
Moth Diversity Increases along a Continent-Wide Gradient of Environmental Productivity in South African Savannahs

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Sampling
2.2. Data Analyses
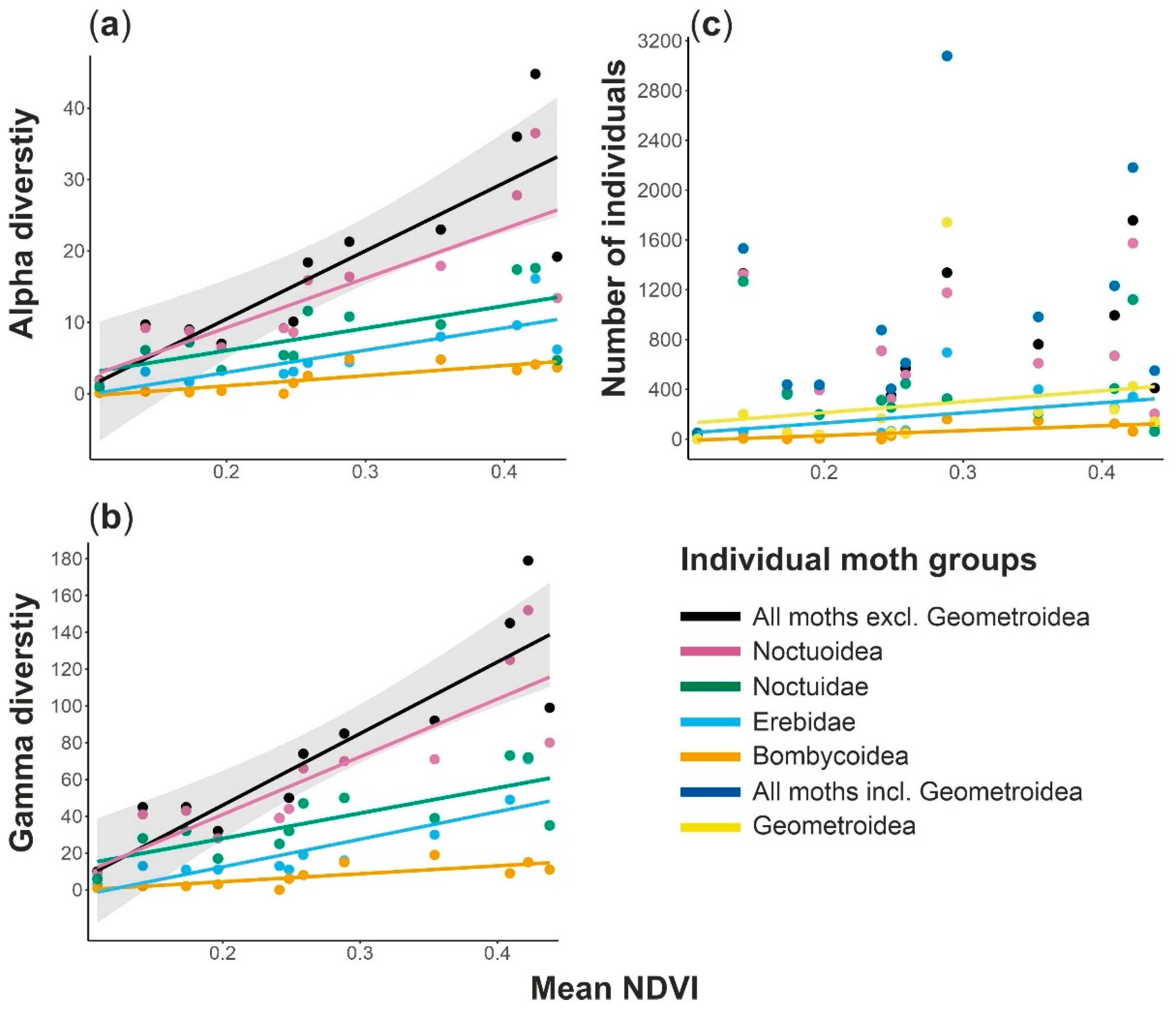
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waide, R.B.; Willig, M.R.; Steiner, C.F.; Mittelbach, G.; Gough, L.; Dodson, S.I.; Juday, G.P.; Parmenter, R. The Relationship between Productivity and Species Richness. Annu. Rev. Ecol. Syst. 1999, 30, 257–300. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Steiner, C.F.; Scheiner, S.M.; Gross, K.L.; Reynolds, H.L.; Waide, R.B.; Willig, M.R.; Dodson, S.I.; Gough, L. What Is the Observed Relationship between Species Richness and Productivity? Ecology 2001, 82, 2381–2396. [Google Scholar] [CrossRef]
- Storch, D. Biodiversity and Its Energetic and Thermal Controls. In Metabolic Ecology; Sibly, R.M., Brown, J.H., Kodric-Brown, A., Eds.; Wiley: Hoboken, NJ, USA, 2012; pp. 120–131. [Google Scholar]
- Pontarp, M.; Bunnefeld, L.; Cabral, J.S.; Etienne, R.S.; Fritz, S.A.; Gillespie, R.; Graham, C.H.; Hagen, O.; Hartig, F.; Huang, S.; et al. The Latitudinal Diversity Gradient: Novel Understanding through Mechanistic Eco-Evolutionary Models. Trends Ecol. Evol. 2019, 34, 211–223. [Google Scholar] [CrossRef]
- Bohdalková, E.; Toszogyova, A.; Šímová, I.; Storch, D. Universality in Biodiversity Patterns: Variation in Species-Temperature and Species-Productivity Relationships Reveals a Prominent Role of Productivity in Diversity Gradients. Ecography 2021, 44, 1366–1378. [Google Scholar] [CrossRef]
- Grime, J.P. Competitive Exclusion in Herbaceous Vegetation. Nature 1973, 242, 344–347. [Google Scholar] [CrossRef]
- Wright, D.H. Species-Energy Theory: An Extension of Species-Area Theory. Oikos 1983, 41, 496. [Google Scholar] [CrossRef]
- Hurlbert, A.H.; Stegen, J.C. When Should Species Richness Be Energy Limited, and How Would We Know? Ecol. Lett. 2014, 17, 401–413. [Google Scholar] [CrossRef]
- Srivastava, D.S.; Lawton, J.H. Why More Productive Sites Have More Species: An Experimental Test of Theory Using Tree-Hole Communities. Am. Nat. 1998, 152, 510–529. [Google Scholar] [CrossRef]
- Storch, D.; Bohdalková, E.; Okie, J. The More-Individuals Hypothesis Revisited: The Role of Community Abundance in Species Richness Regulation and the Productivity-Diversity Relationship. Ecol. Lett. 2018, 21, 920–937. [Google Scholar] [CrossRef]
- Currie, D.J.; Mittelbach, G.G.; Cornell, H.V.; Field, R.; Guegan, J.-F.; Hawkins, B.A.; Kaufman, D.M.; Kerr, J.T.; Oberdorff, T.; O’Brien, E.; et al. Predictions and Tests of Climate-Based Hypotheses of Broad-Scale Variation in Taxonomic Richness. Ecol. Lett. 2004, 7, 1121–1134. [Google Scholar] [CrossRef]
- Šímová, I.; Storch, D.; Keil, P.; Boyle, B.; Phillips, O.L.; Enquist, B.J. Global Species-Energy Relationship in Forest Plots: Role of Abundance, Temperature and Species Climatic Tolerances: Global Species-Energy in Forest Plots. Glob. Ecol. Biogeogr. 2011, 20, 842–856. [Google Scholar] [CrossRef]
- Šímová, I.; Storch, D. The Enigma of Terrestrial Primary Productivity: Measurements, Models, Scales and the Diversity-Productivity Relationship. Ecography 2017, 40, 239–252. [Google Scholar] [CrossRef]
- Chase, J.M.; Leibold, M.A. Spatial Scale Dictates the Productivity—Biodiversity Relationship. Nature 2002, 416, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Cusens, J.; Wright, S.D.; McBride, P.D.; Gillman, L.N. What Is the Form of the Productivity—Animal-Species-Richness Relationship? A Critical Review and Meta-Analysis. Ecology 2012, 93, 2241–2252. [Google Scholar] [CrossRef] [PubMed]
- Lightfoot, D.C.; Whitford, W.G. Productivity of Creosotebush Foliage and Associated Canopy Arthropods Along a Desert Roadside. Am. Midl. Nat. 1991, 125, 310. [Google Scholar] [CrossRef]
- Wenninger, E.J.; Inouye, R.S. Insect Community Response to Plant Diversity and Productivity in a Sagebrush–Steppe Ecosystem. J. Arid Environ. 2008, 72, 24–33. [Google Scholar] [CrossRef]
- Bailey, S.-A.; Horner-Devine, M.C.; Luck, G.; Moore, L.A.; Carney, K.M.; Anderson, S.; Betrus, C.; Fleishman, E. Primary Productivity and Species Richness: Relationships among Functional Guilds, Residency Groups and Vagility Classes at Multiple Spatial Scales. Ecography 2004, 27, 207–217. [Google Scholar] [CrossRef]
- Seto, K.C.; Fleishman, E.; Fay, J.P.; Betrus, C.J. Linking Spatial Patterns of Bird and Butterfly Species Richness with Landsat TM Derived NDVI. Int. J. Remote Sens. 2004, 25, 4309–4324. [Google Scholar] [CrossRef]
- Kaspari, M.; Ward, P.S.; Yuan, M. Energy Gradients and the Geographic Distribution of Local Ant Diversity. Oecologia 2004, 140, 407–413. [Google Scholar] [CrossRef]
- Brasil, L.S.; Silverio, D.V.; Cabette, H.S.R.; Batista, J.D.; Vieira, T.B.; Dias-Silva, K.; de Oliveira-Junior, J.M.B.; de Carvalho, F.G.; Calvão, L.B.; Macedo, M.N.; et al. Net Primary Productivity and Seasonality of Temperature and Precipitation Are Predictors of the Species Richness of the Damselflies in the Amazon. Basic Appl. Ecol. 2019, 35, 45–53. [Google Scholar] [CrossRef]
- Kerr, J.T.; Southwood, T.R.E.; Cihlar, J. Remotely Sensed Habitat Diversity Predicts Butterfly Species Richness and Community Similarity in Canada. Proc. Natl. Acad. Sci. USA 2001, 98, 11365–11370. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.A.; Porter, E.E. Water-Energy Balance and the Geographic Pattern of Species Richness of Western Palearctic Butterflies: Water-Energy Balance and Butterfly Species Richness. Ecol. Entomol. 2003, 28, 678–686. [Google Scholar] [CrossRef]
- Vinson, M.R.; Hawkins, C.P. Broad-Scale Geographical Patterns in Local Stream Insect Genera Richness. Ecography 2003, 26, 751–767. [Google Scholar] [CrossRef]
- Ballesteros-Mejia, L.; Kitching, I.J.; Jetz, W.; Nagel, P.; Beck, J. Mapping the Biodiversity of Tropical Insects: Species Richness and Inventory Completeness of African Sphingid Moths: Mapping the Biodiversity of Tropical Insects. Glob. Ecol. Biogeogr. 2013, 22, 586–595. [Google Scholar] [CrossRef]
- Murphy, B.P.; Andersen, A.N.; Parr, C.L. The Underestimated Biodiversity of Tropical Grassy Biomes. Phil. Trans. R. Soc. B 2016, 371, 20150319. [Google Scholar] [CrossRef]
- Davis, C.L.; Vincent, K. Climate Risk and Vulnerability: A Handbook for Southern Africa; Council for Scientific and Industrial Research: Stellenbosch, South Africa, 2017. [Google Scholar]
- Delabye, S.; Sedláček, O.; Maicher, V.; Tropek, R. New Records of Six Moth (Lepidoptera: Erebidae, Lasiocampidae) Species in South African Countries, with Comments on Their Distribution. Biodivers. Data J. 2020, 8, e59339. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An Extended AVHRR 8-km NDVI Dataset Compatible with MODIS and SPOT Vegetation NDVI Data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Beck, J.; Schwanghart, W. Comparing Measures of Species Diversity from Incomplete Inventories: An Update: Measuring Diversity from Incomplete Inventories. Methods Ecol. Evol. 2010, 1, 38–44. [Google Scholar] [CrossRef]
- Levanoni, O.; Levin, N.; Pe’er, G.; Turbé, A.; Kark, S. Can We Predict Butterfly Diversity along an Elevation Gradient from Space? Ecography 2011, 34, 372–383. [Google Scholar] [CrossRef]
- Peters, M.K.; Hemp, A.; Appelhans, T.; Behler, C.; Classen, A.; Detsch, F.; Ensslin, A.; Ferger, S.W.; Frederiksen, S.B.; Gebert, F.; et al. Predictors of Elevational Biodiversity Gradients Change from Single Taxa to the Multi-Taxa Community Level. Nat. Commun. 2016, 7, 13736. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, P.; Leather, S.R. Predicting a Global Insect Apocalypse: Insect Apocalypse. Insect Conserv. Divers. 2019, 12, 263–267. [Google Scholar]
- Vagle, G.L.; McCain, C.M. Natural Population Variability May Be Masking the More-individuals Hypothesis. Ecology 2020, 101, e03035. [Google Scholar] [PubMed]
- O’Brien, E.M. Climatic Gradients in Woody Plant Species Richness: Towards an Explanation Based on an Analysis of Southern Africa’s Woody Flora. J. Biogeogr. 1993, 20, 181. [Google Scholar] [CrossRef]
- Hejda, M.; Čuda, J.; Pyšková, K.; Zambatis, G.; Foxcroft, L.C.; MacFadyen, S.; Storch, D.; Tropek, R.; Pyšek, P. Water Availability, Bedrock, Disturbance by Herbivores, and Climate Determine Plant Diversity in South-African Savanna. Sci. Rep. 2022, 12, 338. [Google Scholar] [PubMed]
- Siebert, A. Hydroclimate Extremes in Africa: Variability, Observations and Modeled Projections: Hydroclimate Extremes in Africa. Geogr. Compass 2014, 8, 351–367. [Google Scholar]
- Buckley, L.B.; Hurlbert, A.H.; Jetz, W. Broad-Scale Ecological Implications of Ectothermy and Endothermy in Changing Environments: Ectothermy and Endothermy. Glob. Ecol. Biogeogr. 2012, 21, 873–885. [Google Scholar]
- Lawton, J.H. Plant Architecture and the Diversity of Phytophagous Insects. Annu. Rev. Entomol. 1983, 28, 23–39. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal Species Diversity Driven by Habitat Heterogeneity/Diversity: The Importance of Keystone Structures: Animal Species Diversity Driven by Habitat Heterogeneity. J. Biogeogr. 2004, 31, 79–92. [Google Scholar]
- Delabye, S.; Maicher, V.; Sáfián, S.; Doležal, J.; Altman, J.; Janeček, Š.; Kobe, I.N.; Murkwe, M.; Šebek, P.; Tropek, R. Butterfly and Moth Communities Differ in Their Response to Habitat Structure in Rainforests of Mount Cameroon. Biotropica 2021, 53, 567–580. [Google Scholar]
- Novotny, V.; Drozd, P.; Miller, S.E.; Kulfan, M.; Janda, M.; Basset, Y.; Weiblen, G.D. Why Are There So Many Species of Herbivorous Insects in Tropical Rainforests? Science 2006, 313, 1115–1118. [Google Scholar]
- Adler, P.B.; Seabloom, E.W.; Borer, E.T.; Hillebrand, H.; Hautier, Y.; Hector, A.; Harpole, W.S.; O’Halloran, L.R.; Grace, J.B.; Anderson, T.M.; et al. Productivity Is a Poor Predictor of Plant Species Richness. Science 2011, 333, 1750–1753. [Google Scholar] [PubMed]
- Fraser, L.H.; Pither, J.; Jentsch, A.; Sternberg, M.; Zobel, M.; Askarizadeh, D.; Bartha, S.; Beierkuhnlein, C.; Bennett, J.A.; Bittel, A.; et al. Worldwide Evidence of a Unimodal Relationship between Productivity and Plant Species Richness. Science 2015, 349, 302–305. [Google Scholar] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Region (Code) | Elevation (m a.s.l.) | Latitude/ Longitude | Habitat Type | Max./ Mean/ Min. NDVI | Vegetation Cover (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
| All | 30 cm | 2 m | 5 m | >5 m | |||||
| Soussusvlei (S) | 760 | S 24.543° E 15.789° | Namib Desert with very scarce vegetation | 0.1119 0.1086 0.1038 | 14.6 | 9.9 | 4.1 | 0.5 | 0.0 |
| Namibgrens (F) | 1790 | S 23.643° E 16.279° | Namib Escarpment Woodland: dry savannahs and shrubby areas with scattered trees | 0.1448 0.1417 0.1383 | 16.8 | 6.8 | 5.3 | 4.7 | 0.0 |
| Khorixas (KH) | 1040 | S 20.440° E 15.215° | Angolian Mopane Woodland: mosaic of Acacia and mopane woodlands | 0.1864 0.1732 0.1593 | 45.7 | 2.8 | 22.3 | 17.7 | 2.9 |
| Windhoek (WI) | 1800 | S 22.608° E 16.773° | Namib Escarpment Woodland: dry savannahs and shrubby areas with scattered trees | 0.2276 0.1964 0.1719 | 46.9 | 26.0 | 16.2 | 4.7 | 0.0 |
| Etosha (ET) | 1120 | S 19.051° E 16.541° | Angolian Mopane Woodland: mosaics of Acacia and mopane woodlands | 0.2837 0.2412 0.2018 | 93.7 | 39.4 | 32.7 | 21.6 | 0.0 |
| Thakadu (K) | 1120 | S 21.867° E 21.697° | Kalahari Xeric Savannah: dry open savannahs, with scattered trees | 0.2979 0.2481 0.2047 | 106.0 | 35.5 | 49.5 | 20.5 | 0.5 |
| Central Kalahari (CK) | 980 | S 21.288° E 23.716° | Kalahari Acacia Woodland: mosaics of Vachellia, Baikeia and mopane woodlands, and small-leaved savannahs | 0.3487 0.2586 0.1912 | 87.7 | 26.0 | 33.5 | 23.5 | 4.7 |
| Grootfontein (GF) | 1220 | S 19.346° E 18.812° | Kalahari Acacia Woodland: mosaics of Vachellia, Baikeia and mopane woodlands, and small-leaved savannahs | 0.3554 0.2884 0.2264 | 133.2 | 55.0 | 45.0 | 27.5 | 5.7 |
| Bwabwata (BW) | 1030 | S 18.092° E 21.686° | Zambezian Baikiaea Woodlands: mosaic of mopane and Baikeia woodlands, and secondary grasslands | 0.4459 0.3542 0.2689 | 114.6 | 39.5 | 40.3 | 12.5 | 22.3 |
| Hwange (SI) | 1020 | S 18.699° E 26.192° | Zambezian and Mopane Woodlands: mosaic of miombo and mopane woodlands, and shrubby savannahs | 0.5468 0.4090 0.2734 | 106.1 | 42.3 | 35.5 | 18.0 | 10.3 |
| Victoria Falls (VF) | 920 | S 17.872° E 25.721° | Zambezian and Mopane Woodlands: mosaic of mopane and Baikeia woodlands, and secondary grasslands | 0.5431 0.4222 0.3066 | 123.8 | 49.0 | 41.5 | 25.2 | 8.1 |
| Chizarira (XI) | 1010 | S 17.701° E 27.855° | Zambezian and Mopane Woodlands: mosaic of mopane and Baikeia woodlands, and secondary grasslands | 0.5692 0.4379 0.3145 | 124.2 | 45.2 | 27.2 | 32.0 | 15.1 |
| Region | All Moths Exc. Geometroidea | Bombycoidea | Noctuoidea | Erebidae | Noctuidae | All Moths Incl. Geometroidea | Geometroidea | ||||||||||||||
| γ | α | Ab. | γ | α | Ab. | γ | α | Ab. | γ | α | Ab. | γ | α | Ab. | Ab. | Ab. | |||||
| Soussusvlei | 10 | 1.9 | 47 | 1 | 0.1 | 1 | 9 | 1.8 | 46 | 3 | 0.8 | 23 | 6 | 1.0 | 23 | 50 | 3 | ||||
| Namibgrens | 45 | 9.7 | 1331 | 2 | 0.3 | 5 | 41 | 9.2 | 1324 | 13 | 3.1 | 57 | 28 | 6.1 | 1267 | 1532 | 201 | ||||
| Khorixas | 45 | 9.0 | 382 | 2 | 0.2 | 2 | 43 | 8.8 | 380 | 11 | 1.6 | 20 | 32 | 7.2 | 360 | 438 | 56 | ||||
| Windhoek | 32 | 7.0 | 399 | 3 | 0.4 | 4 | 28 | 6.5 | 394 | 11 | 3.2 | 195 | 17 | 3.3 | 199 | 436 | 37 | ||||
| Etosha | 39 | 9.2 | 710 | 0 | 0.0 | 0 | 39 | 9.2 | 710 | 13 | 2.8 | 50 | 25 | 5.4 | 312 | 876 | 166 | ||||
| Thakadu | 50 | 10.1 | 350 | 6 | 1.5 | 27 | 44 | 8.6 | 323 | 11 | 3.1 | 64 | 32 | 5.3 | 254 | 404 | 54 | ||||
| Central Kalahari | 74 | 18.4 | 569 | 8 | 2.5 | 54 | 66 | 15.9 | 515 | 19 | 4.3 | 69 | 47 | 11.6 | 446 | 612 | 43 | ||||
| Grootfontein | 85 | 21.3 | 1337 | 15 | 4.9 | 161 | 70 | 16.4 | 1176 | 16 | 4.4 | 696 | 50 | 10.8 | 325 | 3078 | 1741 | ||||
| Bwabwata | 92 | 23.0 | 762 | 19 | 4.8 | 149 | 71 | 17.9 | 610 | 30 | 8.0 | 399 | 39 | 9.7 | 209 | 982 | 220 | ||||
| Hwange | 145 | 36.0 | 994 | 9 | 3.3 | 124 | 125 | 27.8 | 669 | 49 | 9.6 | 249 | 73 | 17.4 | 406 | 1232 | 238 | ||||
| Victoria Falls | 179 | 44.8 | 1757 | 15 | 4.1 | 63 | 152 | 36.5 | 1574 | 71 | 16.1 | 340 | 72 | 17.6 | 1121 | 2182 | 425 | ||||
| Chizarira | 99 | 19.2 | 410 | 11 | 3.7 | 98 | 80 | 13.4 | 203 | 35 | 6.2 | 102 | 35 | 4.7 | 62 | 550 | 140 | ||||
| Region | Eutellidae | Lasiocampidae | Limacodidae | Notodontidae | Saturniidae | Sphingidae | Eupterotidae | ||||||||||||||
| γ | α | Ab. | γ | α | Ab. | γ | α | Ab. | γ | α | Ab. | γ | α | Ab. | γ | α | Ab. | γ | α | Ab. | |
| Soussusvlei | 0 | 0 | 0 | 1 | 0.1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Namibgrens | 0 | 0 | 0 | 2 | 0.3 | 5 | 2 | 0.2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Khorixas | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0.1 | 1 | 1 | 0.1 | 1 | 0 | 0 | 0 |
| Windhoek | 0 | 0 | 0 | 2 | 0.2 | 2 | 1 | 0.1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0.2 | 2 |
| Etosha | 1 | 1.0 | 348 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Thakadu | 0 | 0 | 0 | 2 | 0.9 | 18 | 0 | 0 | 0 | 1 | 0.2 | 5 | 1 | 0.1 | 1 | 2 | 0.3 | 4 | 1 | 0.2 | 4 |
| Central Kalahari | 0 | 0 | 0 | 4 | 1.5 | 37 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0.1 | 1 | 3 | 0.9 | 16 | 0 | 0 | 0 |
| Grootfontein | 1 | 0.1 | 1 | 6 | 2.1 | 94 | 0 | 0 | 0 | 3 | 1.1 | 154 | 1 | 0.2 | 2 | 8 | 2.6 | 65 | 0 | 0 | 0 |
| Bwabwata | 1 | 0.1 | 1 | 7 | 1.5 | 47 | 2 | 0.3 | 3 | 1 | 0.1 | 1 | 4 | 1.0 | 26 | 7 | 1.9 | 67 | 1 | 0.4 | 9 |
| Hwange | 0 | 0 | 0 | 2 | 0.3 | 3 | 11 | 4.9 | 201 | 3 | 0.8 | 14 | 1 | 0.2 | 2 | 4 | 1.3 | 44 | 2 | 1.5 | 75 |
| Victoria Falls | 0 | 0 | 0 | 5 | 1.1 | 11 | 12 | 4.2 | 120 | 9 | 2.8 | 113 | 2 | 0.2 | 2 | 5 | 1.9 | 35 | 3 | 0.9 | 15 |
| Chizarira | 0 | 0 | 0 | 5 | 1.6 | 60 | 8 | 2.1 | 109 | 10 | 2.5 | 39 | 1 | 0.4 | 5 | 2 | 0.4 | 4 | 3 | 1.3 | 29 |
| Alpha Diversity | Gamma Diversity | Abundance | ||||
|---|---|---|---|---|---|---|
| Linear | Unimodal | Linear | Unimodal | Linear | Unimodal | |
| All moths exc. Geometroidea | 0.68 *** | 0.76 n.s. | 0.75 *** | 0.77 n.s. | 0.19 n.s. | 0.26 n.s. |
| All moths incl. Geometroidea | - | - | - | - | 0.20 n.s. | 0.29 n.s. |
| Geometroidea | - | - | - | - | 0.28 * | 0.34 n.s. |
| Bombycoidea | 0.63 ** | 0.66 n.s. | 0.51 ** | 0.47 n.s. | 0.57 ** | 0.56 n.s. |
| Noctuoidea | 0.60 ** | 0.69 n.s. | 0.71 *** | 0.73 n.s. | 0.03 n.s. | 0.15 n.s. |
| Erebidae | 0.63 ** | 0.77 n.s. | 0.82 *** | 0.80 n.s. | 0.37 * | 0.40 n.s. |
| Noctuidae | 0.39 * | 0.45 n.s. | 0.51 ** | 0.60 n.s. | −0.08 n.s. | −0.05 n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delabye, S.; Storch, D.; Sedláček, O.; Albrecht, T.; Hořák, D.; Maicher, V.; Tószögyová, A.; Tropek, R. Moth Diversity Increases along a Continent-Wide Gradient of Environmental Productivity in South African Savannahs. Insects 2022, 13, 778. https://doi.org/10.3390/insects13090778
Delabye S, Storch D, Sedláček O, Albrecht T, Hořák D, Maicher V, Tószögyová A, Tropek R. Moth Diversity Increases along a Continent-Wide Gradient of Environmental Productivity in South African Savannahs. Insects. 2022; 13(9):778. https://doi.org/10.3390/insects13090778
Chicago/Turabian StyleDelabye, Sylvain, David Storch, Ondřej Sedláček, Tomáš Albrecht, David Hořák, Vincent Maicher, Anna Tószögyová, and Robert Tropek. 2022. "Moth Diversity Increases along a Continent-Wide Gradient of Environmental Productivity in South African Savannahs" Insects 13, no. 9: 778. https://doi.org/10.3390/insects13090778
APA StyleDelabye, S., Storch, D., Sedláček, O., Albrecht, T., Hořák, D., Maicher, V., Tószögyová, A., & Tropek, R. (2022). Moth Diversity Increases along a Continent-Wide Gradient of Environmental Productivity in South African Savannahs. Insects, 13(9), 778. https://doi.org/10.3390/insects13090778