
Three-Dimensional Reconstruction of Monochamus alternatus Galleries Using CT Scans


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Scanning Wood Segments
2.2. Equipment and Software
2.3. Three-Dimensional Reconstruction of M. Alternatus Galleries
2.4. Determination of Gallery Parameters
2.5. Data Analysis and Statistics
3. Results
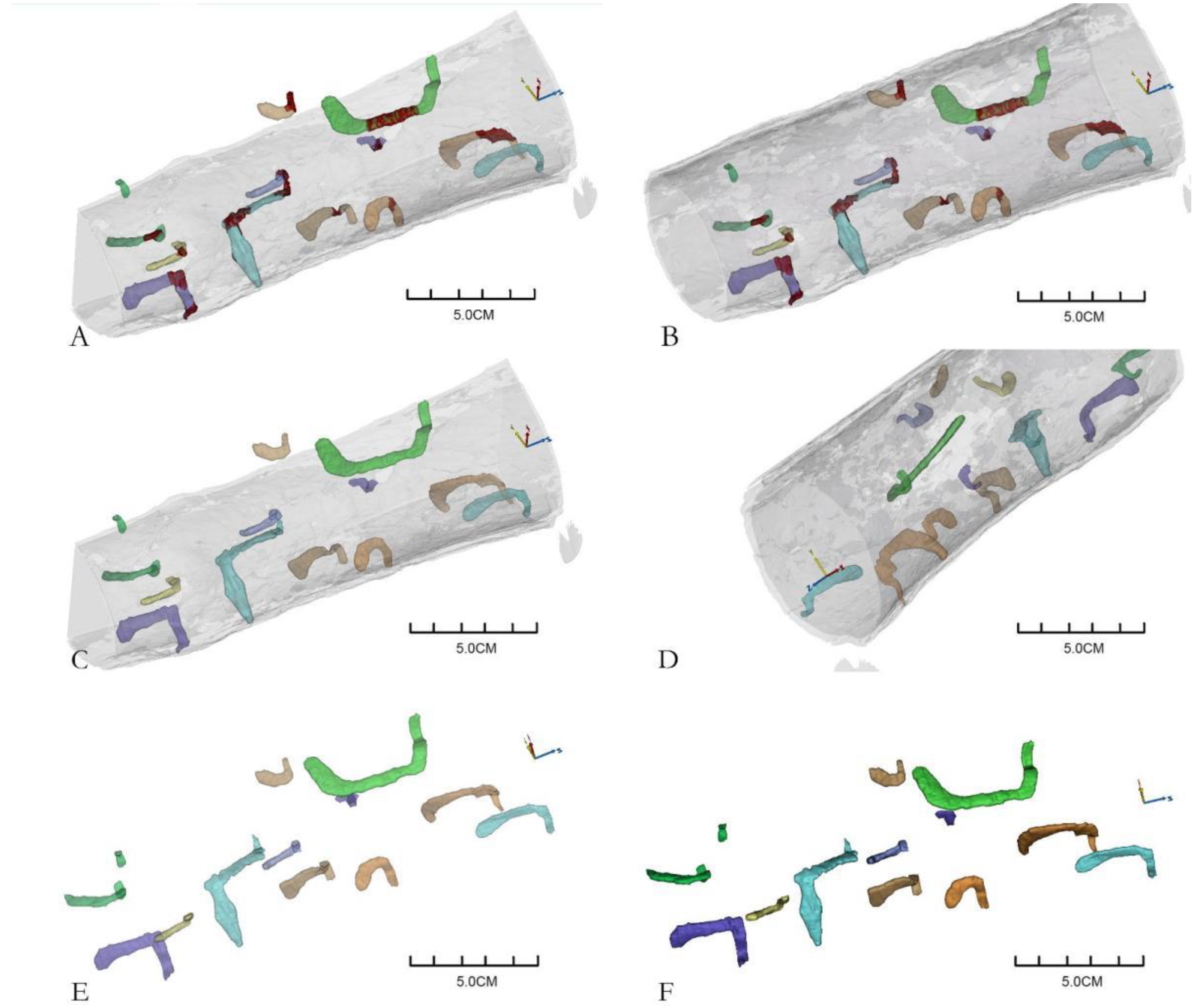
3.1. Scanning Images of M. Alternatus Galleries
3.2. Three-Dimensional Reconstruction of Galleries
3.3. Gallery Types of M. alternatus
3.4. Three-Dimensional Reconstruction of Gallery Blockage
3.5. Structural Parameters of Galleries
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, S. Longicorn Larva; Chongqing Press: Chongqing, China, 1989. [Google Scholar]
- Aukema, E.; Brian, L.; Kent, K.; Corey, C.; Britton, K.O.; Jeffrey, E. Economic impacts of nonnative forest insects in the continental. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef] [PubMed]
- Anbutsu, H.; Togashi, K. Deterred oviposition of Monochamus alternatus (Coleoptera: Cerambycidae) on Pinus densiflora bolts from oviposition scars containing eggs or larvae. Appl. Entomol. Zool. 1996, 31, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q. Evidence for a contact female sex pheromone in Anoplophora chinensis (Forster) (Coloeptera: Cerambycidae: Lamiinae). Coleop Bull. 1998, 52, 363–368. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, Y.; Zhou, J. Biology of RNL’s Boring Trunk and Expelling Farass. J. China Agric. Univ. 1999, 4, 87–91. [Google Scholar] [CrossRef]
- Nowak, D.J.; Pasek, J.E.; Sequeira, R.A.; Crane, D.E.; Mastro, V.C. Potential effect of Anoplophora glabripennis (Coleoptera: Cerambycidae) on urban trees in the United States. Econ. Entomol. 2001, 94, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yang, M.F.; Yang, W.; Yang, C.P.; Dong, C.; Shen, Y.H. A Study on the Spatial Distribution Pattern and the Living-inhabiting Tunnel of the Larvae of Batocera horsfieldi (Hope). J. Sichuan Agric. Univ. 2010, 28, 148–152. [Google Scholar] [CrossRef]
- Tang, Y.; Yang, Z.Q.; Jiang, J.; Wang, X.Y.; Lü, J.; Gao, C. Distribution Pattern of Larvae and Pupae of Massicus raddei in the Trunk of Quercus liaotungensis. Sci. Silvae Sin. 2011, 47, 117–123. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Z.Q.; Zhang, Y.L.; Tang, Y.L. Distribution of Apriona swainsoni larva on the trunk of Sophora japonica and the establishment of its population prediction model. For. Pest. Dis. 2015, 34, 20–22. [Google Scholar] [CrossRef]
- Wei, J.; Yang, Z.; Dai, J.; Du, J. Tritrophic system of tree-trunkborer-insect natural enemy association. Chin. J. Appl. Ecol. 2007, 18, 1125–1131. [Google Scholar] [CrossRef]
- Lv, F.; Hai, X.X.; Wang, Z.G.; Liu, B.X.; Bi, Y.G.; Yan, A.H. Review of the tritrophic interactions of plant-trunk borer-natural enemy. For. Pest. Dis. 2015, 34, 35–39. [Google Scholar] [CrossRef]
- Kundanati, L.; Chahare, N.R.; Jaddivada, S.; Karkisaval, A.G.; Gundiah, N. Cutting mechanics of wood by beetle larval mandibles. J. Mech. Behav. Biomed. Mater. 2020, 112, 104027. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Zheng, Z.G.; Xi, J.F.; Liu, Y.; Lu, J.; Shao, M. Test of pheromone attractant to trap bark beetle in timber. Plant Quar. 2008, 22, 3. [Google Scholar]
- Bilski, P.; Bobinski, P.; Krajewski, A.; Witomski, P. Detection of wood boring insects’ larvae based on the acoustic signal analysis and the artificial intelligence algorithm. Arch. Acoust. J. Pol. Acad. Sci. 2017, 42, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Segaran, N.; Saini, G.; Mayer, J.L.; Naidu, S.; Patel, I.; Alzubaidi, S.; Oklu, R. Application of 3D printing technology in clinical medicine. Med. Pharm. J. Chin. Peoples Lib. Army 2019, 31, 112–116. [Google Scholar]
- Sun, Y.; Tuo, X.; Jiang, Q.; Zhang, H.; Chen, Z.; Zong, S.; Luo, Y. Drilling Vibration Identification Technique of Two Pest Based on Lightweight Neural Networks. Sci. Silvae Sin. 2020, 56, 103–111. [Google Scholar]
- Cruvinel, P.E.; Naime, J.M.; Borges, M.; Macedo, Á.; Zhang, A. Detection of beetle damage in forests by X-ray CT image processing. Rev. Rvore 2003, 27, 1095–1100. [Google Scholar] [CrossRef] [Green Version]
- Aguilera-Olivares, D.; Palma-Onetto, V.; Flores-Prado, L.; Zapata, V.; Niemeyer, H.M. X-ray computed tomography reveals that intraspecific competition promotes soldier differentiation in a one-piece nesting termite. Entomol. Exp. Et Appl. 2017, 163, 26–34. [Google Scholar] [CrossRef]
- Hu, X.; Qu, T.; Zheng, H. Control countermeasures of Bursaphelenchus xylophilus in China. Forest Pest. and Dis. 1997, 3, 30–32. [Google Scholar]
- Yang, B.; Wang, L.F.; Zhao, W.X.; Xu, F.Y.; Zhang, P.; Li, Z.P. The Latent Infection of Bursaphelenchus xylophilus and A New Transmission Way of PWN by Monochamus alternatus. For. Res. 2002, 15, 251–255. [Google Scholar] [CrossRef]
- Mamiya, Y.; Enda, N. Transmission of Bursaphelenchus lignicolus (Nematode: Aphelenchoididae) by Monochamus alternatus (Coleoptera: Cerambycidae). Nematologica 1972, 18, 159–162. [Google Scholar] [CrossRef]
- Zhao, J. The Study of Living and Inhabiting Tunnel of Monochamus alternatus. For. Res. 2005, 18, 62–65. [Google Scholar] [CrossRef]
- Togashi, K.; Kasuga, H.; Yamashita, H.; Iguchi, K. Effect of host tree species on larval body size and pupal-chamber tunnel of Monochamus alternatus (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 2008, 43, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.K.; Ma, S.C.; Yang, H.W.; Yang, Q.H.; Zhou, C.G.; Qiao, L.Q. Study on the gallery system of Monochamus alternatus Hope. For. Res. 2022, 35, 148–153. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, Y.Q.; Zhang, Y.R.; Ren, L.L. Adaptability of external morphological structures and capacity for crop damage of some cerambycid larvae. Chin. J. Appl. Entomol. 2013, 50, 790–799. [Google Scholar]
- Ma AK, W.; Alghamdi, A.A.; Tofailli, K.; Spyrou, N.M. X-ray CT in the detection of palm weevils. J. Radioanal. Nucl. Chem. 2012, 291, 353–357. [Google Scholar] [CrossRef]
- Lyons, C.L.; Tshibalanganda, M.; Plessis, A.D. Using CT-scanning technology to quantify damage of the stem-boring beetle, Aphanasium australe, a biocontrol agent of Hakea sericea in South Africa. Biocontrol. Sci. Technol. 2020, 30, 33–41. [Google Scholar] [CrossRef]
- Hao, D.; Zhang, Y.; Dai, H.; Wang, Y. Oviposition preference of Monochamus alternatus Hope ( Coleoptera:Cerambycidae) to host plants. Acta Entomol. Sin. 2005, 48, 460–464. [Google Scholar] [CrossRef]
- Gregory, N.; AGómez Oliveira, T.; Nichols, E. Big dung beetles dig deeper: Traitbased consequences for faecal parasite transmission. Int. J. Parasitol. 2015, 45, 101–105. [Google Scholar] [CrossRef]
- Gao, S.Q.; Tang, Y.L.; Zhang, Y.L.; Yang, Z.Q.; Fu, F.Y. Distribution of Monochamus alternatus on the Trunks of Pinus Massoniana. For. Res. 2015, 28, 708–712. [Google Scholar]
- Wen, X.; Li, D.; Liao, F.F.; Wang, Z.H.; Xiong, C.Y. The spatial structure and distribution pattern of Monochamus alternatus Hope larvae on the bait-trees in Pinus massoniana forests. J. South. Agric. 2018, 49, 1995–2000. [Google Scholar]
- Johnson, M.P.; Allcock, A.L.; Pye, S.E.; Fitton, S.C.M. The effects of dispersal mode on the spatial distribution patterns of Intertidal molluscs. J. Anim. Ecol. 2001, 70, 641–649. [Google Scholar] [CrossRef]
- Tang, C.; Huang, S.; Chen, Q.S.; Song, H.F.; Kang, W.T.; He, X.Y. Biological characteristics of Monochamus alternatus: Life habit. J. Biosaf. 2005, 14, 209–213. [Google Scholar]
- Jiang, Y.; Yang, Z.Q.; Wang, X.Y.; Hou, Y.X. Molecular identification of sibling species of Sclerodermus (Hymenoptera: Bethylidae) that parasitize buprestid and cerambycid beetles by using partial sequences of mitochondrial DNA cytochrome oxidase subunit 1 and 28S ribosomal RNA gene. PLoS ONE 2015, 10, e0119573. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.F. Monochamus Alternatus, Forest Insects of China; Xiao, G.R., Ed.; Chinese Forestry Press: Beijing, China, 1992; pp. 483–485. [Google Scholar]
- Anbutsu, H.; Togashi, K. Effects of spatio-temporal intervals between newly-hatched larvae on larval survival and development in Monochamus alternatus (Coleoptera: Cerambycidae). Res. Popul Ecol. 1997, 39, 181–189. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, H. A preliminary study on the Apriona swainsoni larval boring and wormhole system. Plant. Prot. 2009, 35, 119–122. [Google Scholar] [CrossRef]
- Chai, Z.Q.; Chen, G.H.; Zhu, Q.; Yang, F.J.; Ding, L.F.; Chen, H.M.; Zhang, X.M.; Li, Z.B.; Wu, J.X.; Ti, Y. Distribution and population dynamics of Xylotrechus quadripes Chevrolat on coffee trunk in Puer. Southwest China J. Agric. Sci. 2020, 33, 2519–2523. [Google Scholar] [CrossRef]
- Niu, Y.; Wei, R.; Yang, Z.Q.; Bao-Fu, D. Study on Frass Extruded by Larvae of Anoplophora glabripennis. For. Res. 2010, 23, 114–119. [Google Scholar]
- Arellano, L. A novel method for measuring dung removal by tunneler dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae) in pastures. Coleopt. Bull. 2016, 70, 185–188. [Google Scholar] [CrossRef]
- Taylor, K.L. The introduction and establishment of insect parasitoids to control Sirex noctilio in Australia. Entomophaga 1977, 21, 429–440. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Z.; Zhang, Y.; Wang, X.; Wu, C.; Ma, S. Biocontrol of the Overwinter Monochamus alternatus with Dastarcus helophoroides. Sci. Silvae Sin. 2014, 50, 92–98. [Google Scholar] [CrossRef]
- Gao, S.; Zhang, Y.; Tang, Y.; Yang, Z.; Wang, X.; Lu, J. Overwintering Characteristics and Cold-Hardiness of Biotype of Dastarcus helophoroides (Coleoptera: Bothrideridae) on Monochamus alternatus (Coleoptera: Cerambycidae). Sci. Silvae Sin. 2016, 52, 68–74. [Google Scholar]
- Hanks, L.M. Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annu. Rev. Entomol. 1999, 44, 483–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Z.; Hou, X.; Li, Z.; Zhou, Y. Application of Computed Tomography (CT) in Nondestructive Testing of Wood. China Wood Ind. 2016, 30, 50–53. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GalleriesType | Entrance Holes Width/mm | Gallery Depth/mm | Vertical Length/mm | Blockage Length/mm | Blockage Volume/mm3 | Total Length of the Gallery/mm | Boring Volume/mm3 |
|---|---|---|---|---|---|---|---|
| C | 2.90 ± 0.44 a | 29.51 ± 8.90 a | 47.68 ± 16.78 a | 25.73 ± 17.10 a | 806.59 ± 777.51a | 79.16 ± 25.77 a | 2988.01 ± 274.9 a |
| S | 3.10 ± 0.26 a | 36.44 ± 5.93 a | 51.10 ± 16.48 a | 38.29 ± 8.69 a | 952.73 ± 296.72 a | 108.73 ± 17.35 a | 3669.26 ± 794.4 a |
| Y | 2.81 ± 0.27 a | 33.11 ± 10.17 a | 42.60 ± 10.10 a | 30.52 ± 21.84 a | 558.21 ± 423.15 a | 88.49 ± 29.04 a | 2530.80 ± 705 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Gao, S.; Wang, J.; Li, W.; Hou, Q.; Qiu, J. Three-Dimensional Reconstruction of Monochamus alternatus Galleries Using CT Scans. Insects 2022, 13, 692. https://doi.org/10.3390/insects13080692
Yang H, Gao S, Wang J, Li W, Hou Q, Qiu J. Three-Dimensional Reconstruction of Monochamus alternatus Galleries Using CT Scans. Insects. 2022; 13(8):692. https://doi.org/10.3390/insects13080692
Chicago/Turabian StyleYang, Huawei, Shangkun Gao, Jinxing Wang, Wen Li, Qingfeng Hou, and Jianfeng Qiu. 2022. "Three-Dimensional Reconstruction of Monochamus alternatus Galleries Using CT Scans" Insects 13, no. 8: 692. https://doi.org/10.3390/insects13080692
APA StyleYang, H., Gao, S., Wang, J., Li, W., Hou, Q., & Qiu, J. (2022). Three-Dimensional Reconstruction of Monochamus alternatus Galleries Using CT Scans. Insects, 13(8), 692. https://doi.org/10.3390/insects13080692