
Effects of Sublethal Concentrations of Tetracycline Hydrochloride on the Biological Characteristics and Wolbachia Titer in Parthenogenesis Trichogramma pretiosum

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing and Maintenance of T. pretiosum
2.2. Determination of LC5, LC15, and LC35 of Tetracycline Hydrochloride
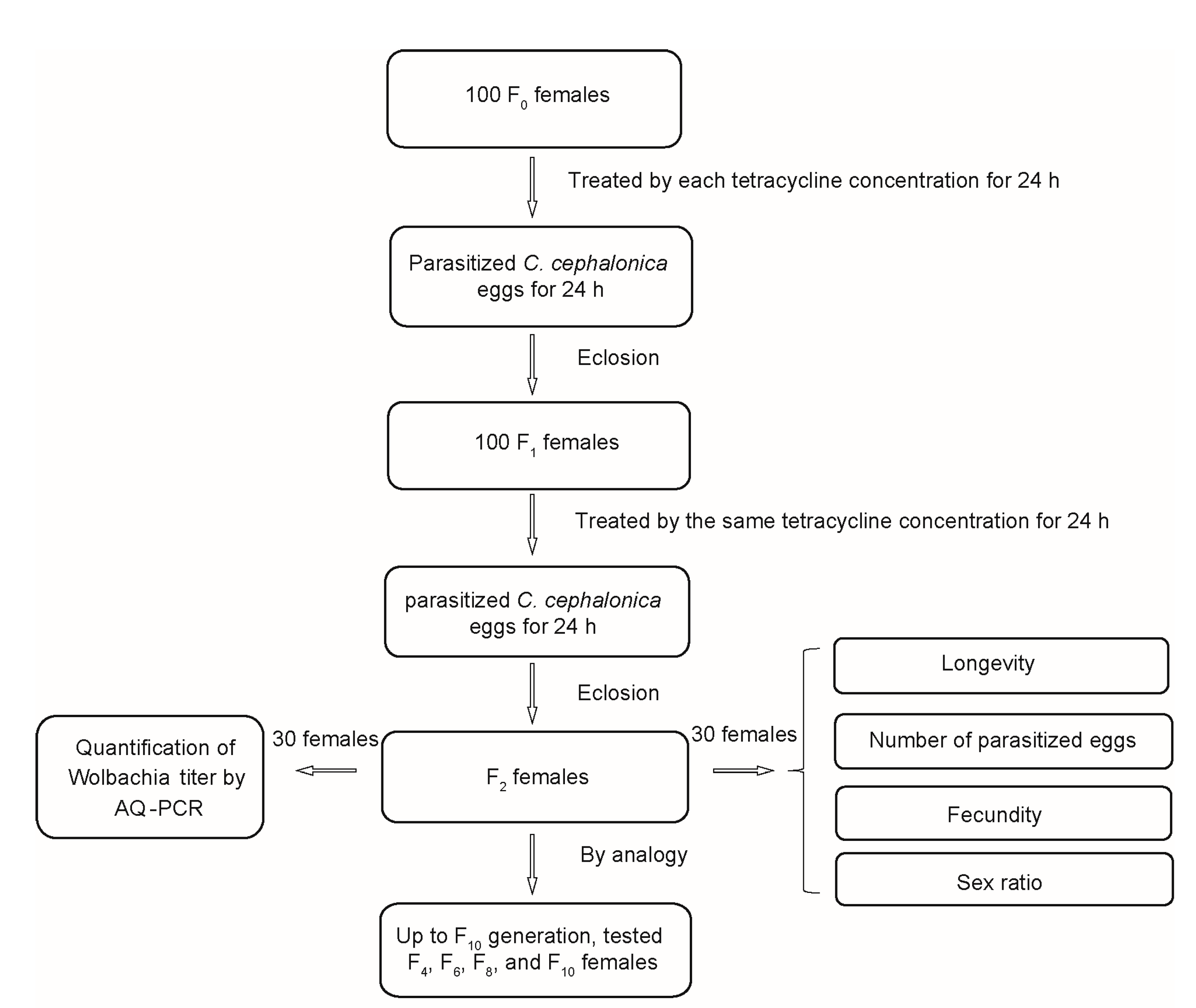
2.3. Tetracycline Treatments
- (1)
- The first assay was carried out to evaluate the sublethal effects of tetracycline on the parasitoid.
- (2)
- The second assay was conducted to detect the Wolbachia titer using AQ-RCR (see quantification of Wolbachia below for details).
- (3)
- The third assay was conducted to keep the F2 generation in order to obtain the F3 generation.
2.4. Quantifications of Wolbachia
2.5. Statistical Analysis
3. Results
3.1. Concentration–Mortality Response
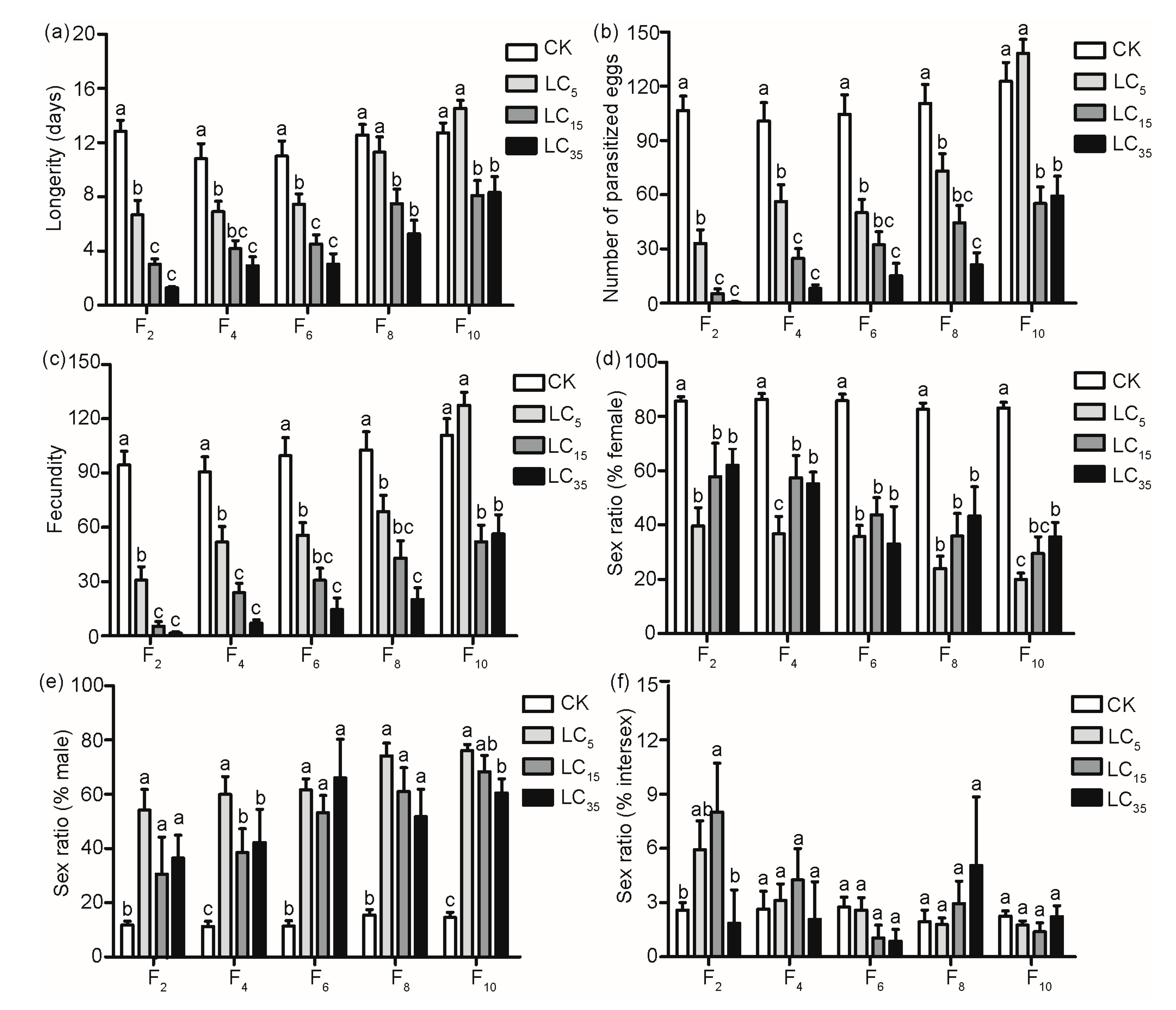
3.2. Sublethal Effects of Tetracycline on Biological Parameters of Treated Female Parasitoids and Their Offspring
3.3. Sublethal Effects of Tetracycline on Life Table Parameters of T. pretiosum
3.4. Sublethal Effects of Tetracycline on Wolbachia Titer in T. pretiosum
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Smith, S.M. Biological control with trichogramma: Advances, successes, and potential of their use. Annu. Rev. Entomol. 1996, 41, 375–406. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.J.; Kelly, S.E.; Hunter, M.S. Population biology of cytoplasmic incompatibility: Maintenance and spread of cardinium symbionts in a parasitic wasp. Genetics 2008, 178, 1003–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.J.; Pannebakker, B.A.; van de Zande, L.; Schwander, T.; Wertheim, B.; Beukeboom, L.W. Diploid males support a two-step mechanism of endosymbiont-induced thelytoky in a parasitoid wasp. BMC Evol. Biol. 2015, 15, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Russell, J.E.; Stouthamer, R. The genetics and evolution of obligate reproductive parasitism in Trichogramma pretiosum infected with parthenogenesis-inducing Wolbachia. Heredity 2010, 106, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Tulgetske, G.M.; Stouthamer, R. Characterization of intersex production in Trichogramma kaykai infected with parthenogenesis-inducing Wolbachia. Naturwissenschaften 2012, 99, 143–152. [Google Scholar] [CrossRef]
- Zug, R.; Hammerstein, P. Still a host of hosts for Wolbachia: Analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef] [Green Version]
- Vavre, F.; de Jong, J.H.; Stouthamer, R. Cytogenetic mechanism and genetic consequences of thelytoky in the wasp Trichogramma cacoeciae. Heredity 2004, 93, 592–596. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.J.; Schwander, T. Patterns and mechanisms in instances of endosymbiont-induced parthenogenesis. J. Evol. Biol. 2017, 30, 868–888. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.T.; Wang, D.S.; He, Y.R. First discovery of Trichogramma oleae (Hymenoptera: Trichogrammatidae) in the wild in China and detection of its endosymbiont Wolbachia. Acta Entomol. Sin. 2021, 64, 409–418. [Google Scholar]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?—A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, D.P.; Pamilo, P.; Kathirithamby, J. Horizontal transmission of Wolbachia by strepsipteran endoparasites? A response to Noda et al., 2001. Mol. Ecol. 2004, 13, 507–509. [Google Scholar] [CrossRef] [PubMed]
- Narita, S.; Shimajiri, Y.; Nomura, M. Strong cytoplasmic incompatibility and high vertical transmission rate can explain the high frequencies of Wolbachia infection in Japanese populations of Colias erate poliographus (Lepidoptera: Pieridae). Bull. Entomol. Res. 2008, 99, 385–391. [Google Scholar] [CrossRef]
- Sasaki, T.; Kubo, T.; Ishikawa, H. Interspecific transfer of Wolbachia between two lepidopteran insects expressing cytoplasmic incompatibility: A Wolbachia variant naturally infecting Cadra cautella causes male killing in Ephestia kuehniella. Genetics 2002, 162, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, J.L.; Liu, C.; Wang, C.P.; Walker, T.; Wang, Y.F. Differentially expressed profiles in the larval testes of Wolbachia infected and uninfected Drosophila. BMC Genom. 2011, 12, 595. [Google Scholar] [CrossRef] [Green Version]
- Hurst, G.D.D.; Jiggins, F.M.; Hinrich Graf von der Schulenbury, J.; Bertrand, D.; West, S.A.; Goriacheva, I.I.; Zakharov, I.A.; Werren, J.H.; Stouthamer, R.; Majerus, M.E.N. Male-killing Wolbachia in two species of insect. Proc. R. Soc. Lond. B. 1999, 266, 735–740. [Google Scholar] [CrossRef] [Green Version]
- Narita, S.; Kageyama, D.; Nomura, M.; Fukatsu, T. Unexpected mechanism of symbiont-induced reversal of insect sex: Feminizing Wolbachia continuously acts on the butterfly Eurema hecabe during larval development. Appl. Environ. Microbiol. 2007, 73, 4332–4341. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.Z.; Li, S.J.; Xue, X.; Yin, X.J.; Ren, S.X.; Jiggins, F.M.; Greeff, J.M.; Qiu, B.L. The intracellular bacterium Wolbachia uses parasitoid wasps as phoretic vectors for efficient horizontal transmission. PLoS Pathog. 2015, 10, e1004672. [Google Scholar] [CrossRef] [Green Version]
- Vavre, F.; Girin, C.; Boulétreau, M. Phylogenetic status of a fecundity-enhancing Wolbachia that does not induce thelytoky in Trichogramma. Insect Mol. Biol. 1999, 8, 67–72. [Google Scholar] [CrossRef]
- Silva, I.M.M.S.; Van Meer, M.M.M.; Roskam, M.M.; Hoogenboom, A.; Gort, G.; Stouthamer, R. Biological control potential of Wolbachia-infected versus uninfected wasps: Laboratory and greenhouse evaluation of Trichogramma cordubensis and T. deion strains. Biocontrol Sci. Technol. 2010, 10, 223–238. [Google Scholar] [CrossRef]
- Stouthamer, R. Influence of microbe-associated parthenogenesis on the fecundity of Trichogramma deion and T. pretiosum. Entomol. Exp. Appl. 2011, 67, 183–192. [Google Scholar] [CrossRef]
- Hohmann, C.L.; Luck, R.F.; Stouthamer, R. Effect of Wolbachia on the survival and reproduction of Trichogramma kaykai Pinto & Stouthamer (Hymenoptera: Trichogrammatidae). Neotrop. Entomol. 2001, 30, 607–612. [Google Scholar]
- Kazuki, M.; Yohsuke, T. Comparison of life history characters of arrhenotokous and Wolbachia-associated thelytokous Trichogramma kaykai pinto and stouthamer (Hymenoptera: Trichogrammatidae). Ann. Entomol. Soc. Am. 2004, 4, 765–769. [Google Scholar]
- Farrokhi, S.; Ashouri, A.; Shirazi, J.; Allahyari, H.; Huigens, M.E. A comparative study on the functional response of Wolbachia-infected and uninfected forms of the parasitoid wasp Trichogramma brassicae. J. Insect Sci. 2010, 10, 167. [Google Scholar] [CrossRef] [Green Version]
- Ning, S.F.; Zhou, J.C.; Liu, Q.Q.; Zhao, Q.; Dong, H. Gradual, temperature-induced change of secondary sexual characteristics in Trichogramma pretiosum infected with parthenogenesis-inducing Wolbachia. PeerJ 2019, 7, e7567. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.X.; Qi, L.D.; Jiang, R.; Du, Y.Z.; Li, Y.X. Incomplete removal of Wolbachia with tetracycline has two-edged reproductive effects in the thelytokous wasp Encarsia formosa (Hymenoptera: Aphelinidae). Sci. Rep. 2017, 7, 44014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Liu, X.; Guo, H. Population dynamics of Wolbachia in Laodelphax striatellus (Fallén) under successive stress of antibiotics. Curr. Microbiol. 2019, 76, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.S.; He, Y.R.; Guo, X.L.; Luo, Y.L. Acute toxicities and sublethal effects of some conventional insecticides on Trichogramma chilonis (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 2012, 105, 1157–1163. [Google Scholar] [CrossRef] [Green Version]
- Finney, D.J. Probit Analysis: A Statistical Treatment of the Sigmoid Response Curve, 3rd ed.; Cambridge University Press: Cambridge, UK, 1972. [Google Scholar]
- Li, Y.Y.; Floate, K.D.; Fields, P.G.; Pang, B.P. Review of treatment methods to remove Wolbachia bacteria from arthropods. Symbiosis 2014, 62, 1–15. [Google Scholar] [CrossRef]
- Zhong, Y.; Li, Z.X. Infuences of tetracycline on the reproduction of the B biotype of Bemisia tabaci (Homoptera: Aleyrodidae). Appl. Entomol. Zool. 2013, 48, 241–246. [Google Scholar] [CrossRef]
- Zha, X.F.; Zhang, W.J.; Zhou, C.Y.; Zhang, L.Y.; Xiang, Z.H.; Xia, Q.Y. Detection and characterization of Wolbachia infection in silkworm. Genet. Mol. Biol. 2014, 37, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Hoerauf, A.; Nissen-Pähle, K.; Schmetz, C.; Henkle-Dührsen, K.; Blaxter, M.L.; Büttner, D.W.; Gallin, M.Y.; Al-Qaoud, K.M.; Lucius, R.; Fleischer, B. Tetracycline therapy targets intracellular bacteria in the filarial nematode Litomosoides sigmodontis and results in filarial infertility. J. Clin. Investig. 1999, 103, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Pike, N.; Kingcombe, R. Antibiotic treatment leads to the elimination of Wolbachia endosymbionts and sterility in the diplodiploid collembolan Folsomia candida. BMC Biol. 2009, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Grenier, S.; Gomes, S.M.; Pintureau, B.; Lassablière, F.; Bolland, P. Use of tetracycline in larval diet to study the effect of Wolbachia on host fecundity and clarify taxonomic status of Trichogramma species in cured bisexual lines. J. Invertebr. Pathol. 2002, 80, 13–21. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [Green Version]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How antibiotics kill bacteria: From targets to networks. Nat. Rev. Microbiol. 2010, 8, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Campbell, E.A.; Korzheva, N.; Mustaev, A.; Murakami, K.; Nair, S.; Goldfarb, A.; Darst, S.A. Structural mechanism for rifampicin inhibition of bacterial RNA polymerase. Cell 2001, 104, 901–912. [Google Scholar] [CrossRef]
- Werren, J.H.; Loehlin, D.W. Curing Wolbachia infections in Nasonia (parasitoid wasp). Cold Spring Harb. Protoc. 2009, 10, pdb.prot5312. [Google Scholar] [CrossRef] [Green Version]
- Werren, J.H.; Loehlin, D.W. The parasitoid wasp Nasonia: An emerging model system with haploid male genetics. Cold Spring Harb. Protoc. 2009, 10, pdb.emo134. [Google Scholar] [CrossRef] [Green Version]
- Kyei-poku, G.K.; Floate, K.D.; Benkel, B.; Goettel, M.S. Elimination of Wolbachia from Urolepis rufipes (Hymenoptera: Pteromalidae) with heat and antibiotic treatments: Implications for host reproduction. Biocontrol Sci. Technol. 2010, 13, 341–354. [Google Scholar] [CrossRef]
- Liu, H.Y.; Wang, Y.K.; Zhi, C.C.; Xiao, J.H.; Huang, D.W. A novel approach to eliminate Wolbachia infections in Nasonia vitripennis revealed different antibiotic resistance between two bacterial strains. FEMS Microbiol. Lett. 2014, 355, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Heimpel, G.E.; de Boer, J.G. Sex determination in the hymenoptera. Annu. Rev. Entomol. 2008, 53, 209–230. [Google Scholar] [CrossRef]
- Stouthamer, R.; Russell, J.E.; Vavre, F.; Leonard, N. Intragenomic conflictin populations infected by parthenogenesis inducing Wolbachia ends with irreversible loss of sexual reproduction. BMC Evol. Biol. 2010, 10, 229. [Google Scholar] [CrossRef]
- Almeida, R.P.; Stouthamer, R. Phylogeny of the Trichogramma endosymbiont Wolbachia, an alpha-proteobacteria (Rickettsiae). Braz. J. Biol. 2018, 78, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Giorgini, M.; Monti, M.M.; Caprio, E.; Stouthamer, R.; Hunter, M.S. Feminization and the collapse of haplodiploidy in an asexual parasitoid wasp harboring the bacterial symbiont Cardinium. Heredity 2009, 102, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Verhulst, E.C.; Beukeboom, L.W.; van de Zande, L. Maternal control of haplodiploid sex determination in the wasp Nasonia. Science 2010, 328, 620–623. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Antibiotic | Slope | LC5 (mg mL−1) (95%CI) | LC15 (mg mL−1) (95%CI) | LC35 (mg mL−1) (95%CI) |
|---|---|---|---|---|
| Tetracycline | 2.56 (0.36) | 0.79 (0.36–1.21) | 1.37 (0.79–1.85) | 2.46 (1.82–3.00) |
| Variable | Source of Variation | df | F | p-Value |
|---|---|---|---|---|
| Longevity | Tetracycline concentration (A) | 3 | 84.2 | <0.001 |
| Generation (B) | 4 | 25.57 | <0.001 | |
| (A) × (B) | 12 | 2.65 | 0.002 | |
| Number of parasitized eggs | Tetracycline concentration (A) | 3 | 117.65 | <0.001 |
| Generation (B) | 4 | 28.27 | <0.001 | |
| (A) × (B) | 12 | 3.24 | <0.001 | |
| Fecundity | Tetracycline concentration (A) | 3 | 107.65 | <0.001 |
| Generation (B) | 4 | 27.97 | <0.001 | |
| (A) × (B) | 12 | 3.06 | <0.001 | |
| Sex ratio (% female) | Tetracycline concentration (A) | 3 | 131.05 | <0.001 |
| Generation (B) | 4 | 12.8 | <0.001 | |
| (A) × (B) | 12 | 2.17 | 0.012 | |
| Sex ratio (% male) | Tetracycline concentration (A) | 3 | 121.41 | <0.001 |
| Generation (B) | 4 | 15.15 | <0.001 | |
| (A) × (B) | 12 | 2.72 | 0.001 | |
| Sex ratio (% intersex) | Tetracycline concentration (A) | 3 | 1.1 | 0.347 |
| Generation (B) | 4 | 2.7 | 0.03 | |
| (A) × (B) | 12 | 1.76 | 0.053 |
| Generation | Tetracycline Concentration | R0 | rm | λ | T (d) | DT |
|---|---|---|---|---|---|---|
| F2 | CK | 76.48 | 0.29 | 1.34 | 14.77 | 2.37 |
| LC5 | 7.55 | 0.18 | 1.20 | 11.51 | 3.83 | |
| LC15 | 2.75 | 0.09 | 1.09 | 11.50 | 7.67 | |
| LC35 | 2.40 | 0.08 | 1.08 | 11.56 | 8.63 | |
| F4 | CK | 70.61 | 0.30 | 1.35 | 14.39 | 2.30 |
| LC5 | 13.73 | 0.23 | 1.26 | 11.50 | 3.00 | |
| LC15 | 10.27 | 0.20 | 1.23 | 11.50 | 3.45 | |
| LC35 | 5.17 | 0.15 | 1.15 | 11.62 | 4.60 | |
| F6 | CK | 82.94 | 0.32 | 1.37 | 13.90 | 2.16 |
| LC5 | 16.04 | 0.24 | 1.27 | 11.77 | 2.88 | |
| LC15 | 12.07 | 0.20 | 1.22 | 12.36 | 3.45 | |
| LC35 | 3.66 | 0.11 | 1.12 | 11.54 | 6.27 | |
| F8 | CK | 84.15 | 0.30 | 1.34 | 14.98 | 2.30 |
| LC5 | 11.69 | 0.21 | 1.23 | 11.90 | 3.29 | |
| LC15 | 8.65 | 0.18 | 1.19 | 12.25 | 3.83 | |
| LC35 | 4.60 | 0.13 | 1.14 | 11.55 | 5.30 | |
| F10 | CK | 87.83 | 0.31 | 1.36 | 14.56 | 2.23 |
| LC5 | 23.11 | 0.26 | 1.30 | 12.05 | 2.23 | |
| LC15 | 10.09 | 0.19 | 1.21 | 11.96 | 3.63 | |
| LC35 | 15.70 | 0.21 | 1.23 | 13.05 | 3.29 |
| Generation | Tetracycline Concentration | Copies μL−1 | Significance |
|---|---|---|---|
| F2 | CK | 10,241.97 ± 373.91 | a |
| LC5 | 259.51 ± 80.29 | b | |
| LC15 | 74.55 ± 7.31 | b | |
| LC35 | 32.75 ± 1.36 | b | |
| F4 | CK | 10,005.82 ± 298.34 | a |
| LC5 | 89.78 ± 20.81 | b | |
| LC15 | 387.12 ± 26.01 | b | |
| LC35 | 111.22 ± 9.20 | b | |
| F6 | CK | 11,003.97 ± 425.56 | a |
| LC5 | 71.12 ± 0.87 | b | |
| LC15 | 242.76 ± 20.71 | b | |
| LC35 | 91.52 ± 2.84 | b | |
| F8 | CK | 10,653.92 ± 303.91 | a |
| LC5 | 28.77 ± 3.56 | c | |
| LC15 | 75.4 ± 2.82 | c | |
| LC35 | 686.84 ± 33.86 | b | |
| F10 | CK | 10,846.11 ± 429.88 | a |
| LC5 | 83.75 ± 7.20 | b | |
| LC15 | 42.44 ± 1.45 | b | |
| LC35 | 469.23 ± 57.71 | b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nian, X.; Tao, X.; Xiao, Z.; Wang, D.; He, Y. Effects of Sublethal Concentrations of Tetracycline Hydrochloride on the Biological Characteristics and Wolbachia Titer in Parthenogenesis Trichogramma pretiosum. Insects 2022, 13, 559. https://doi.org/10.3390/insects13060559
Nian X, Tao X, Xiao Z, Wang D, He Y. Effects of Sublethal Concentrations of Tetracycline Hydrochloride on the Biological Characteristics and Wolbachia Titer in Parthenogenesis Trichogramma pretiosum. Insects. 2022; 13(6):559. https://doi.org/10.3390/insects13060559
Chicago/Turabian StyleNian, Xiaoge, Xiaobing Tao, Zhuangting Xiao, Desen Wang, and Yurong He. 2022. "Effects of Sublethal Concentrations of Tetracycline Hydrochloride on the Biological Characteristics and Wolbachia Titer in Parthenogenesis Trichogramma pretiosum" Insects 13, no. 6: 559. https://doi.org/10.3390/insects13060559
APA StyleNian, X., Tao, X., Xiao, Z., Wang, D., & He, Y. (2022). Effects of Sublethal Concentrations of Tetracycline Hydrochloride on the Biological Characteristics and Wolbachia Titer in Parthenogenesis Trichogramma pretiosum. Insects, 13(6), 559. https://doi.org/10.3390/insects13060559