
Exclusion and Repulsion of Popillia japonica (Coleoptera: Scarabaeidae) Using Selected Coverings on High Tunnel Structures for Primocane Red Raspberry



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
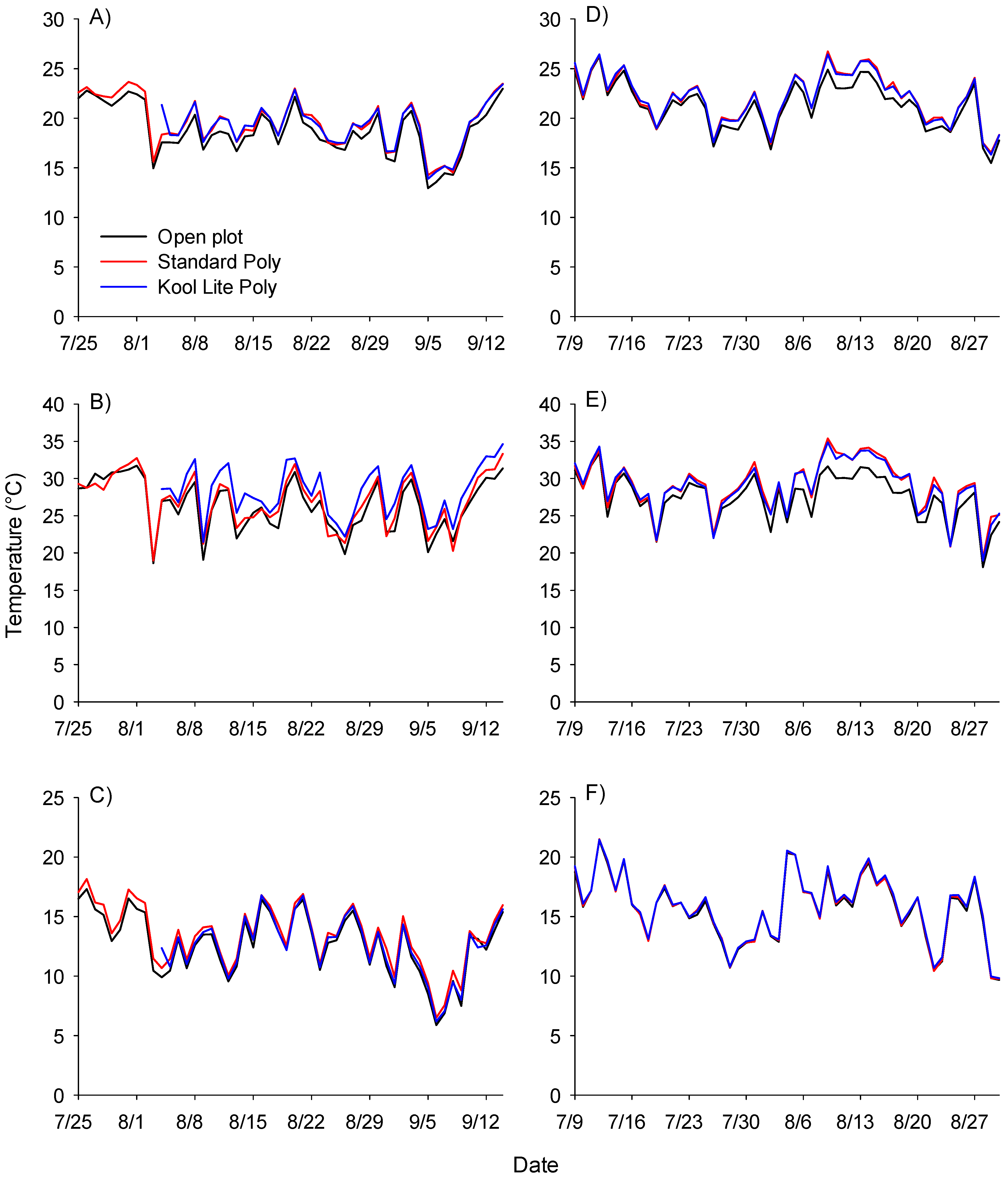
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Digiacomo, G.; Hadrich, J.; Hutchison, W.D.; Peterson, H.; Rogers, M. Economic impact of Spotted Wing Drosophila (Diptera: Drosophilidae) yield loss on Minnesota raspberry farms: A grower survey. J. Integr. Pest Manag. 2019, 10, 11. [Google Scholar] [CrossRef]
- Ebbenga, D.; Wold-Burkness, S.J.; Burkness, E.C.; Hutchison, W.D. Japanese Beetle: An Emerging Pest of Fruit Crops, FruitEdge; University of Minnesota Extension: St. Paul, MN, USA, 2021; Available online: https://fruitedge.umn.edu/japanese-beetle/japanese-beetle-emerging-pest-fruit-crops (accessed on 30 March 2022).
- Ebbenga, D.N.; Burkness, E.C.; Clark, M.; Hutchison, W.D. Impact of adult Popillia japonica (Coleoptera: Scarabaeidae) foliar feeding injury on fruit yield and quality of a temperate, cold-hardy wine grape, ‘Frontenac’. Front. Insect Sci. 2022, 2, 887659. [Google Scholar] [CrossRef]
- Potter, D.A.; Held, D.W. Biology and management of the Japanese beetle. Annu. Rev. Entomol. 2002, 47, 175–205. [Google Scholar] [CrossRef] [PubMed]
- Bushway, L.; Pritts, M.; Handley, D. Raspberry and Blackberry Production Guide for the Northeast, Midwest, and Eastern Canada; NRAES-35; Cornell University: Ithaca, NY, USA, 2008; Available online: https://ecommons.cornell.edu/handle/1813/66930 (accessed on 22 August 2022).
- Fleming, W.E. Biology of the Japanese Beetle; No. 1449; US Department of Agriculture, USDA: Washington, DC, USA, 1972.
- Ebbenga, D.N.; Burkness, E.C.; Hutchison, W.D. Optimizing the use of semiochemical-based traps for efficient monitoring of Popillia japonica (Coleoptera: Scarabaeidae): Validation of a volumetric approach. J. Econ. Entomol. 2022, 115, 869–876. [Google Scholar] [CrossRef] [PubMed]
- Burkness, E.C.; Ebbenga, D.N.; Hutchison, W.D. Evaluation of foliar insecticide control of Japanese beetle in raspberry, 2019. Arthropod Manag. Tests 2020, 45, tsaa009. [Google Scholar] [CrossRef]
- Althoff, E.M.; Rice, K.B. Japanese beetle (Coleoptera: Scarabaeidae) invasion of North America: History, ecology, and management. J. Integr. Pest Manag. 2022, 13, 2. Available online: https://academic.oup.com/jipm/article/13/1/2/6503655 (accessed on 25 May 2022). [CrossRef]
- Nandwani, D.; Nwosisi, S. Global trends in organic agriculture. In Organic Farming for Sustainable Agriculture; Nandwani, D., Ed.; Springer: Berlin, Germany, 2016; pp. 1–35. [Google Scholar]
- Janke, R.R.; Altamimi, M.E.; Khan, M. The use of high tunnels to produce fruit and vegetable crops in North America. Agric. Sci. 2017, 8, 692–715. [Google Scholar] [CrossRef]
- Yao, S.; Rosen, C.J. Primocane fruiting raspberry production in high tunnels in a cold region of the upper midwestern United States. HortTechnology 2011, 21, 429–434. [Google Scholar] [CrossRef]
- Hanson, E.J.; Weihe, M.V.; Schilder, A.C.; Chanon, A.M.; Scheerens, J.C. High tunnel and open field production of floricane- and primocane-fruiting raspberry cultivars. HortTechnology 2011, 21, 412–418. [Google Scholar] [CrossRef]
- Rogers, M.; Burkness, E.C.; Hutchison, W.D. Evaluation of high tunnels for management of Drosophila suzukii in fall-bearing red raspberries: Potential for reducing insecticide use. J. Pest Sci. 2016, 89, 815–821. [Google Scholar] [CrossRef]
- Leach, H.; Van Timmeren, S.; Isaacs, R. Exclusion netting delays and reduces Drosophila suzukii (Diptera: Drosophilidae) infestation in raspberries. J. Econ. Entomol. 2016, 109, 2151–2158. [Google Scholar] [CrossRef] [PubMed]
- Ebbenga, D.; Burkness, E.C.; Hutchison, W.D. Exclusion netting as an alternative management strategy for Spotted-wing Drosophila (Diptera: Drosophilidae) in wine grapes. J. Econ. Entomol. 2019, 112, 2287–2294. [Google Scholar] [CrossRef] [PubMed]
- Candian, V.; Pansa, M.G.; Santoro, K.; Spadaro, D.; Briano, R.; Peano, C.; Tavella, L.; Tedeschi, R. First multi-target application of exclusion net in nectarine orchards: Effectiveness against pests and impact on beneficial arthropods, postharvest rots and fruit quality. Insects 2021, 12, 210. [Google Scholar] [CrossRef] [PubMed]
- Cramer, M.E.; Demchak, K.; Marini, R.; Leskey, T.C. UV-blocking high-tunnel plastics reduce Japanese beetle (Popillia japonica) in red raspberry. HortScience 2019, 54, 903–909. [Google Scholar] [CrossRef]
- SAS Institute. Base SAS 9.4 Procedures Guide: Statistical Procedures; SAS Institute Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Kowles, K.A.; Switzer, P.V. Dynamics of aggregation formation in Japanese beetles, Popillia japonica. J. Insect Behav. 2012, 25, 207–221. [Google Scholar] [CrossRef]
- Carew, J.G.; Gillespie, T.; White, J.; Wainwright, H.; Brennan, R.; Battey, N.H. The control of the annual growth cycle in raspberry. J. Hortic. Sci. Biotechol. 2000, 75, 495–503. [Google Scholar] [CrossRef]
- Galvan, T.L.; Burkness, E.C.; Hutchison, W.D. Enumerative and binomial sequential sampling plans for the multicolored Asian lady beetle (Coleoptera: Coccinellidae) in wine grapes. J. Econ. Entomol. 2007, 100, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Santoiemma, G.; Battisti, A.; Gusella, G.; Cortese, G.; Tosi, L.; Gilioli, G.; Sperandio, G.; Ciampitti, M.; Cavagna, B.; Mori, N. Chemical control of Popillia japonica adults on high-value crops and landscape plants of northern Italy. Crop Prot. 2021, 150, 105808. [Google Scholar] [CrossRef]
- Kistner-Thomas, E.J. The potential global distribution and voltinism of the Japanese beetle (Coleoptera: Scarabaeidae) under current and future climates. J. Insect Sci. 2019, 19, 16. [Google Scholar] [CrossRef] [PubMed]
- Venette, R.C.; Hutchison, W.D. Invasive Insect Species: Grand Challenges, Strategies and Opportunities. Front. Insect Sci. 2021, 1, 650520. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Mean (±SEM) Number Adult Beetles (Total for 25 Leaves and 25 Flower Clusters Per Plot) | |||||
|---|---|---|---|---|---|---|
| Aug. 1 * | Aug. 10 | Aug. 17 | Aug. 24 | Aug. 31 | Sept. 7 | |
| Standard poly + ends; bumblebees added | 5.50 ± 0.87 b | 0.00 ± 0.00 | 1.50 ± 0.50 b | 0.25 ± 0.25 b | 0.00 ± 0.00 b | 0.50 ± 0.29 b |
| Kool Lite poly + ends | 3.50 ± 0.65 b | 0.00 ± 0.00 | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b |
| Open Plot | 43.50 ± 2.60 a | 5.50 ± 3.20 | 14.50 ± 4.25 a | 3.00 ± 1.00 a | 9.25 ± 1.25 a | 5.50 ± 2.02 a |
| NS | ||||||
| Treatment | Mean (±SEM) Number Adult Beetles (Total for 25 Leaves and 25 Flower Clusters Per Plot) | ||||
|---|---|---|---|---|---|
| July 16 | July 30 * | Aug. 9 | Aug. 16 | Aug. 23 | |
| Standard poly + ends; bumblebees added | 3.25 ± 1.31 b | 0.25 ± 0.25 b | 0.50 ± 0.50 b | 0.25 ± 0.25 | 0.25 ± 0.25 b |
| Kool Lite poly + ends | -- | 2.25 ± 1.03 b | 0.00 ± 0.00 b | 0.00 ± 0.00 | 0.00 ± 0.00 b |
| Open Plot | 22.00 ± 2.58 a | 12.75 ± 2.78 a | 30.00 ± 8.51 a | 1.00 ± 0.41 | 4.00 ± 1.47 a |
| NS | |||||
| Treatment | Mean (±SEM) Number Adult Beetles (Total for 25 Leaves and 25 Flower Clusters Per Plot) | |||||||
|---|---|---|---|---|---|---|---|---|
| July 10 | July 17 | July 24 | July 31 | Aug. 7 * | Aug. 14 | Aug. 21 | Aug. 28 | |
| Standard poly + ends | -- | -- | -- | -- | -- | 1.00 ± 0.41 b | 0.50 ± 0.50 b | 0.50 ± 0.50 b |
| Standard poly—no ends | 0.25 ± 0.25 b | 3.50 ± 0.96 | 0.50 ± 0.29 | 0.50 ± 0.50 | 2.75 ± 0.48 b | 3.25 ± 2.02 ab | 2.25 ± 1.03 b | 1.25 ± 0.48 ab |
| Open Plot | 8.50 ± 2.96 a | 13.75 ± 3.57 | 3.50 ± 1.19 | 6.50 ± 2.87 | 12.50 ± 1.32 a | 13.75 ± 5.50 a | 17.25 ± 5.88 a | 6.75 ± 3.25 a |
| NS | NS | NS | ||||||
| Year | Total Number of Adult Beetles | |||||
|---|---|---|---|---|---|---|
| Per 25 Leaves | Per 25 Flower Clusters | Proportion on Leaves | Proportion on Flower Clusters | Χ2 | p Value | |
| 2017 (n = 72) | 270 | 100 | 0.73 | 0.27 | 77.2 | p < 0.0001 |
| 2018 (n = 48) | 189 | 16 | 0.92 | 0.08 | 144.3 | p < 0.0001 |
| 2019 (n = 44) | 218 | 29 | 0.88 | 0.12 | 143.1 | p < 0.0001 |
| Year/Treatment | Mean Daily Temperature (°C) | ||
|---|---|---|---|
| Average | Maximum | Minimum | |
| 2017 | |||
| Standard poly + ends | 19.56 ± 0.09 a | 26.92 ± 0.25 | 13.37 ± 0.10 a |
| Kool Lite poly + ends | 19.14 ± 0.03 b | 28.27 ± 0.23 | 12.53 ± 0.03 b |
| Open Plot | 18.84 ± 0.07 c | 26.46 ± 0.73 | 12.74 ± 0.19 ab |
| NS | |||
| 2018 | |||
| Standard poly + ends | 22.09 ± 0.00 a | 29.00 ± 0.15 | 15.81 ± 0.05 |
| Kool Lite poly + ends | 22.06 ± 0.15 a | 28.87 ± 0.56 | 15.88 ± 0.03 |
| Open Plot | 21.36 ± 0.01 b | 27.52 ± 0.06 | 15.70 ± 0.05 |
| NS | NS | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burkness, E.C.; Ebbenga, D.N.; Toninato, A.G.; Hutchison, W.D. Exclusion and Repulsion of Popillia japonica (Coleoptera: Scarabaeidae) Using Selected Coverings on High Tunnel Structures for Primocane Red Raspberry. Insects 2022, 13, 771. https://doi.org/10.3390/insects13090771
Burkness EC, Ebbenga DN, Toninato AG, Hutchison WD. Exclusion and Repulsion of Popillia japonica (Coleoptera: Scarabaeidae) Using Selected Coverings on High Tunnel Structures for Primocane Red Raspberry. Insects. 2022; 13(9):771. https://doi.org/10.3390/insects13090771
Chicago/Turabian StyleBurkness, Eric C., Dominique N. Ebbenga, Adam G. Toninato, and William D. Hutchison. 2022. "Exclusion and Repulsion of Popillia japonica (Coleoptera: Scarabaeidae) Using Selected Coverings on High Tunnel Structures for Primocane Red Raspberry" Insects 13, no. 9: 771. https://doi.org/10.3390/insects13090771
APA StyleBurkness, E. C., Ebbenga, D. N., Toninato, A. G., & Hutchison, W. D. (2022). Exclusion and Repulsion of Popillia japonica (Coleoptera: Scarabaeidae) Using Selected Coverings on High Tunnel Structures for Primocane Red Raspberry. Insects, 13(9), 771. https://doi.org/10.3390/insects13090771