
Copper Chaperone for Superoxide Dismutase FoCCS1 in Frankliniella occidentalis May Be Associated with Feeding Adaptation after Host Shifting

 , , and
, , and
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
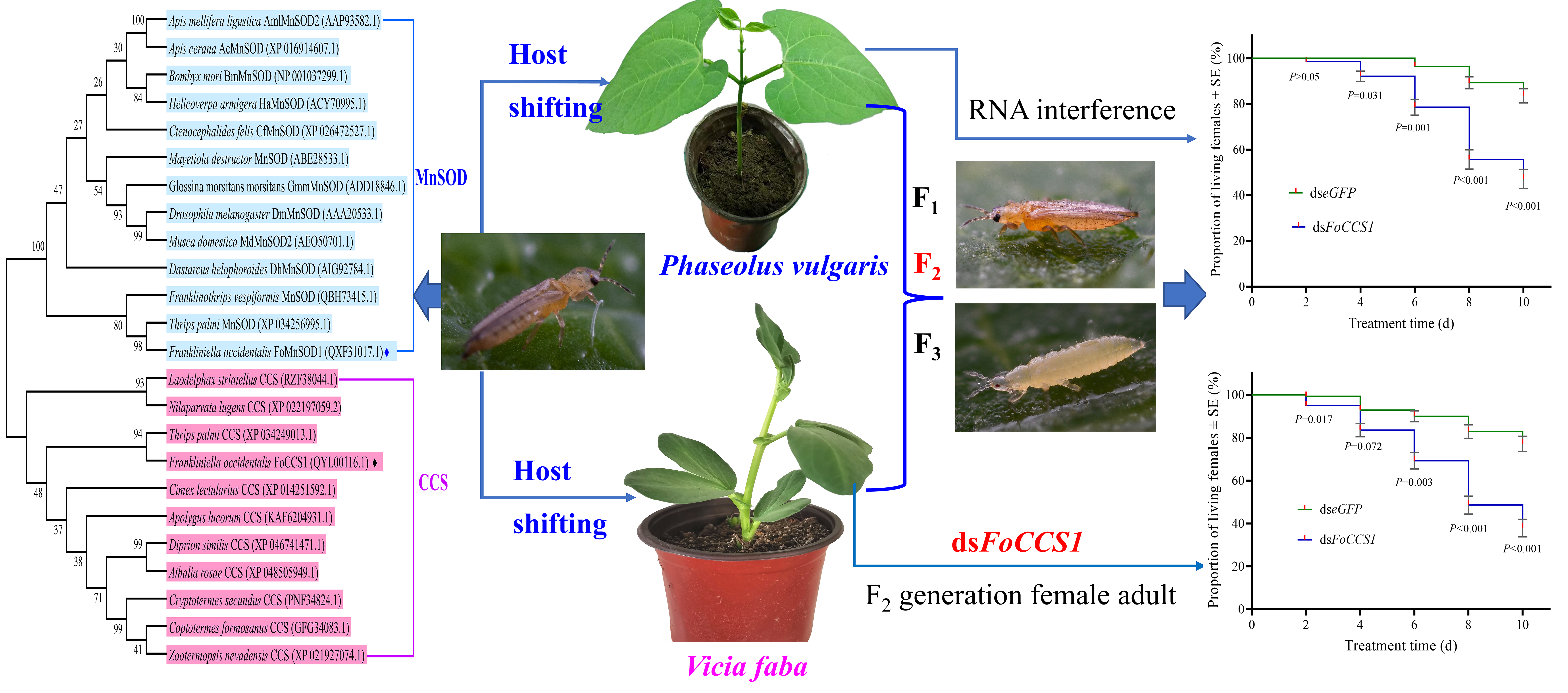
2.1. Insect Culture and Host Plant Cultivation
2.2. Reverse-Transcription PCR and Cloning by Rapid Amplification of the cDNA Ends of the F. occidentalis SOD Gene
2.3. Bioinformatics Analysis of the SOD Gene Sequence
2.4. Expression Profiles of FoCCS1 and FoMnSOD2 at the Different Developmental Stages of F. occidentalis
2.5. Effects of Host Shifting on the Expression Levels of FoCCS1 and FoMnSOD2 in F. occidentalis
2.6. Synthesis of FoCCS1 Double-Stranded RNA
2.7. FoCCS1 RNA Interference and Determination of SOD Enzyme Activity
2.8. Effects of RNAi on the Survival Rate of Adult Females and the Number of Offspring Nymphs of F. occidentalis
2.9. Statistical Analyses
3. Results
3.1. Sequence Characterization of FoCCS1 and FoMnSOD2
3.2. Homologous Alignment and Phylogenetic Analysis
3.3. Analysis of Expression Profiles in the Different Developmental Stages
3.4. Expression of FoCCS1 in F. occidentalis Feeding on Different Hosts
3.5. Expression of FoMnSOD2 in F. occidentalis Feeding on Different Hosts
3.6. FoCCS1 Expression Level and SOD Activity Downregulated by RNAi
3.7. RNAi Reduced the Survival Rate and Fecundity of Adult F. occidentalis Females
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reitz, S.R.; Gao, Y.L.; Kirk, W.D.J.; Hoddle, M.S.; Leiss, K.A.; Funderburk, J.E. Invasion biology, ecology, and management of western flower thrips. Annu. Rev. Entomol. 2020, 65, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Reitz, S.R. Biology and ecology of the western flower thrips (Thysanoptera: Thripidae): The making of a pest. Flarida Entomol. 2009, 92, 7–13. [Google Scholar] [CrossRef]
- Zhang, B.; Qian, W.Q.; Qiao, X.; Xi, Y.; Wan, F.H. Invasion biology, ecology, and management of Frankliniella occidentalis in China. Arch. Insect Biochem. Physiol. 2019, 102, e21613. [Google Scholar] [CrossRef] [PubMed]
- Mouden, S.; Sarmiento, K.F.; Klinkhamer, P.G.; Leiss, K.A. Integrated pest management in western flower thrips: Past, present and future. Pest Manag. Sci. 2017, 73, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Zhi, J.R.; Gai, H.T. Effects of host plants and temperature on Frankliniella occidentalis growth and development. Chin. J. Ecol. 2011, 30, 558–563. [Google Scholar] [CrossRef]
- Mason, P.A. On the role of host phenotypic plasticity in host shifting by parasites. Ecol. Lett. 2016, 19, 121–132. [Google Scholar] [CrossRef]
- Forbes, A.A.; Devine, S.N.; Hippee, A.C.; Tvedte, E.S.; Ward, A.; Widmayer, H.A.; Wilson, C.J. Revisiting the particular role of host shifts in initiating insect speciation. Evolution 2017, 71, 1126–1137. [Google Scholar] [CrossRef]
- Cui, N.; Yang, P.C.; Guo, K.; Kang, L.; Cui, F. Large-scale gene expression reveals different adaptations of Hyalopterus persikonus to winter and summer host plants. Insect Sci. 2017, 24, 431–442. [Google Scholar] [CrossRef]
- Erb, M.; Reymond, P. Molecular interactions between plants and insect herbivores. Annu. Rev. Plant. Biol. 2019, 70, 527–557. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Maffei, M.E.; Mithöfer, A.; Boland, W. Insects feeding on plants: Rapid signals and responses preceding the induction of phytochemical release. Phytochemistry 2007, 68, 2946–2959. [Google Scholar] [CrossRef]
- Torres, M.A. ROS in biotic interactions. Physiol. Plant 2010, 138, 414–429. [Google Scholar] [CrossRef]
- Kant, M.R.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.C.J.; Villarroel, C.A.; Ataide, L.M.S.; Dermauw, W.; Glas, J.J.; et al. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef]
- Argandoña, V.H.; Chaman, M.; Cardemil, L.; Muñoz, O.; Zúñiga, G.E.; Corcuera, L.J. Ethylene production and peroxidase activity in aphid-infested barley. J. Chem. Ecol. 2001, 27, 53–68. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; War, M.Y.; Ignacimuthu, S. Jasmonic acid-mediated-induced resistance in groundnut (Arachis hypogaea L.) against Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae). J. Plant Growth Regul. 2011, 30, 512–523. [Google Scholar] [CrossRef]
- Van Raamsdonk, J.M.; Hekimi, S. Superoxide dismutase is dispensable for normal animal lifespan. Proc. Natl. Acad. Sci. USA 2012, 109, 5785–5790. [Google Scholar] [CrossRef]
- Colinet, D.; Cazes, D.; Belghazi, M.; Gatti, J.L.; Poirié, M. Extracellular superoxide dismutase in insects: Characterization, function, and interspecific variation in parasitoid wasp venom. J. Biol. Chem. 2011, 286, 40110–40121. [Google Scholar] [CrossRef]
- Corona, M.; Robinson, G.E. Genes of the antioxidant system of the honey bee: Annotation and phylogeny. Insect Mol. Biol. 2006, 15, 687–701. [Google Scholar] [CrossRef]
- Price, D.L.; Rothstein, J.; Gitlin, J.D. Copper chaperone for superoxide dismutase is essential to activate mammalian Cu/Zn superoxide dismutase. Proc. Natl. Acad. Sci. USA 2000, 97, 2886–2891. [Google Scholar] [CrossRef]
- Schmidt, P.J.; Ramos-Gomez, M.; Culotta, V.C. A gain of superoxide dismutase (SOD) activity obtained with CCS, the copper metallochaperone for SOD1. Biol. Chem. 1999, 274, 36952–36956. [Google Scholar] [CrossRef] [Green Version]
- Miyayama, T.; Ishizuka, Y.; Iijima, T.; Hiraoka, D.; Ogra, Y. Roles of copper chaperone for superoxide dismutase 1 and metallothionein in copper homeostasis. Metallomics 2011, 3, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Boyd, S.D.; Calvo, J.S.; Liu, L.; Ullrich, M.S.; Skopp, A.; Meloni, G.; Winkler, D.D. The yeast copper chaperone for copper-zinc superoxide dismutase (CCS1) is a multifunctional chaperone promoting all levels of SOD1 maturation. J. Biol. Chem. 2019, 294, 1956–1966. [Google Scholar] [CrossRef] [PubMed]
- Niedzwiecki, A.; Reveillaud, I.; Fleming, J.E. Changes in superoxide dismutase and catalase in aging heat-shocked Drosophila. Free Radic. Res. Commun. 1992, 17, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Gao, Y.F.; Liu, Y.J.; Ge, X.D.; Li, X.; Li, F.S. Identification of the gene MdSOD3 and its role in resisting heavy metal stress in housefly (Musca domestica). Acta Entomol. Sin. 2012, 55, 267–275. [Google Scholar] [CrossRef]
- Liu, L.; Hou, X.L.; Yue, W.B.; Xie, W.; Zhang, T.; Zhi, J.R. Response of protective enzymes in western flower thrips (Thysanoptera: Thripidae) to two leguminous plants. Environ. Entomol. 2020, 49, 1191–1197. [Google Scholar] [CrossRef]
- Dietz, K.J.; Mittler, R.; Noctor, G. Recent progress in understanding the role of reactive oxygen species in plant cell signaling. Plant Physiol. 2016, 171, 1535–1539. [Google Scholar] [CrossRef]
- Mhamdi, A.; Van Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef]
- Zheng, Y.T.; Li, H.B.; Lu, M.X.; Du, Y.Z. Evaluation and validation of reference genes for qRT-PCR normalization in Frankliniella occidentalis (Thysanoptera: Thripidae). PLoS ONE 2014, 9, e111369. [Google Scholar] [CrossRef]
- Zhang, T.; Zhi, J.R.; Li, D.Y.; Liu, L.; Zeng, G. Effect of different double-stranded RNA feeding solutions on the RNA interference of V-ATPase-B in Frankliniella occidentalis. Entomol. Exp. Appl. 2022, 170, 427–436. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bannister, J.V.; Bannister, W.H.; Rotilio, G. Aspects of the structure, function, and applications of superoxide dismutase. CRC Crit. Rev. Biochem. 1987, 22, 111–180. [Google Scholar] [CrossRef]
- Zhu, J.J.; Jiang, F.; Wang, X.H.; Yang, P.C.; Bao, Y.Y.; Zhao, W.; Wang, W.; Lu, H.; Wang, Q.S.; Cui, N.; et al. Genome sequence of the small brown planthopper, Laodelphax striatellus. GigaScience 2017, 6, 1–12. [Google Scholar] [CrossRef]
- Itakura, S.; Yoshikawa, Y.; Togami, Y.; Umezawa, K. Draft genome sequence of the termite, Coptotermes formosanus: Genetic insights into the pyruvate dehydrogenase complex of the termite. J. Asia-Pac. Entomol. 2020, 23, 666–674. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, H.W.; Wang, H.; Huang, T.Y.; Liu, B.; Yang, B.; Yin, L.J.; Li, B.; Zhang, Y.; Zhang, S.; et al. Apolygus lucorum genome provides insights into omnivorousness and mesophyll feeding. Mol. Ecol. Resour. 2021, 21, 287–300. [Google Scholar] [CrossRef]
- Brandt, A.; Bast, J.; Scheu, S.; Meusemann, K.; Donath, A.; Schütte, K.; Machida, R.; Kraaijeveld, K. No signal of deleterious mutation accumulation in conserved gene sequences of extant asexual hexapods. Sci. Rep. 2019, 9, 5338. [Google Scholar] [CrossRef]
- Park, S.Y.; Nair, P.M.; Choi, J. Characterization and expression of superoxide dismutase genes in Chironomus riparius (Diptera, Chironomidae) larvae as a potential biomarker of ecotoxicity. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 156, 187–194. [Google Scholar] [CrossRef]
- Blackney, M.J.; Cox, R.; Shepherd, D.; Parker, J.D. Cloning and expression analysis of Drosophila extracellular Cu Zn superoxide dismutase. Biosci. Rep. 2014, 34, e00164. [Google Scholar] [CrossRef]
- Li, X.H.; Wang, D.J.; Lei, Z.R.; Wang, H.H. Comparison of life tables for experimental populations of individual-rearing and group-rearing Frankliniella occidentalis. Sci. Agric. Sin. 2021, 54, 959–968. [Google Scholar] [CrossRef]
- Deepashree, S.; Shivanandappa, T.; Ramesh, S.R. Genetic repression of the antioxidant enzymes reduces the lifespan in Drosophila melanogaster. J. Comp. Physiol. B 2022, 192, 1–13. [Google Scholar] [CrossRef]
- Gao, X.L.; Li, J.M.; Xu, H.X.; Yan, G.H.; Jiu, M.; Liu, S.S.; Wang, X.W. Cloning of a putative extracellular Cu/Zn superoxide dismutase and functional differences of superoxide dismutases in invasive and indigenous whiteflies. Insect Sci. 2015, 22, 52–64. [Google Scholar] [CrossRef]
- Gao, X.L.; Li, J.M.; Wang, Y.L.; Jiu, M.; Yan, G.H.; Liu, S.S.; Wang, X.W. Cloning, expression and characterization of mitochondrial manganese superoxide dismutase from the Whitefly, Bemisia tabaci. Int. J. Mol. Sci. 2013, 14, 871–887. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, P.; Escudero-Martinez, C.M.; Eves-van den Akker, S.; Bos, J. Transcriptional changes in the aphid species Myzus cerasi under different host and environmental conditions. Insect Mol. Biol. 2020, 29, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Li, J.M.; Su, Y.L.; Gao, X.L.; He, J.; Liu, S.S.; Wang, X.W. Molecular characterization and oxidative stress response of an intracellular Cu/Zn superoxide dismutase (CuZnSOD) of the whitefly, Bemisia tabaci. Arch. Insect Biochem. Physiol. 2011, 77, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Ansenberger-Fricano, K.; Ganini, D.; Mao, M.; Chatterjee, S.; Dallas, S.; Mason, R.P.; Stadler, K.; Santos, J.H.; Bonini, M.G. The peroxidase activity of mitochondrial superoxide dismutase. Free Radic. Biol. Med. 2013, 54, 116–124. [Google Scholar] [CrossRef]
- Phillips, J.P.; Campbell, S.D.; Michaud, D.; Charbonneau, M.; Hilliker, A.J. Null mutation of copper/zinc superoxide dismutase in Drosophila confers hypersensitivity to paraquat and reduced longevity. Proc. Natl. Acad. Sci. USA 1989, 86, 2761–2765. [Google Scholar] [CrossRef]
- Duttaroy, A.; Paul, A.; Kundu, M.; Belton, A. A Sod2 null mutation confers severely reduced adult life span in Drosophila. Genetics 2003, 165, 2295–2299. [Google Scholar] [CrossRef]
- Kirby, K.; Hu, J.; Hilliker, A.J.; Phillips, J.P. RNA interference-mediated silencing of Sod2 in Drosophila leads to early adult-onset mortality and elevated endogenous oxidative stress. Proc. Natl. Acad. Sci. USA 2002, 99, 16162–16167. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | ORF Finder | Number of Amino Acids | Molecular Weight | Theoretical pI | Phosphorylation Site | Signature |
|---|---|---|---|---|---|---|
| FoCCS1 | 912 bp | 303 | 31,978.98 | 5.84 | 2 (S), 0 (T), 0 (Y) | GNSGhRlACgiI |
| FoMnSOD2 | 678 bp | 226 | 25,514.35 | 8.43 | 1 (S), 2 (T), 2 (Y) | DvWEHAYY |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Liu, L.; Zhi, J.-R.; Jia, Y.-L.; Yue, W.-B.; Zeng, G.; Li, D.-Y. Copper Chaperone for Superoxide Dismutase FoCCS1 in Frankliniella occidentalis May Be Associated with Feeding Adaptation after Host Shifting. Insects 2022, 13, 782. https://doi.org/10.3390/insects13090782
Zhang T, Liu L, Zhi J-R, Jia Y-L, Yue W-B, Zeng G, Li D-Y. Copper Chaperone for Superoxide Dismutase FoCCS1 in Frankliniella occidentalis May Be Associated with Feeding Adaptation after Host Shifting. Insects. 2022; 13(9):782. https://doi.org/10.3390/insects13090782
Chicago/Turabian StyleZhang, Tao, Li Liu, Jun-Rui Zhi, Yu-Lian Jia, Wen-Bo Yue, Guang Zeng, and Ding-Yin Li. 2022. "Copper Chaperone for Superoxide Dismutase FoCCS1 in Frankliniella occidentalis May Be Associated with Feeding Adaptation after Host Shifting" Insects 13, no. 9: 782. https://doi.org/10.3390/insects13090782
APA StyleZhang, T., Liu, L., Zhi, J. -R., Jia, Y. -L., Yue, W. -B., Zeng, G., & Li, D. -Y. (2022). Copper Chaperone for Superoxide Dismutase FoCCS1 in Frankliniella occidentalis May Be Associated with Feeding Adaptation after Host Shifting. Insects, 13(9), 782. https://doi.org/10.3390/insects13090782