). Int. J. Pest Manag. 1992, 38, 438–449. [Google Scholar] [CrossRef]
- Göllner-Scheiding, U. Die Tingidae (Netzwanzen) der Äthiopis (Insecta, Heteroptera: Tingoidea). Katalog der afrikanischen Arten. Nova Supplemcnta Entomol. Keltern 2004, 17, 1–173. [Google Scholar]
- Deckert, J.; Göllner-Scheiding, U. Lace bugs of Namibia (Heteroptera, Tingoidea, Tingidae). Denisia 2006, 19, 823–856. [Google Scholar]
- Schaefer, C.W.; Panizzi, A.R. Heteroptera of Economic Importance; CRC: New York, NY, USA, 2010; 828p. [Google Scholar]
- Smith-Pardo, A.H. The lace bug Cochlochila bullita (Stål) (Heteroptera: Tingidae), an important pest of cultivated herbs in Asia, intercepted at U.S. ports of entry. Bol. Del Mus. Entomológico Fr. Luís Gallego 2013, 5, 16–24. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position Number on the Map | Country | Locality Data | Coordinates |
|---|---|---|---|
| 1 | India | West Bengal: Pareshnath, Bankura district | 22°57′21.3″ N, 86°44′51.4″ E |
| 2 | India | Bihar State: Bettiah, West Champaran district | 26°48′05″ N, 84°30′10″ E |
| 3 | Iran | Hormozgan province: Abbassi, Bang-e Tang | 27°27′ N, 56°18′ E |
| 4 | Ghana | Accra Metropolis district: Legon | 5°39′ N, 0°11′ W |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lis, B.; Zielińska, A.; Lis, J.A. The King’s Lace Bug Recaredus rex Distant, 1909 (Hemiptera: Heteroptera: Tingidae): Systematic Position, First Palaearctic and Afrotropical Records, and Ecological Niche Modelling. Insects 2022, 13, 558. https://doi.org/10.3390/insects13060558
Lis B, Zielińska A, Lis JA. The King’s Lace Bug Recaredus rex Distant, 1909 (Hemiptera: Heteroptera: Tingidae): Systematic Position, First Palaearctic and Afrotropical Records, and Ecological Niche Modelling. Insects. 2022; 13(6):558. https://doi.org/10.3390/insects13060558
Chicago/Turabian StyleLis, Barbara, Anna Zielińska, and Jerzy A. Lis. 2022. "The King’s Lace Bug Recaredus rex Distant, 1909 (Hemiptera: Heteroptera: Tingidae): Systematic Position, First Palaearctic and Afrotropical Records, and Ecological Niche Modelling" Insects 13, no. 6: 558. https://doi.org/10.3390/insects13060558
APA StyleLis, B., Zielińska, A., & Lis, J. A. (2022). The King’s Lace Bug Recaredus rex Distant, 1909 (Hemiptera: Heteroptera: Tingidae): Systematic Position, First Palaearctic and Afrotropical Records, and Ecological Niche Modelling. Insects, 13(6), 558. https://doi.org/10.3390/insects13060558