
Warming in Cold Seasons Increases the Abundance of Ground-Dwelling Collembola in Permafrost Wetlands


 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
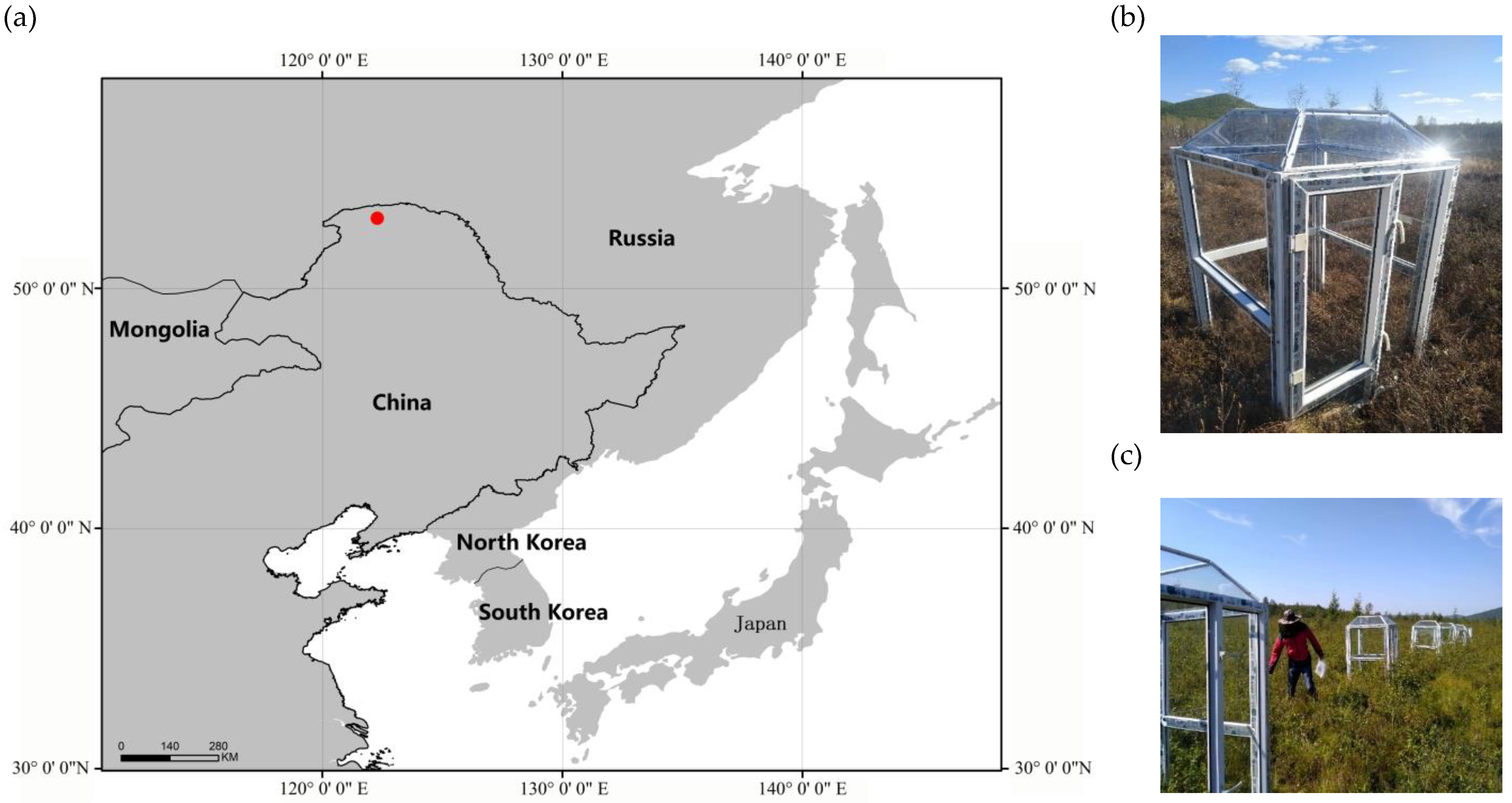
2.1. Study Locality, Plots, and Experimental Design
2.2. Sampling and Environmental Viariables
2.3. Data Analyses
3. Results
3.1. Environmental, Soil and Plant Properties
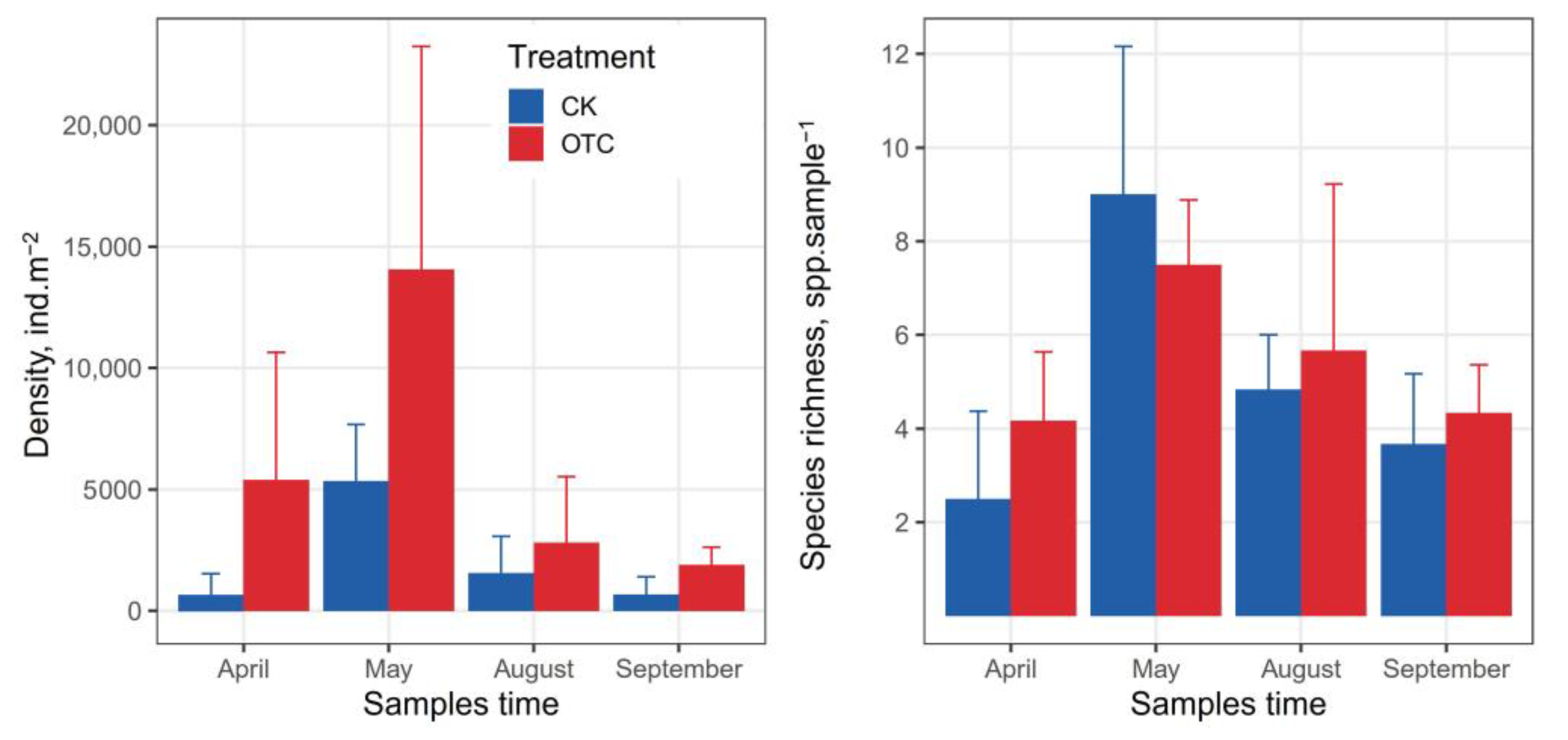
3.2. Abundance and Species Richness of Collembola
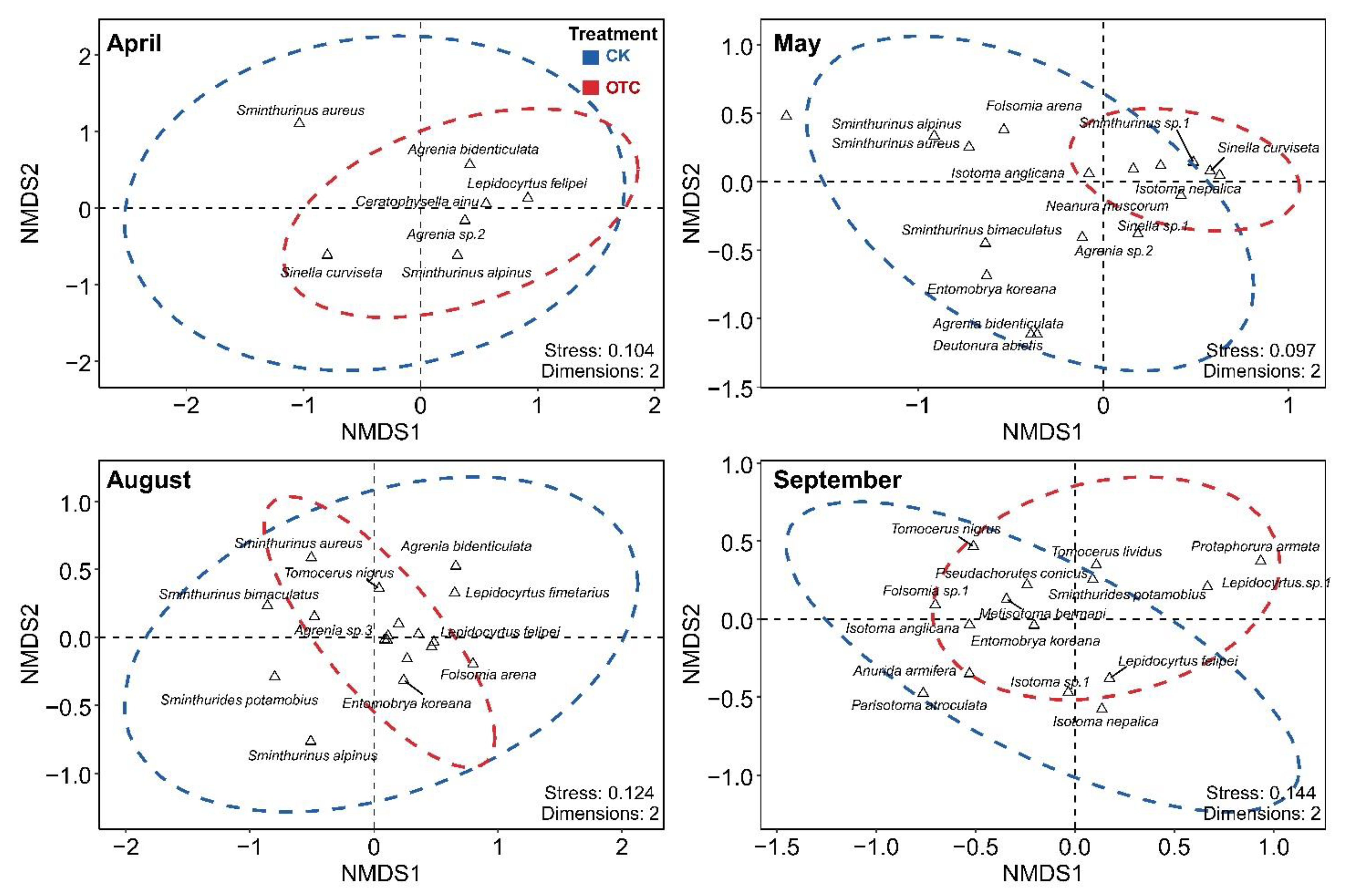
3.3. Collembola Community Composition
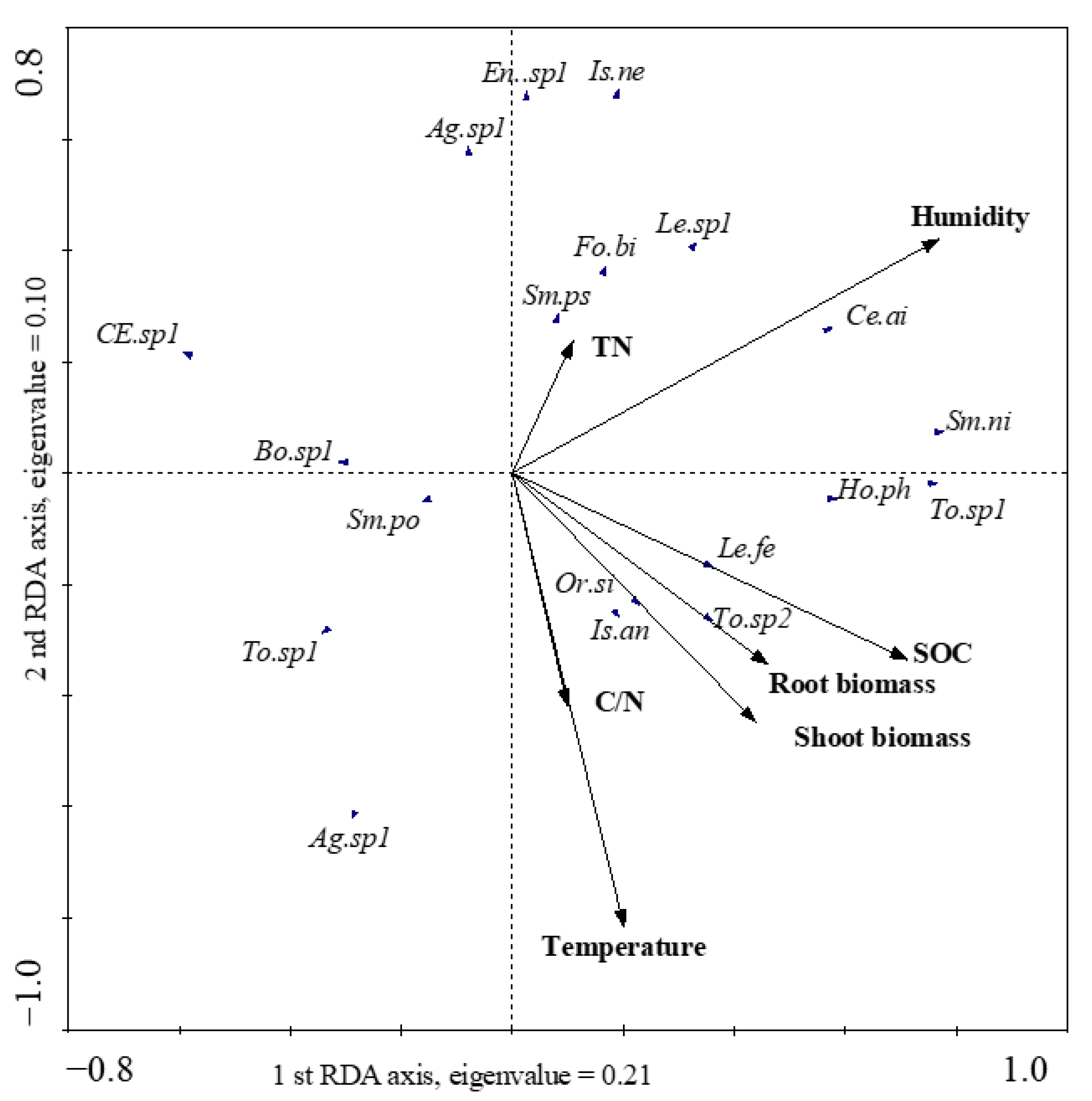
3.4. Relationships between Collembola Communities and Environmental Factors
4. Discussion
4.1. The Effect of Warming on Abundance and Species Richness of Collembola
4.2. The Effect of Warming and Sampling Time on Community Composition
4.3. Relationship with Environmental Factors
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agrenia bidenticulata (Tullberg, 1876) (Ag.bi) | Neanura muscorum (Templeton, 1836) (Ne.mu) |
| Agrenia sp.1 (Ag.sp1) | Parisotoma atroculata (Potapov, 1991) (Pr.at) |
| Agrenia sp.2 (Ag.sp2) | Protaphorura armata (Tullberg, 1869) (Pr.ar) |
| Anurida armifera (Babenko, Shveenkova & Kuznetsova, 2019) (An.ar) | Pseudachorutes conicus (Lee and Kim, 1984) (Ps.co) |
| Ceratophysella ainu (Yosii, 1972) (Ce.ai) | Sinella curviseta (Brook, 1882) (Si.cu) |
| Deutonura abietis (Yosii, 1969) (De.ab) | Sinella sp.1 (Si.sp1) |
| Entomobrya koreana (Yosii, 1965) (En.ko) | Sminthurides potamobius (Yosii, 1970) (Sm.po) |
| Folsomia arena (Potapov and Babenko, 2006) (Fo.ar) | Sminthurides pseudassimillis (Stach, 1956) (Sm.ps) |
| Fosolmia bidendata (Lee, 1974) (Fo.bi) | Sminthurinus alpinus (Gisin, 1953) (Sm.al) |
| Isotoma anglicana (Lubbock, 1862) (Is.an) | Sminthurinus aureus (Lubbock, J, 1862) (Sm.au) |
| Isotoma nepalica (Yosii, 1966) (Is.ne) | Sminthurinus bimaculatus (Axelson, 1902) (Sm.bi) |
| Isotoma sp.1 (Is.sp1) | Sminthurinus sp.1 (Sm.sp1) |
| Lepidocyrtus felipei (Wang, Chen and Christiansen, 2003) (Le.fe) | Tomocerus nigrus (Sun, Liang and Huang, 2006) (To.ni) |
| Lepidocyrtus fimetarius (Gisin, 1964) (Le.fi) | Tomocerus lividus (Tullberg, 1877) (To.li) |
| Lepidocyrtus sp.1 (Le.sp1) | Tomocerus minor (Lubbock, 1862) (To.mi) |
| Metisotoma bermani (Potapov, Babenko, Fjellberg, and Schulz, 2018) (Me.be) |
References
- Sun, J.; Li, X.Z.; Wang, X.W.; Lv, J.J.; Hu, Y.M. Latitudinal pattern in species diversity and its response to global warming in permafrost wetlands in the Great Hing’an Mountains, China. Russ. J. Ecol. 2011, 42, 123–132. [Google Scholar] [CrossRef]
- Song, Y.Y.; Song, C.C.; Shi, F.X.; Wang, M.Q.; Ren, J.S.; Wang, X.W.; Jiang, L. Linking plant community composition with the soil C pool, N availability and enzyme activity in boreal peatlands of Northeast China. Appl. Soil Ecol. 2019, 140, 144–154. [Google Scholar] [CrossRef]
- Jiang, L.; Song, Y.; Sun, L.; Song, C.; Wang, X.; Ma, X.; Liu, C.; Gao, J. Effects of warming on carbon emission and microbial abundances across different soil depths of a peatland in the permafrost region under anaerobic condition. Appl. Soil Ecol. 2020, 156, 103712. [Google Scholar] [CrossRef]
- Song, Y.Y.; Liu, C.; Wang, X.W.; Ma, X.Y.; Jiang, L.; Zhu, J.P.; Gao, J.L.; Song, C.C. Microbial abundance as an indicator of soil carbon and nitrogen nutrient in permafrost peatlands. Ecol. Indic. 2020, 115, 106362. [Google Scholar] [CrossRef]
- Rustad, L.E.; Campbell, J.L.; Marion, G.M.; Norby, R.J.; Mitchell, M.J.; Hartley, A.E.; Cornelissen, J.H.C.; Gurevitch, J.; Gcet-News. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 2001, 126, 543–562. [Google Scholar] [CrossRef]
- Birkemoe, T.; Leinaas, H.P. Effects of temperature on the development of an arctic Collembola (Hypogastrura tullbergi). Funct. Ecol. 2000, 14, 693–700. [Google Scholar] [CrossRef]
- Huhta, V.; Hänninen, S.M. Effects of temperature and moisture fluctuations on an experimental soil microarthropod community. Pedobiologia 2001, 45, 279–286. [Google Scholar] [CrossRef]
- Birkemoe, T.; Somme, L. Population dynamics of two collembolan species in an Arctic tundra. Pedobiologia 1998, 42, 131–145. [Google Scholar]
- Kardol, P.; Reynolds, W.N.; Norby, R.J.; Classen, A.T. Climate change effects on soil microarthropod abundance and community structure. Appl. Soil Ecol. 2011, 47, 37–44. [Google Scholar] [CrossRef]
- Convey, P.; Block, W.; Peat, H.J. Soil arthropods as indicators of water stress in Antarctic terrestrial habitats? Glob. Change Biol. 2003, 9, 1718–1730. [Google Scholar] [CrossRef]
- Convey, P.; Peck, L.S. Antarctic environmental change and biological responses. Sci. Adv. 2019, 5, eaaz0888. [Google Scholar] [CrossRef] [PubMed]
- Hertzberg, K.; Leinaas, H.P.; Ims, R.A. Patterns of abundance and demography: Collembola in a habitat patch gradient. Ecography 1994, 17, 349–359. [Google Scholar] [CrossRef]
- Krab, E.J.; Berg, M.P.; Aerts, R.; van Logtestijn, R.S.P.; Cornelissen, J.H.C. Vascular plant litter input in subarctic peat bogs changes Collembola diets and decomposition patterns. Soil Biol. Biochem. 2013, 63, 106–115. [Google Scholar] [CrossRef]
- Chang, L.; Wu, H.; Wu, D.; Sun, X. Effect of tillage and farming management on Collembola in marsh soils. Appl. Soil Ecol. 2013, 64, 112–117. [Google Scholar] [CrossRef]
- Day, T.A.; Ruhland, C.T.; Strauss, S.L.; Park, J.H.; Krieg, M.L.; Krna, M.A.; Bryant, D.M. Response of plants and the dominant microarthropod, Cryptopygus antarcticus, to warming and contrasting precipitation regimes in Antarctic tundra. Glob. Change Biol. 2010, 15, 1640–1651. [Google Scholar] [CrossRef]
- Jucevica, E.; Melecis, V. Global warming affect Collembola community: A long-term study. Pedobiologia 2006, 50, 177–184. [Google Scholar] [CrossRef]
- A’Bear, A.D.; Boddy, L.; Jones, T.H. Bottom-up determination of soil collembola diversity and population dynamics in response to interactive climatic factors. Oecologia 2013, 173, 1083–1087. [Google Scholar] [CrossRef] [PubMed]
- Blankinship, J.C.; Niklaus, P.A.; Hungate, B.A. A meta-analysis of responses of soil biota to global change. Oecologia 2011, 165, 553–565. [Google Scholar] [CrossRef]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Mundepi, A.; Phillips, C.A.; Callaham, A. Soil fauna responses to natural disturbances, invasive species, and global climate change: Current state of the science and a call to action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Filser, J. The role of Collembola in carbon and nitrogen cycling in soil. Pedobiologia 2002, 46, 234–245. [Google Scholar] [CrossRef]
- Miao, Y.; Song, C.; Sun, L.; Wang, X.; Meng, H.; Mao, R. Growing season methane emission from a boreal peatland in the continuous permafrost zone of Northeast China: Effects of active layer depth and vegetation. Biogeosciences 2012, 9, 4455–4464. [Google Scholar] [CrossRef]
- Siddiky, M.R.K.; Kohler, J.; Cosme, M.; Rillig, M.C. Soil biota effects on soil structure: Interactions between arbuscular mycorrhizal fungal mycelium and Collembola. Soil Biol. Biochem. 2012, 50, 33–39. [Google Scholar] [CrossRef]
- Zou, Y.; van Telgen, M.D.; Chen, J.; Xiao, H.; de Kraker, J.; Bianchi, F.J.; van der Werf, W. Modification and application of a leaf blower-vac for field sampling of arthropods. J. Vis. Exp. 2016, 114, e54655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiansen, K.; Bellinger, P. The Collembola of North America north of the Rio Grande. A taxonomic analysis. Deut. Entomol. Z. 1984, 31, 8. [Google Scholar]
- Hopkin, S.P. Biology of the Springtails (Insecta: Collembola); Oxford University Press: Oxford, UK, 1997; pp. 1–330. [Google Scholar]
- Pinheiro, J.; Bates, D.; R Core Team. NLME: Linear and Nonlinear Mixed Effects Models; European Environment Agency: Copenhagen, Denmark, 2022. [Google Scholar]
- Lenth, R.V. EMMEANS: Estimated Marginal Means, aka Least-Squares Means; European Environment Agency: Copenhagen, Denmark, 2021. [Google Scholar]
- Krab, E.J.; Van, S.L.I.M.; Cornelissen, J.H.C.; Berg, M.P. How extreme is an extreme climatic event to a subarctic peatland springtail community? Soil Biol. Biochem. 2013, 59, 16–24. [Google Scholar] [CrossRef]
- Batzer, D.P.; Wu, H. Ecology of Terrestrial Arthropods in Freshwater Wetlands. Annu. Rev. Entomol. 2020, 65, 101–119. [Google Scholar] [CrossRef] [Green Version]
- Juan-Ovejero, R.; Benito, E.; Barreal, M.E.; Rodeiro, J.; Briones, M.J.I. Tolerance to fluctuating water regimes drives changes in mesofauna community structure and vertical stratification in peatlands. Pedobiologia 2019, 76, 150571. [Google Scholar] [CrossRef]
- Laiho, R.; Silvan, N.; Cárcamo, H.; Vasander, H. Effects of water level and nutrients on spatial distribution of soil mesofauna in peatlands drained for forestry in Finland. Appl. Soil Ecol. 2001, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dou, Y.; Chang, L.; Zhang, B.; Wu, D. Effect of Soybean Cultivation on Soil Collembola Community in Marshland of Sanjiang Plain, China. Russ. J. Ecol. 2018, 49, 570–576. [Google Scholar] [CrossRef]
- Chang, L.; Sun, X.; Wang, B.; Gao, M.; Chen, L.; Liang, A.; Wu, D. Green more than brown food resources drive the effect of simulated climate change on Collembola: A soil transplantation experiment in Northeast China. Geoderma 2021, 392, 115008. [Google Scholar] [CrossRef]
- Thakur, M.P.; Künne, T.; Griffin, J.N.; Eisenhauer, N. Warming magnifies predation and reduces prey coexistence in a model litter arthropod system. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kustec, J. Top-Down and Bottom-Up Effects on Collembola Communities in Soil Food Webs. Master’s Thesis, The University of Western Ontario, London, ON, Canada, 2018. [Google Scholar]
- Coulson, S.J.; Hodkinson, I.D.; Wooley, C.; Webb, N.R.; Block, W.; Worland, M.R.; Bale, J.S.; Strathdee, A.T. Effects of experimental temperature elevation on high-arctic soil microarthropod populations. Polar Biol. 1996, 16, 147–153. [Google Scholar] [CrossRef]
- Makkonen, M.; Berg, M.P.; van Hal, J.R.; Callaghan, T.V.; Press, M.C.; Aerts, R. Traits explain the responses of a sub-arctic Collembola community to climate manipulation. Soil Biol. Biochem. 2011, 43, 377–384. [Google Scholar] [CrossRef]
- Dollery, R.; Hodkinson, I.D.; Jónsdóttir, I.S. Impact of warming and timing of snow melt on soil microarthropod assemblages associated with Dryas-dominated plant communities on Svalbard. Ecography 2006, 29, 111–119. [Google Scholar] [CrossRef]
- Hågvar, S.; Klanderud, K. Effect of simulated environmental change on alpine soil arthropods. Glob. Change Biol. 2009, 15, 2972–2980. [Google Scholar] [CrossRef]
- Sjursen, H.; Michelsen, A.; Jonasson, S. Effects of long-term soil warming and fertilisation on microarthropod abundances in three sub-arctic ecosystems. Appl. Soil Ecol. 2005, 30, 148–161. [Google Scholar] [CrossRef]
- Bokhorst, S.; Huiskes, A.; Convey, P.; van Bodegom, P.M.; Aerts, R. Climate change effects on soil arthropod communities from the Falkland Islands and the Maritime Antarctic. Soil Biol. Biochem. 2008, 40, 1547–1556. [Google Scholar] [CrossRef]
- Richardson, S.J.; Press, M.C.; Parsons, A.N.; Hartley, S.E. How do nutrients and warming impact on plant communities and their insect herbivores ? A 9-year study from a sub-Arctic heath. J. Ecol. 2002, 90, 544–556. [Google Scholar] [CrossRef]
- Bokhorst, S.; Huiskes, A.; Convey, P.; Aerts, R. The effect of environmental change on vascular plant and cryptogam communities from the Falkland Islands and the Maritime Antarctic. BMC Ecol. 2007, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, U.N.; Wall, D.H. The future of soil invertebrate communities in polar regions: Different climate change responses in the Arctic and Antarctic? Ecol. Lett. 2013, 16, 409–419. [Google Scholar] [CrossRef]
- Harte, J.; Rawa, A.; Price, V. Effects of manipulated soil microclimate on mesofaunal biomass and diversity. Soil Biol. Biochem. 1996, 28, 313–322. [Google Scholar] [CrossRef]
- John, C.M.; Kevin, M.; Heikki, S.; Peter, C.D.R. Top-down is bottom-up: Does predation in the rhizosphere regulate above ground. Ecology 2003, 84, 846–857. [Google Scholar]
- Scheu, S.; Schaefer, M. Bottom-up control of the soil macrofauna community in a beechwood on limestone: Manipulation of food resources. Ecology 1998, 79, 1573–1585. [Google Scholar] [CrossRef]
- Roos, R.E.; Birkemoe, T.; Asplund, J.; Ľuptáčik, P.; Raschmanová, N.; Alatalo, J.; Olsen, S.L.; Klanderud, K. Legacy effects of experimental environmental change on soil micro-arthropod communities. Ecosphere 2020, 11, e03030. [Google Scholar] [CrossRef]




| Samples | Shoot Biomass | Root Biomass | Soil Nitrogen | Soil Organic Carbon | C/N | Air Humidity (%) | Air Temperature (℃) |
|---|---|---|---|---|---|---|---|
| CK | 752.7 ± 58.03 a | 70.37 ± 21.22 b | 6.27 ± 1.72 a | 487.77 ± 29.93 a | 85.08 ± 28.98 a | 13.02 ± 3.48 a | 18.6 ± 0.36 b |
| OTC | 965.47 ± 425.8 a | 144.17 ± 39.85 a | 5.2 ± 2.2 a | 537.27 ± 73.87 a | 112.77 ± 34.78 a | 17.29 ± 6.71 a | 19.8 ± 0.35 a |
| Variables | Abundance of Collembola | Richness of Collembola | ||||
|---|---|---|---|---|---|---|
| Df | F-Value | p | Df | F-Value | p | |
| Warming | 1.10 | 18.6619 | 0.002 | 1.10 | 1.6323 | 0.23 |
| Time | 3.30 | 11.6548 | <0.001 | 3.30 | 13.5216 | <0.001 |
| Warming × Time | 3.30 | 2.0633 | 0.126 | 3.30 | 2.105 | 0.121 |
| Environmental Variables | % | F | p |
|---|---|---|---|
| Humidity | 15 | 1.78 | 0.084 |
| Temperature | 13 | 1.62 | 0.116 |
| Root biomass | 8 | 0.94 | 0.508 |
| Shoot biomass | 3 | 0.45 | 0.880 |
| Total nitrogen | 4 | 0.42 | 0.870 |
| Total % variance explained | 43 | —— | —— |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Xie, Z.; Dou, Y.; Sun, X.; Chang, L.; Wu, D. Warming in Cold Seasons Increases the Abundance of Ground-Dwelling Collembola in Permafrost Wetlands. Insects 2023, 14, 33. https://doi.org/10.3390/insects14010033
Zhang S, Xie Z, Dou Y, Sun X, Chang L, Wu D. Warming in Cold Seasons Increases the Abundance of Ground-Dwelling Collembola in Permafrost Wetlands. Insects. 2023; 14(1):33. https://doi.org/10.3390/insects14010033
Chicago/Turabian StyleZhang, Shaoqing, Zhijing Xie, Yongjing Dou, Xin Sun, Liang Chang, and Donghui Wu. 2023. "Warming in Cold Seasons Increases the Abundance of Ground-Dwelling Collembola in Permafrost Wetlands" Insects 14, no. 1: 33. https://doi.org/10.3390/insects14010033
APA StyleZhang, S., Xie, Z., Dou, Y., Sun, X., Chang, L., & Wu, D. (2023). Warming in Cold Seasons Increases the Abundance of Ground-Dwelling Collembola in Permafrost Wetlands. Insects, 14(1), 33. https://doi.org/10.3390/insects14010033