

Evidence for Reconsidering the Taxonomic Status of Closely Related Oligonychus Species in punicae Complex (Acari: Prostigmata: Tetranychidae)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
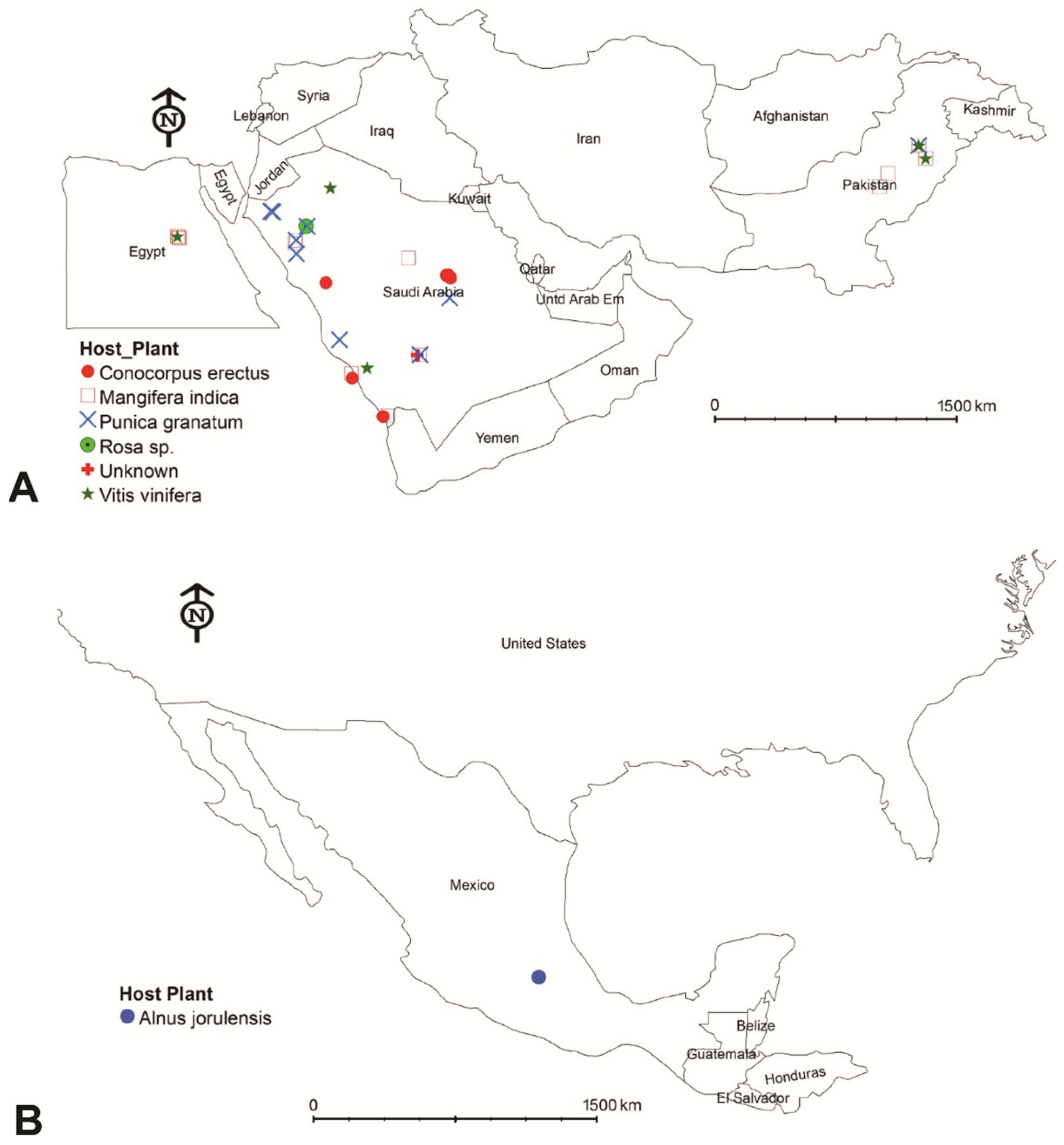
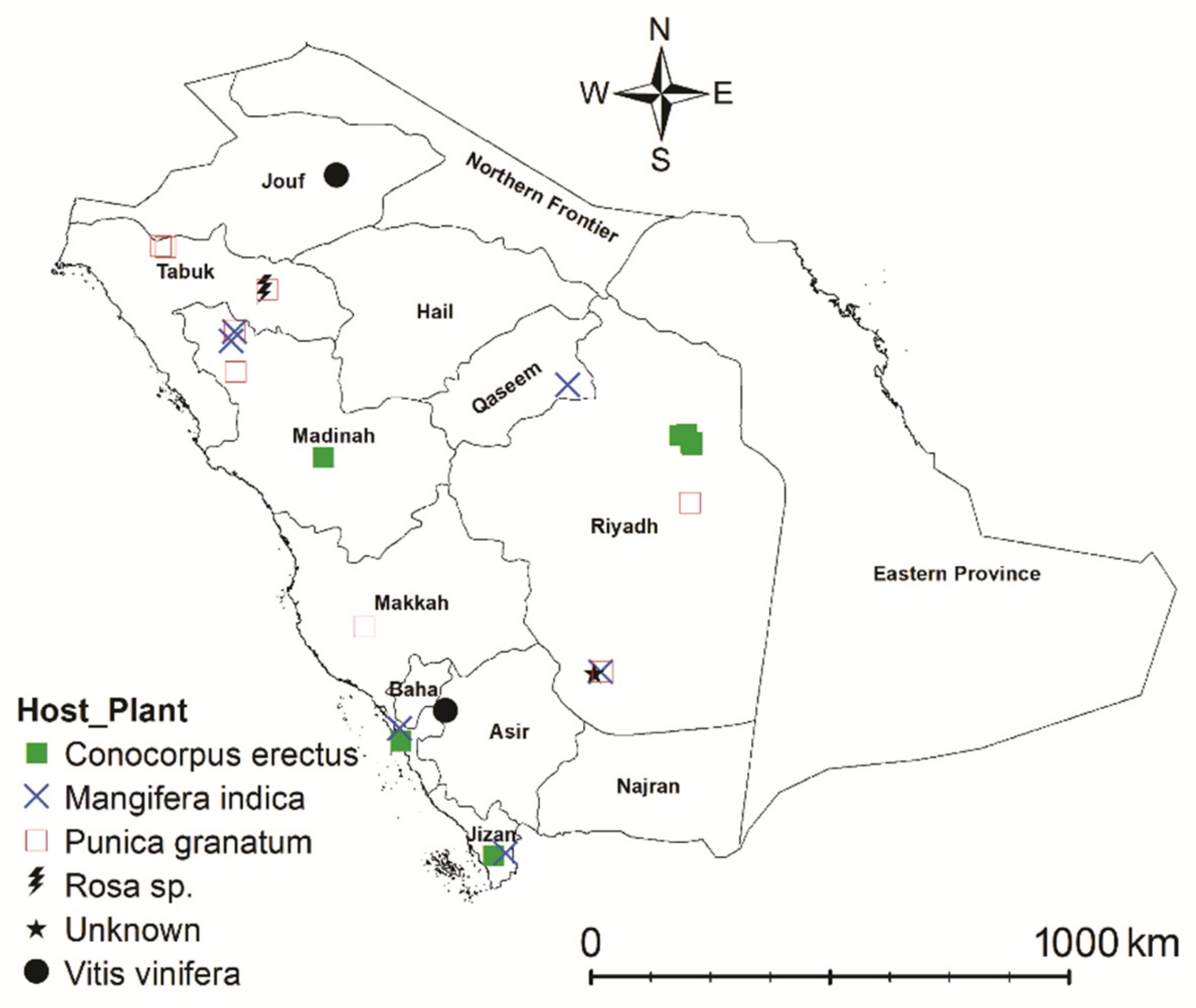
2.1. Spider Mite Samples Collection, Preservation, and Processing
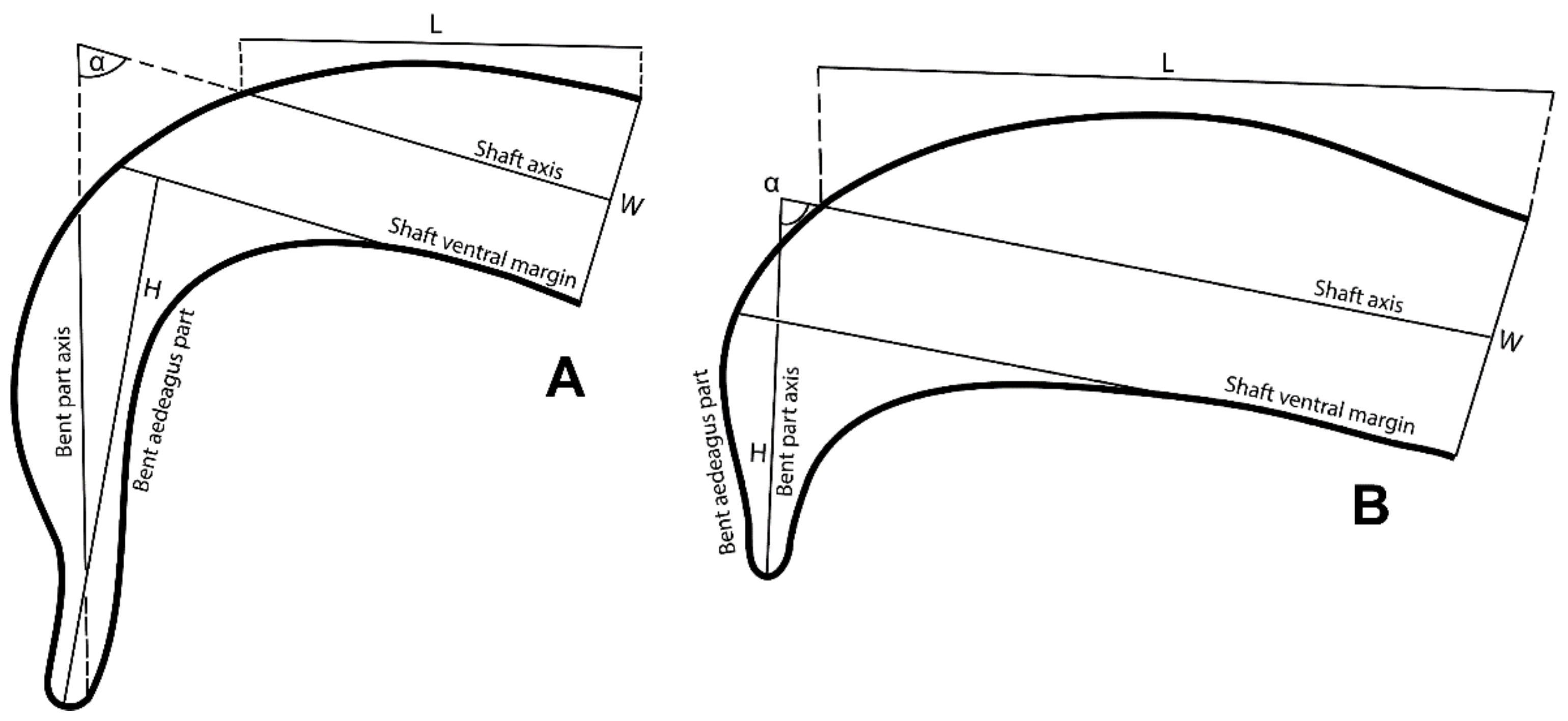
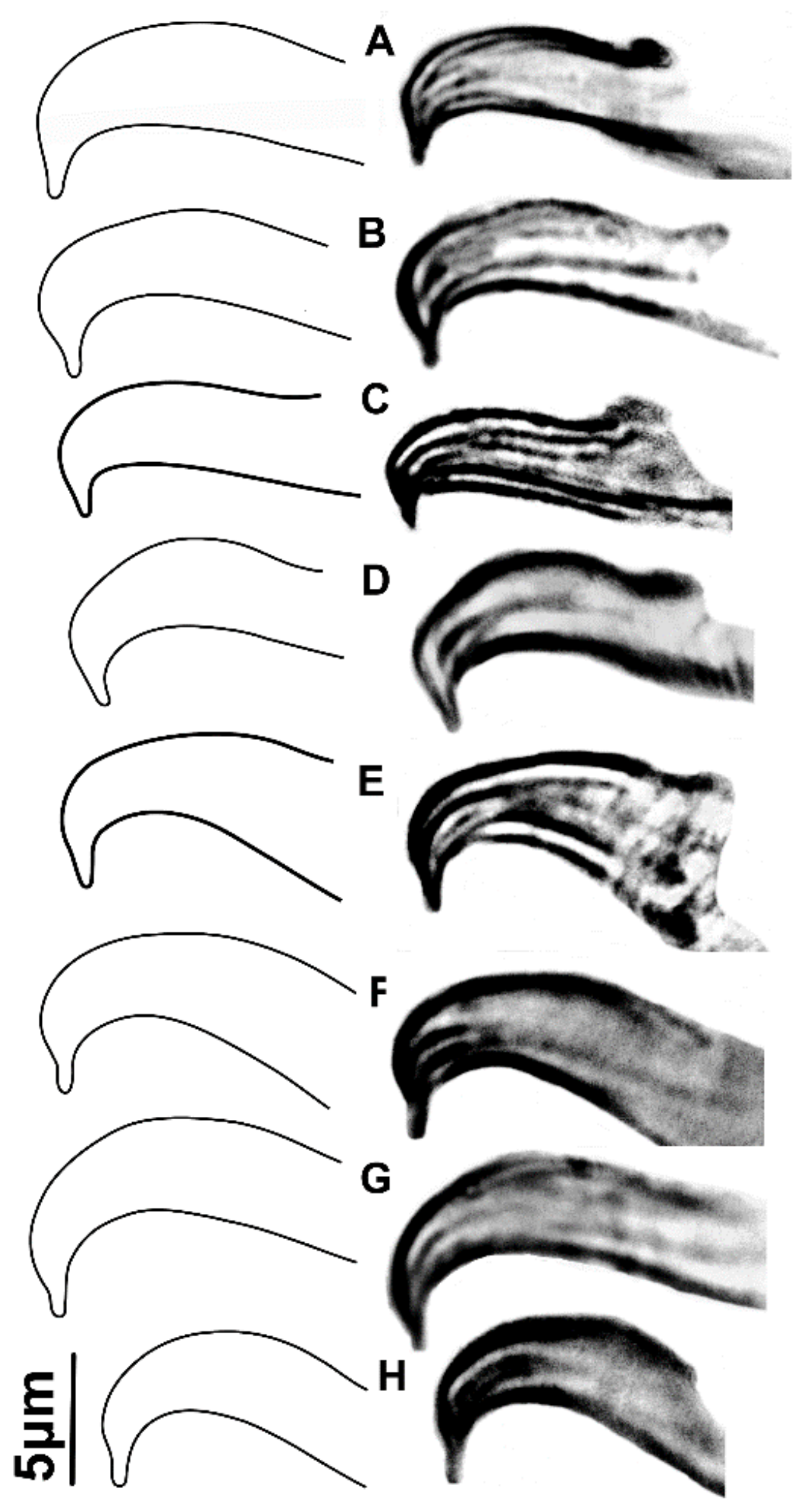
2.2. Morphological Study
2.3. Molecular Study
2.3.1. DNA Extraction and Amplification of ITS2 and COI Regions
2.3.2. DNA Sequencing and Analysis
2.3.3. Phylogenetic and Genetic Distances Analysis
3. Results
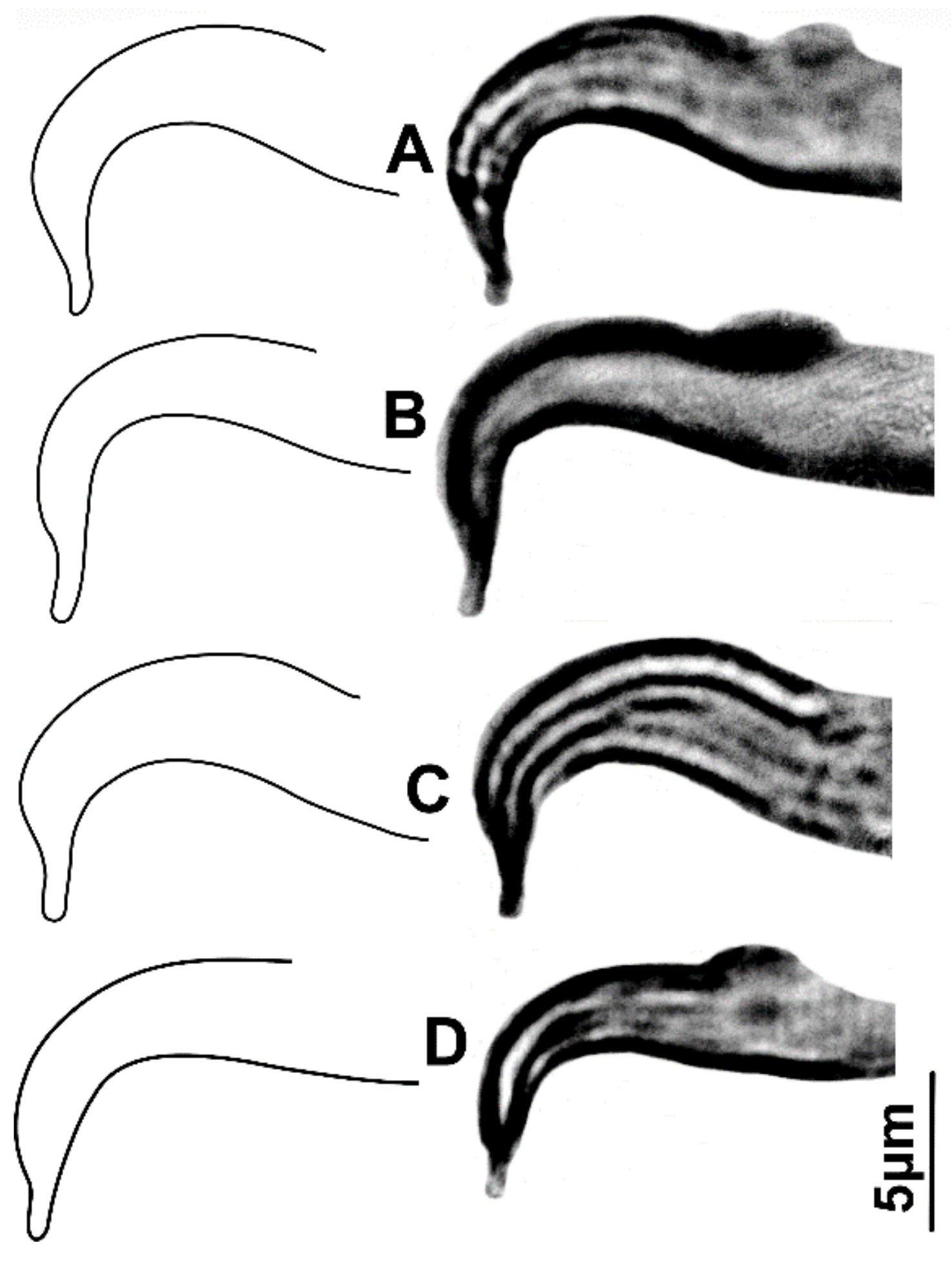
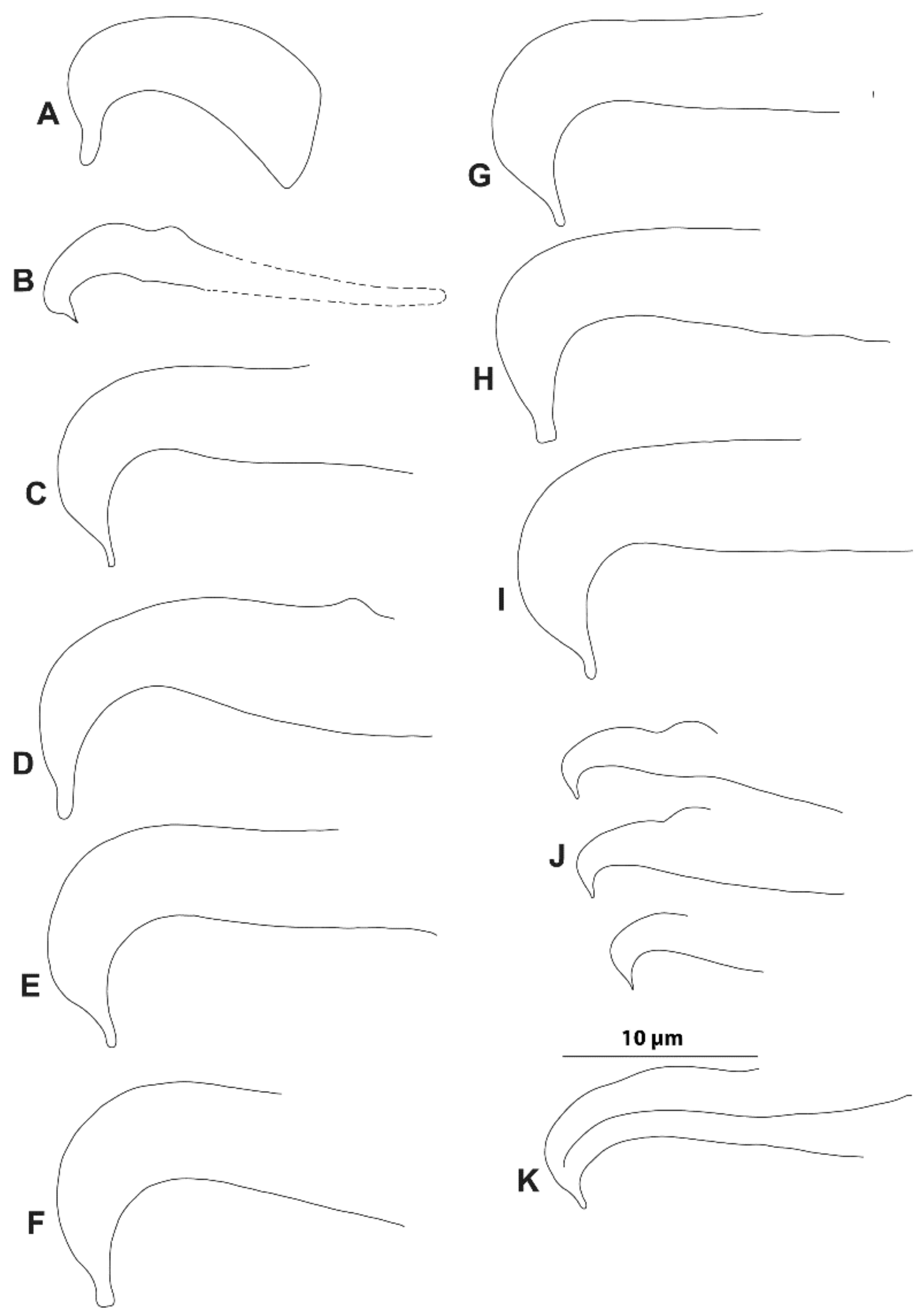
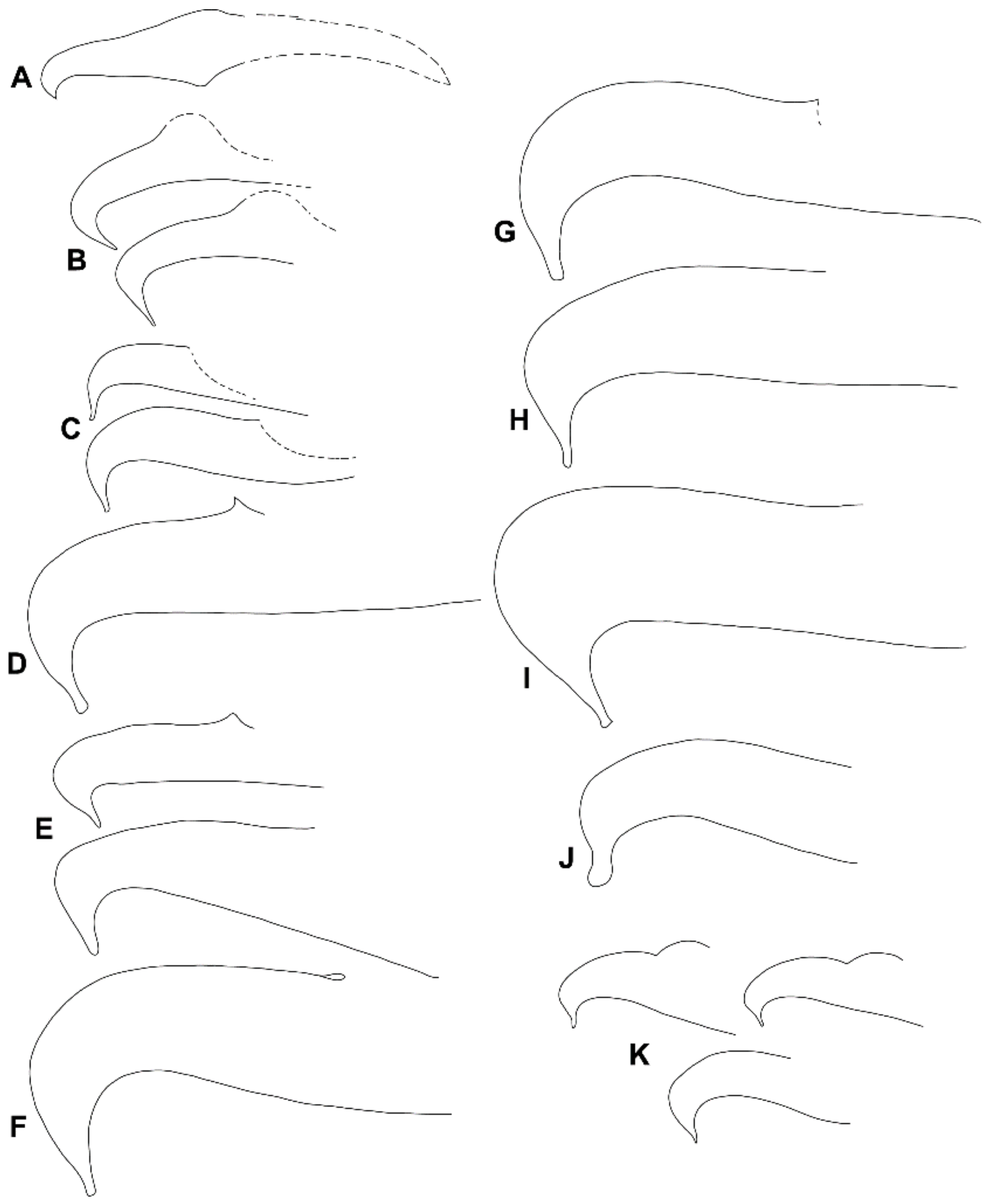
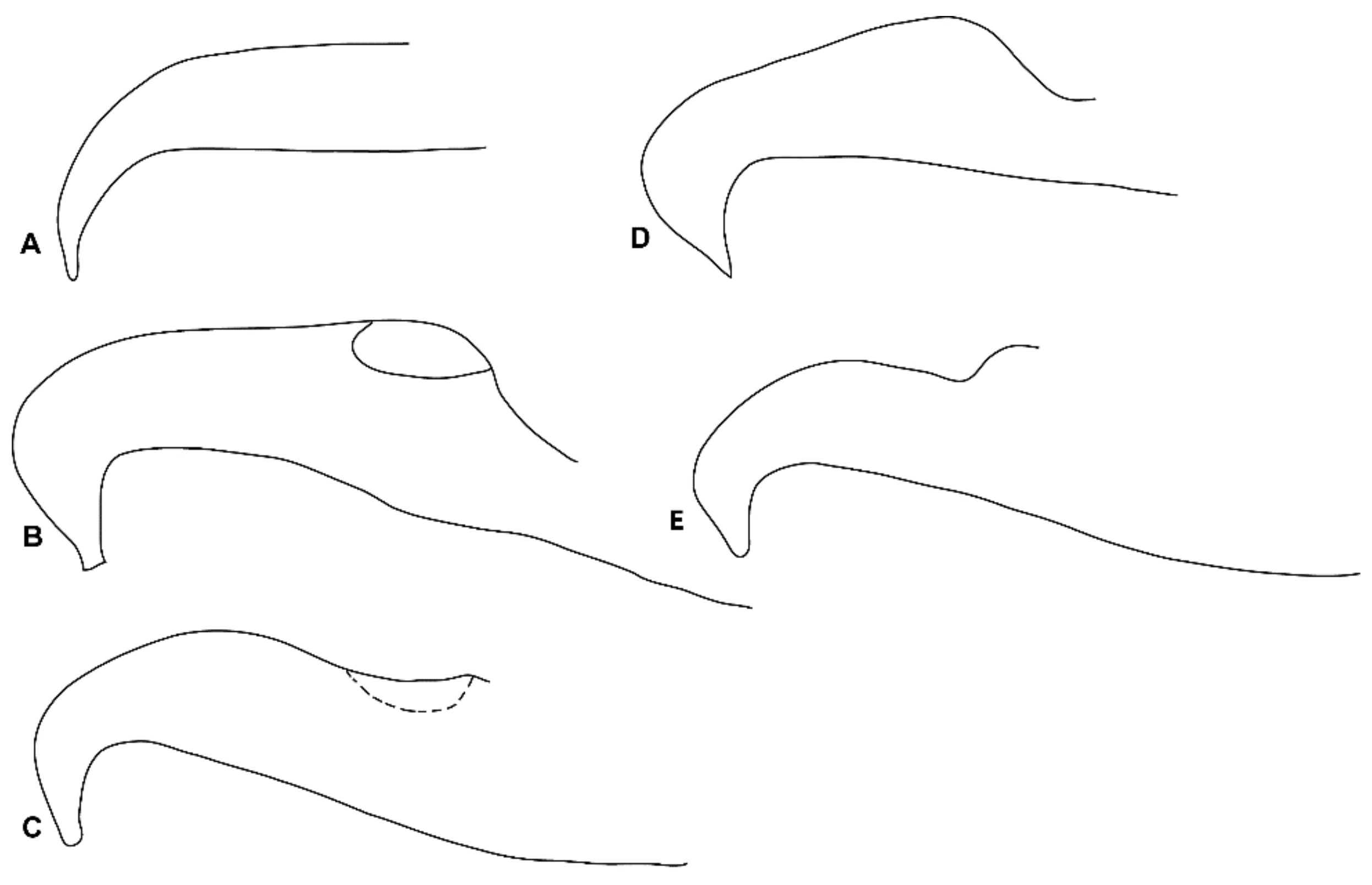
3.1. Morphological Analysis
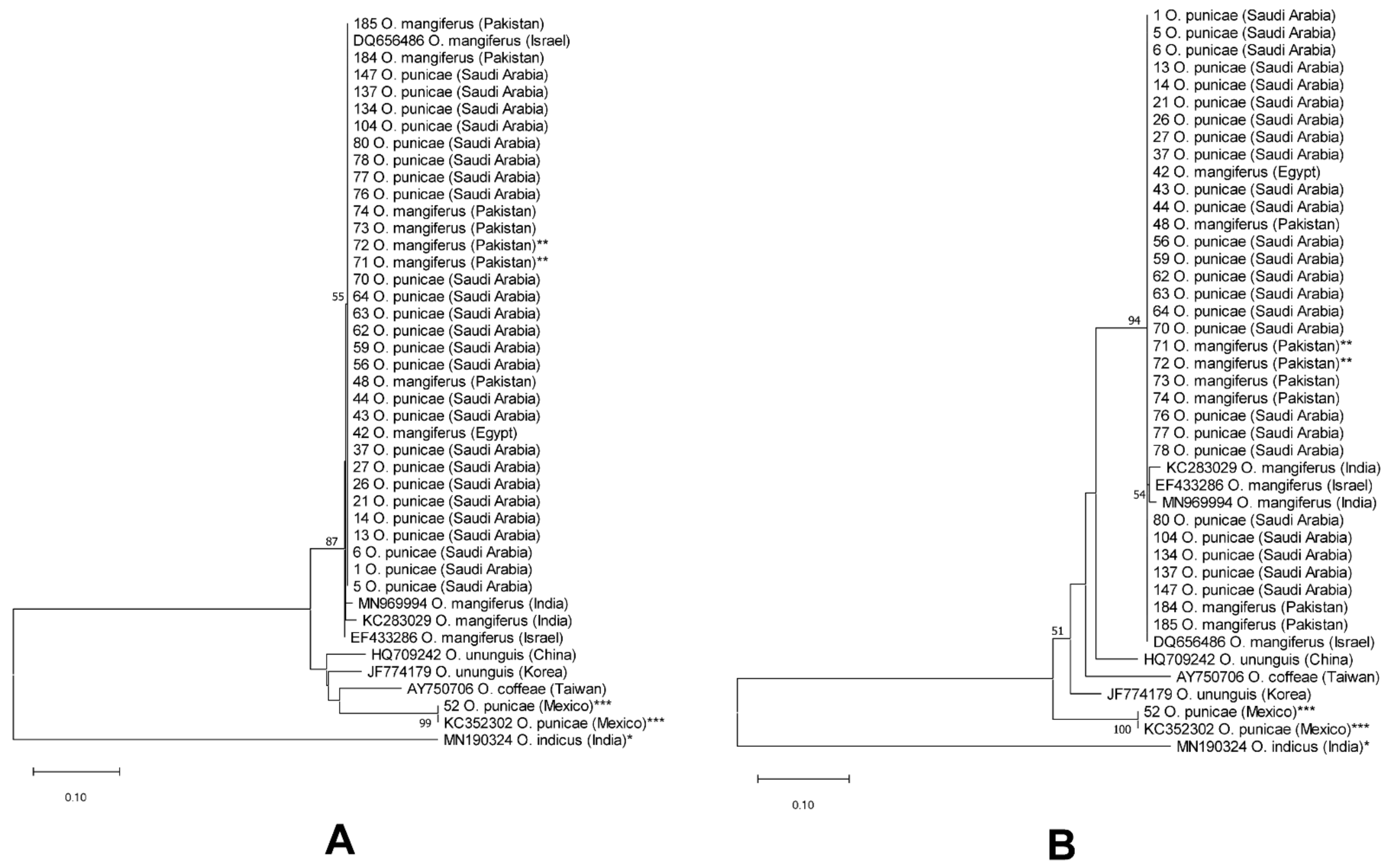
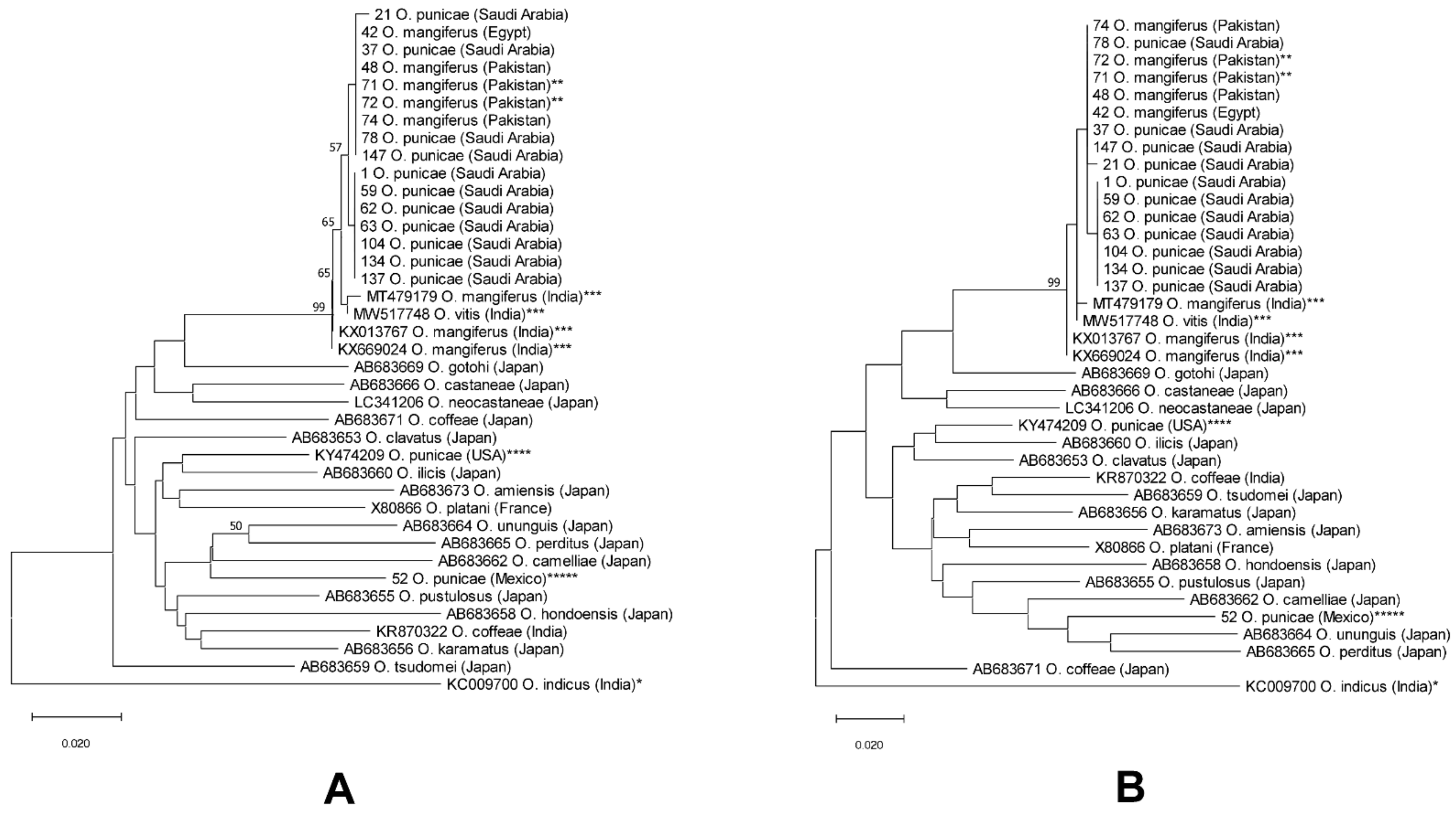
3.2. Molecular Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mushtaq, H.M.S.; Alatawi, F.J.; Kamran, M.; Flechtmann, C.H.W. The genus Oligonychus Berlese (Acari, Prostigmata, Tetranychidae): Taxonomic assessment and a key to subgenera, species groups, and subgroups. ZooKeys 2021, 1079, 89–127. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, H.M.S.; Kamran, M.; Alatawi, F.J. New species, new records, and re-descriptions of two species of the genus Oligonychus Berlese (Acari: Prostigmata: Tetranychidae) from Saudi Arabia. Syst. Appl. Acarol. 2022, 27, 2568–2582. [Google Scholar]
- Meyer, M.K.P.S. African Tetranychidae (Acari: Prostigmata)—With Reference to the World Genera, Entomology Memoir; Department of Agriculture and Water Supply: Pretoria, South Africa, 1987; pp. 1–175. [Google Scholar]
- Pritchard, A.E.; Baker, E.W. A Revision of the Spider Mite Family Tetranychidae; Pacific Coast Entomological Society: San Francisco, CA, USA, 1955; pp. 1–472. [Google Scholar] [CrossRef]
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants. University of California Press: Berkeley, CA, USA; pp. 1–614.
- Li, J.; Yi, T.C.; Guo, J.J.; Jin, D.C. Ontogenetic development and redescription of Oligonychus pratensis (Banks, 1912) (Acari: Tetranychidae). Zootaxa 2018, 4486, 349–375. [Google Scholar] [CrossRef]
- Meyer, M.K.P.S. A Revision of the Tetranychidae of Africa (Acari) with a Key to the Genera of the World, Entomology Memoir; Department of Agricultural Technical Services: Pretoria, South Africa, 1974; pp. 1–291. [Google Scholar]
- Mitrofanov, V.I.; Zapletina, A. A new species of the genus Oligonychus from Azerbaijan. Dokl. Akad. Nauk SSSR 1973, 29, 50–52. [Google Scholar]
- Rahman, K.A.; Sapra, A.N. Mites of the family Tetranychidae from Lyallpur with descriptions of four new species. P. Indian Acad. Sci. 1940, 11, 17–196. [Google Scholar] [CrossRef]
- Khanjani, M.; Khanjani, M.; Seeman, O. The spider mites of the genus Oligonychus Berlese (Acari: Tetranychidae) from Iran. Syst. Appl. Acarol. 2018, 23, 223–287. [Google Scholar] [CrossRef]
- Gupta, S.K. Contribution to our knowledge of tetranychid mites (Acarina) with descriptions of three new species from India. Orient. Insects 1976, 10, 327–351. [Google Scholar]
- Gupta, S.K. Plant Mites of India, Handbook; Zoological Survey of India: Calcutta, India, 1985; pp. 1–520. [Google Scholar]
- Gupta, S.K.; Gupta, Y.N. A Taxonomic Review of Indian Tetranychidae (Acari: Prostigmata) with Description of New Species, Redescriptions of Known Species and Keys to Genera and Species; Zoological Survey of India: Calcutta, India, 1994; pp. 1–196. [Google Scholar]
- McGregor, E.A. Mites of the family Tetranychidae. Am. Midl. Nat. 1950, 44, 257–420. [Google Scholar] [CrossRef]
- Nassar, O.; Ghai, S. Taxonomic studies on tetranychoid mites infesting vegetable and fruit crops in Delhi and surrounding areas. Orient. Insects 1981, 15, 333–396. [Google Scholar] [CrossRef]
- Zaher, M.A.; Gomaa, E.A.; El-Enany, M.A. Spider mites of Egypt (Acari: Tetranychidae). Int. J. Acarol. 1982, 8, 91–114. [Google Scholar] [CrossRef]
- Zeity, M. Tetranychid Mite Fauna of Major Agroecosystems in Karnatka and Some Aspects of Molecular Characterization of Selected Genera of Spider Mites. Ph.D. Thesis, University of Agricultural Sciences, Bangalore, India, 2015. [Google Scholar]
- Hirst, S. Descriptions of new mites including four new species of “red spider”. Proc. Zool. Soc. Lond. 1926, 96, 825–841. [Google Scholar] [CrossRef]
- Zaher, M.A.; Shehata, K.K. Oligonychus vitis n. sp. (Acarina: Tetranychidae). Bull. Inst. Egypte 1965, 49, 67–69. [Google Scholar]
- Spider Mites Web. Available online: http://www1.montpellier.inra.fr/CBGP/spmweb (accessed on 20 October 2022).
- Cromroy, H.L. A preliminary survey of the plant mites of Puerto Rico. J. Agric. Univ. P. R. 1958, 42, 39–144. [Google Scholar]
- Ehara, S.; Gotoh, T. Two new species of Oligonychus closely related to Oligonychus gotohi Ehara (Acari: Tetranychidae). Int. J. Acarol. 2007, 33, 15–20. [Google Scholar] [CrossRef]
- Lara, J.R.; Rugman-Jones, P.F.; Stouthamer, R.; Hoddle, M.S. Population genetics of Oligonychus perseae (Acari: Tetranychidae) collected from avocados in Mexico and California. Fla. Entomol. 2017, 100, 616–626. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yi, T.-C.; Guo, J.-J.; Jin, D.-C. Redescription of three species of Oligonychus (Acari, Tetranychidae) from China. Syst. Appl. Acarol. 2019, 24, 1071–1106. [Google Scholar] [CrossRef]
- Lo, P.K.C.; Ho, C.C. The spider mite family Tetranychidae in Taiwan I. The genus Oligonychus. J. Taiwan Mus. 1989, 42, 59–76. [Google Scholar]
- Arabuli, T.; Gotoh, T. A new species of spider mite, Oligonychus neocastaneae sp. nov. (Acari: Tetranychidae), from Japan. Zootaxa 2018, 4378, 563–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotoh, T.; Abe, S.; Kitashima, Y.; Ehara, S. Divergence in host range and reproductive compatibility in three strains of Oligonychus gotohi Ehara (Acari: Tetranychidae). Int. J. Acarol. 2007, 33, 7–13. [Google Scholar] [CrossRef]
- Matsuda, T.; Hinomoto, N.; Singh, R.N.; Gotoh, T. Molecular-based identification and phylogeny of Oligonychus species (Acari: Tetranychidae). J. Econ. Entomol. 2012, 105, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Ben-David, T.; Melamed, S.; Gerson, U.; Morin, S. ITS2 sequences as barcodes for identifying and analyzing spider mites (Acari: Tetranychidae). Exp. Appl. Acarol. 2007, 41, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, T.; Fukumoto, C.; Hinomoto, N.; Gotoh, T. DNA-Based Identification of Spider Mites: Molecular Evidence for Cryptic Species of the genus Tetranychus (Acari: Tetranychidae). J. Econ. Entomol. 2013, 106, 463–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirza, J.H.; Kamran, M.; Saleh, A.A.; Alatawi, F.J. Molecular and phenotypic variations in Eutetranychus orientalis (Klein) populations from Saudi Arabia. PLoS ONE 2020, 15, e0233389. [Google Scholar] [CrossRef] [PubMed]
- Navajas, M.; Gutierrez, J.; Lagnel, J.; Boursot, P. Mitochondrial cytochrome oxidase I in tetranychid mites: A comparison between molecular phylogeny and changes of morphological and life history traits. Bull. Entomol. Res. 1996, 86, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Gotoh, T.; Araki, R.; Boubou, A.; Migeon, A.; Ferragut, F.; Navajas, M. Evidence of co-specificity between Tetranychus evansi and Tetranychus takafujii (Acari: Prostigmata, Tetranychidae): Comments on taxonomic and agricultural aspects. Int. J. Acarol. 2009, 35, 485–501. [Google Scholar] [CrossRef]
- Navajas, M.; Gutierrez, J.; Williams, M.; Gotoh, T. Synonymy between two spider mite species, Tetranychus kanzawai and T. hydrangeae (Acari: Tetranychidae), shown by ribosomal ITS2 sequences and cross-breeding experiments. Bull. Entomol. Res. 2001, 91, 117–123. [Google Scholar] [CrossRef]
- Zeity, M.; Srinivas, N.; Gowda, C.C. Are Tetranychus macfarlanei Baker & Pritchard and Tetranychus malaysiensis Ehara (Acari: Tetranychidae) one species? Morphological and molecular evidences for synonymy between these two spider mite species and a note on invasiveness of T. macfarlanei on okra and eggplant in India. Syst. Appl. Acarol. 2017, 22, 467. [Google Scholar] [CrossRef]
- Alatawi, F.J.; Kamran, M. Spider mites (Acari: Tetranychidae) of Saudi Arabia: Two new species, new records and a key to all known species. J. Nat. Hist. 2018, 52, 429–455. [Google Scholar] [CrossRef]
- Guzman-Valencia, S.; Santillán-Galicia, M.; Guzmán-Franco, A.; González-Hernández, H.; Carrillo-Benítez, M.; Suárez-Espinoza, J. Contrasting effects of geographical separation on the genetic population structure of sympatric species of mites in avocado orchards. Bull. Entomol. Res. 2014, 104, 610–621. [Google Scholar] [CrossRef]
- Marei, F.A.; Negm, M.W.; Nasser, M.A.; Eraky, S.A. Population dynamics of Oligonychus mangiferus and Aceria mangiferae (Acari: Tetranychidae, Eriophyidae) on two mango cultivars in Assiut governorate, with an annotated checklist of mango mites in Egypt. Int. J. Entomol. Nematol. 2020, 6, 149–155. [Google Scholar]
- Bolland, H.R.; Gutierrez, J.; Flechtmann, C.H.W. World Catalogue of the Spider Mite Family (Acari: Tetranychidae); Brill Academic Publishers: Leiden, The Netherlands, 1998; pp. 1–392. [Google Scholar]
- Baker, E.W.; Tuttle, D.M. A Guide to the Spider Mites (Tetranychidae) of the United States; Indira Publishing House: West Bloomfield, MI, USA, 1994; pp. 1–347. [Google Scholar]
- Ben-David, T.; Ueckermann, E.; Gerson, U. An annotated list of the spider mites (Acari: Prostigmata: Tetranychidae) of Israel. Isr. J. Entomol. 2013, 43, 125–148. [Google Scholar]
- Tseng, Y.H. A Monograph of the Mite Family Tetranychidae (Acarina: Trombidiformes) from Taiwan; Taiwan Museum Special Publication Series: Taiwan, China, 1990; pp. 1–226. [Google Scholar]
- Tuttle, D.M.; Baker, E.W.; Abbatiello, M. Spider mites of Mexico (Acarina: Tetranychidae). Int. J. Acarol. 1976, 2, 1–102. [Google Scholar] [CrossRef]
- Mushtaq, H.M.S.; Kamran, M.; Alatawi, F.J. Keys to World Species of the Genus Oligonychus Berlese (Acari, Prostigmata, Tetranychidae): Taxonomic Notes and New Oligonychus Species; King Saud University: Riyadh, Saudi Arabia, 2022; manuscript in preparation. [Google Scholar]
- Baker, E.W.; Pritchard, A.E. Arañas rojas de América Central (Acarina: Tetranychidae). Rev. Soc. Mex. Hist. Nat. 1962, 23, 309–340. [Google Scholar]
- Spider Mites of Australia (Including Key Exotic Southeast Asian Pest Species). Available online: https://keys.lucidcentral.org/keys/v3/spider_mites_australia/key/spider_mites_of_australia/Media/Html/entities/Oligonychus_punicae_Hirst_1926.htm (accessed on 10 July 2022).
- Hall, T.A. BIOEDIT: A user-friendly biological sequence alignment editor and analysis program for WINDOWS 95/98/NT. Nucl. Acid. S. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [Green Version]
- Felsen-stein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar]
- Baker, E.W.; Pritchard, A.E. The tetranychoid mites of Africa. Hilgardia 1960, 29, 455–574. [Google Scholar] [CrossRef] [Green Version]
- Ben-David, T. Molecular Characterization of Israel’s Spider Mites (Acari: Tetranychidae). Ph.D. Thesis, The Hebrew University of Jerusalem, Jerusalem, Israel, 2008. [Google Scholar]
- Estebanes-Gonzalez, M.L.; Baker, E.W. Arañas rojas de Mexico (Acarina: Tetranychidae). An. Esc. Nac. Cien. Biol. 1968, 15, 61–133. [Google Scholar]
- Navajas, M.; Gutierrez, J.; Bonato, O.; Bolland, H.R.; Mapangou-Divassa, S. Intraspecific diversity of the cassava green mite Mononychellus progresivus (Acari: Tetranychidae) using comparisons of mitochondrial and nuclear ribosomal DNA sequences and cross-breeding. Exp. Appl. Acarol. 1994, 18, 351–360. [Google Scholar] [CrossRef]
- Sivakumar, K.; Kunchithpatham, R. Phytophagous mite diversity of Tetranychus spp. and Oligonychus spp. (Acari: Tetranychidae) found on different host plants in Coimbatore district and surrounding regions. BioSci. Trends 2014, 7, 4113–4117. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mushtaq, H.M.S.; Kamran, M.; Saleh, A.A.; Alatawi, F.J. Evidence for Reconsidering the Taxonomic Status of Closely Related Oligonychus Species in punicae Complex (Acari: Prostigmata: Tetranychidae). Insects 2023, 14, 3. https://doi.org/10.3390/insects14010003
Mushtaq HMS, Kamran M, Saleh AA, Alatawi FJ. Evidence for Reconsidering the Taxonomic Status of Closely Related Oligonychus Species in punicae Complex (Acari: Prostigmata: Tetranychidae). Insects. 2023; 14(1):3. https://doi.org/10.3390/insects14010003
Chicago/Turabian StyleMushtaq, Hafiz Muhammad Saqib, Muhammad Kamran, Amgad A. Saleh, and Fahad Jaber Alatawi. 2023. "Evidence for Reconsidering the Taxonomic Status of Closely Related Oligonychus Species in punicae Complex (Acari: Prostigmata: Tetranychidae)" Insects 14, no. 1: 3. https://doi.org/10.3390/insects14010003
APA StyleMushtaq, H. M. S., Kamran, M., Saleh, A. A., & Alatawi, F. J. (2023). Evidence for Reconsidering the Taxonomic Status of Closely Related Oligonychus Species in punicae Complex (Acari: Prostigmata: Tetranychidae). Insects, 14(1), 3. https://doi.org/10.3390/insects14010003