

Group-Living Spider Cyrtophora citricola as a Potential Novel Biological Control Agent of the Tomato Pest Tuta absoluta


{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collecting and Rearing C. citricola Spiderlings Prior to Experimental Setup
2.2. Rearing T. absoluta Moths and Flies
2.3. Experimental Setup of Prey Capture Assays
2.4. Prey Capture Assays
2.5. Seasonal Web Size Measurement and Effects of Egg Predators
2.6. Statistics
2.6.1. Spider Sizes
2.6.2. Control Prey Capture
2.6.3. Prey Capture Treatments
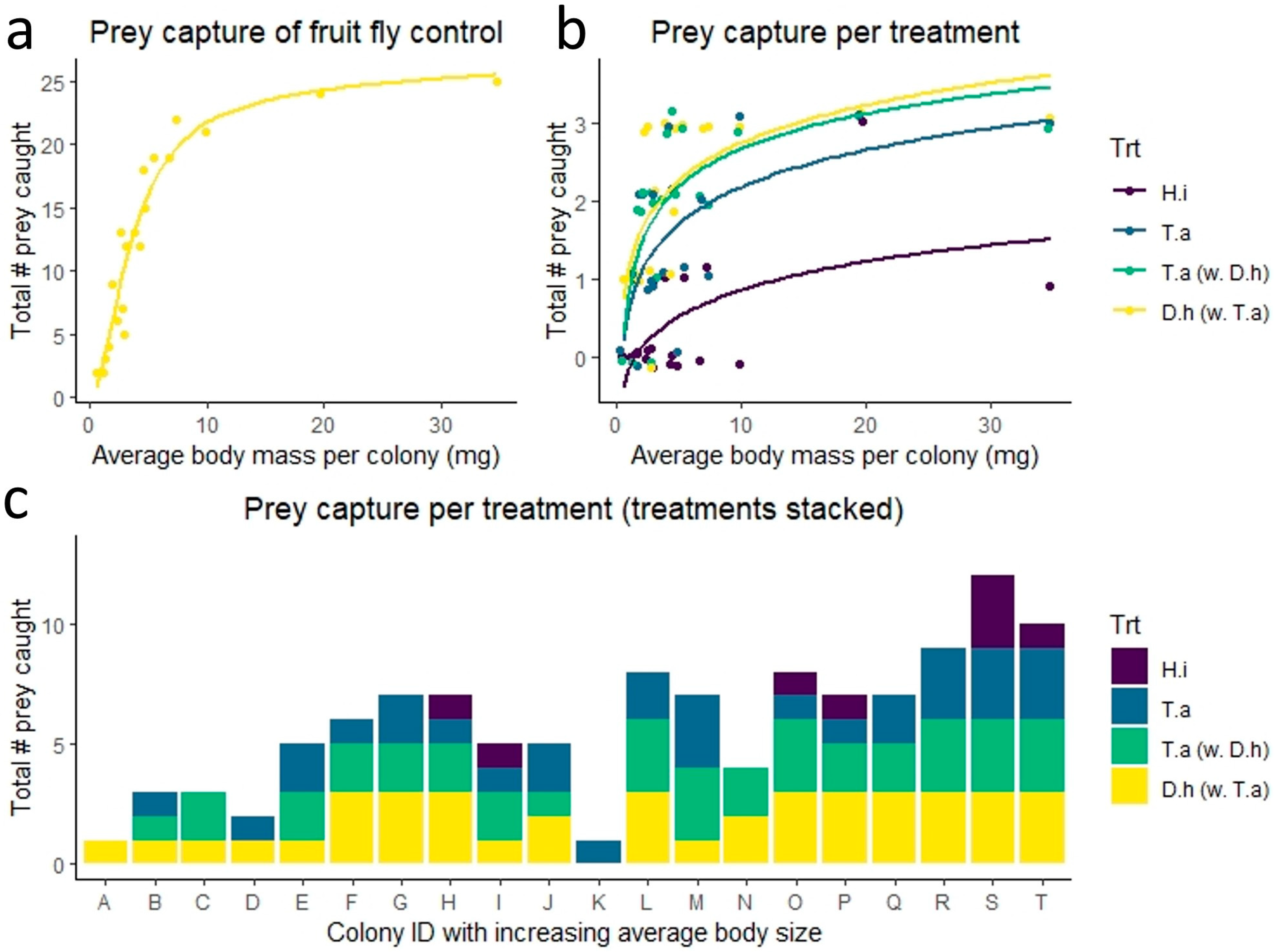
3. Results
3.1. Spider Sizes
3.2. Control Prey Capture
3.3. Prey Capture Treatments
3.4. Seasonality
3.5. Wasp Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Campos, M.R.; Biondi, A.; Adiga, A.; Guedes, R.N.C.; Desneux, N. From the Western Palaearctic region to beyond: Tuta absoluta 10 years after invading Europe. J. Pest Sci. 2017, 90, 787–796. [Google Scholar] [CrossRef]
- Arnó, J.; Molina, P.; Aparicio, Y.; Denis, C.; Gabarra, R.; Riudavets, J. Natural enemies associated with Tuta absoluta and functional biodiversity in vegetable crops. BioControl 2021, 66, 613–623. [Google Scholar] [CrossRef]
- Tarusikirwa, V.L.; Machekano, H.; Mutamiswa, R.; Chidawanyika, F.; Nyamukondiwa, C. Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) on the “offensive” in Africa: Prospects for integrated pest management initiatives. Insects 2020, 11, 764. [Google Scholar] [CrossRef]
- Gebremariam, G. Tuta absoluta: A global looming challenge in tomato production. J. Biol. Agric. Healthc. 2015, 5, 57–63. [Google Scholar]
- Tropea Garzia, G.; Siscaro, G.; Biondi, A.; Zappalà, L. Tuta absoluta, a South American pest of tomato now in the EPPO region: Biology, distribution and damage. EPPO Bull. 2012, 42, 205–210. [Google Scholar] [CrossRef]
- Michalko, R.; Pekár, S.; Entling, M.H. An updated perspective on spiders as generalist predators in biological control. Oecologia 2019, 189, 21–36. [Google Scholar] [CrossRef]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Ruescas, D.C.; Tabone, E.; Frandon, J.; et al. Biological invasion of European tomato crops by Tuta absoluta: Ecology, geographic expansion and prospects for biological control. J. Pest Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Chakraborty, S.; Newton, A.C. Climate change, plant diseases and food security: An overview. Plant Pathol. 2011, 60, 2–14. [Google Scholar] [CrossRef]
- Ferracini, C.; Bueno, V.H.P.; Dindo, M.L.; Ingegno, B.L.; Luna, M.G.; Salas Gervassio, N.G.; Sánchez, N.E.; Siscaro, G.; van Lenteren, J.C.; Zappalà, L.; et al. Natural enemies of Tuta absoluta in the Mediterranean basin, Europe and South America. Biocontrol Sci. Technol. 2019, 29, 578–609. [Google Scholar] [CrossRef]
- Van Den Bosch, R. Biological control of insects. Annu. Rev. Ecol. Syst. 1971, 2, 45–66. [Google Scholar] [CrossRef]
- Buragohain, P.; Saikia, D.K.; Sotelo-Cardona, P.; Srinivasan, R. Evaluation of bio-pesticides against the south american tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) in India. Horticulturae 2021, 7, 325. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Picanço, M.C. The tomato borer Tuta absoluta in South America: Pest status, management and insecticide resistance. EPPO Bull. 2012, 42, 211–216. [Google Scholar] [CrossRef]
- Ecobichon, D.J. Pesticide use in developing countries. Toxicology 2001, 160, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Terziev, V.; Petkova-Georgieva, S. The pesticides toxic impact on the human health condition and the ecosystem. SSRN Electron. J. 2019, 5, 1314–1320. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Matviishyn, T.M.; Husak, V.V.; Storey, J.M.; Storey, K.B. Pesticide toxicity: A mechanistic approach. EXCLI J. 2018, 17, 1101–1136. [Google Scholar] [CrossRef]
- Dlamini, B.E.; Dlamini, N.; Masarirambi, M.T.; Nxumalo Kwanele, A. Control of the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) larvae in laboratory using entomopathogenic nematodes from subtropical environment. J. Nematol. 2020, 52, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Yanira Olivera, S.; Arroyo, N.; Asencio, C. Cyrtophora citricola as a potential component of an integrated pest management program for citrus in Puerto Rico. FASEB J. 2013, 27 (Suppl. 1). [Google Scholar] [CrossRef]
- Desneux, N.; Han, P.; Mansour, R.; Arnó, J.; Brévault, T.; Campos, M.R. Integrated pest management of Tuta absoluta: Practical implementations across different world regions. J. Pest Sci. 2022, 95, 17–39. [Google Scholar] [CrossRef]
- Gonzalez-Cabrera, J.; Molla, O.; Monton, H.; Urbaneja, A. Efficacy of Bacillus thuringiensis (Berliner) in controlling the tomato borer, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). BioControl 2011, 56, 71–80. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Alves, S.B.; Rampelotti-Ferreira, F.T.; Urbano, M.R.; Demétrio, C.G.B.; Delalibera, I. Potential of a granulovirus isolate to control Phthorimaea operculella (Lepidoptera: Gelechiidae). BioControl 2010, 55, 657–671. [Google Scholar] [CrossRef]
- Aigbedion-Atalor, P.O.; Mohamed, S.A.; Hill, M.P.; Zalucki, M.P.; Azrag, A.G.A.; Srinivasan, R.; Ekesi, S. Host stage preference and performance of Dolichogenidea gelechiidivoris (Hymenoptera: Braconidae), a candidate for classical biological control of Tuta absoluta in Africa. Biol. Control 2020, 144, 104215. [Google Scholar] [CrossRef]
- Contreras, J.; Mendoza, J.E.; Martínez-Aguirre, M.R.; García-Vidal, L.; Izquierdo, J.; Bielza, P. Efficacy of enthomopathogenic fungus Metarhizium anisopliae against Tuta absoluta (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2014, 107, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Kamali, S.; Karimi, J.; Koppenhöfer, A.M. New insight into the management of the tomato leaf miner, Tuta absoluta (Lepidoptera: Gelechiidae) with entomopathogenic nematodes. J. Econ. Entomol. 2018, 111, 112–119. [Google Scholar] [CrossRef]
- Chuang, A.; Gates, M.W.; Grinsted, L.; Askew, R.; Leppanen, C. Two hymenopteran egg sac associates of the tent-web orbweaving spider, Cyrtophora citricola (Forskål, 1775) (Araneae, Araneidae). ZooKeys 2019, 874, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Lubin, Y.; Bilde, T. The evolution of sociality in spiders. Adv. Study Behav. 2007, 37, 83–145. [Google Scholar] [CrossRef]
- World Spider Catalog. Available online: https://wsc.nmbe.ch/species/3773 (accessed on 17 October 2022).
- Urbaneja, A.; González-Cabrera, J.; Arnó, J.; Gabarra, R. Prospects for the biological control of Tuta absoluta in tomatoes of the Mediterranean basin. Pest Manag. Sci. 2012, 68, 1215–1222. [Google Scholar] [CrossRef]
- Rypstra, A.L. Foraging flocks of spiders. Behav. Ecol. Sociobiol. 1979, 5, 291–300. [Google Scholar] [CrossRef]
- Yip, E.C.; Rao, D.; Smith, D.R.; Lubin, Y. Interacting maternal and spatial cues influence natal—Dispersal out of social groups. Oikos 2019, 128, 1793–1804. [Google Scholar] [CrossRef]
- Yip, E.C.; Levy, T.; Lubin, Y. Bad neighbors: Hunger and dominance drive spacing and position in an orb-weaving spider colony. Behav. Ecol. Sociobiol. 2017, 71, 128. [Google Scholar] [CrossRef]
- Johannesen, J.; Wennmann, J.T.; Lubin, Y. Dispersal behaviour and colony structure in a colonial spider. Behav. Ecol. Sociobiol. 2012, 66, 1387–1398. [Google Scholar] [CrossRef]
- Leborgne, R.; Cantarella, T.; Pasquet, A. Colonial life versus solitary life in Cyrtophora citricola (Araneae, Araneidae). Insectes Soc. 1998, 45, 125–134. [Google Scholar] [CrossRef]
- Yip, E.C.; Smith, D.R.; Lubin, Y. Long-term colony dynamics and fitness in a colonial tent-web spider Cyrtophora citricola. Front. Ecol. Evol. 2021, 9, 725647. [Google Scholar] [CrossRef]
- Uetz, G.W. The “ricochet effect” and prey capture in colonial spiders. Oecologia 1989, 81, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Su, I.; Buehler, M.J. Mesomechanics of a three-dimensional spider web. J. Mech. Phys. Solids 2020, 144, 104096. [Google Scholar] [CrossRef]
- Grinsted, L.; Lubin, Y. Spiders: Evolution of group living and social behavior. In Encyclopedia of Animal Behavior; Choe, J.C., Ed.; Academic Press: London, UK, 2019; pp. 632–640. [Google Scholar] [CrossRef]
- Sunderland, K. Mechanisms underlying the effects of spiders on pest populations. J. Arachnol. 1999, 27, 308–316. [Google Scholar]
- Wise, D.H. Cannibalism, food limitation, intraspecific competition, and the regulation of spider populations. Annu. Rev. Entomol. 2006, 51, 441–465. [Google Scholar] [CrossRef]
- Nyffeler, M. Prey selection of spiders in the field. Am. Arachnol. Soc. 1999, 27, 317–324. [Google Scholar]
- Marc, P.; Canard, A.; Ysnel, F. Spiders (Araneae) useful for pest limitation and bioindication. Agric. Ecosyst. Environ. 1999, 74, 229–273. [Google Scholar] [CrossRef]
- Grinsted, L.; Deutsch, E.K.; Jimenez-Tenorio, M.; Lubin, Y. Evolutionary drivers of group foraging: A new framework for investigating variance in food intake and reproduction. Evolution 2019, 73, 2106–2121. [Google Scholar] [CrossRef]
- Chuang, A.; Leppanen, C. Location and web substrate records of Cyrtophora citricola (Araneae: Araneidae) in southern Spain, including Tenerife. Rev. Ibérica Aracnol. 2018, 33, 89–100. [Google Scholar]
- Riechert, S.E.; Lockley, T. Spiders as biological control agents. Annu. Rev. Entomol. 1984, 29, 299–320. [Google Scholar] [CrossRef]
- Yip, E.C.; Berner-Aharon, N.; Smith, D.R.; Lubin, Y. Coy males and seductive females in the sexually cannibalistic colonial spider, Cyrtophora citricola. PLoS ONE 2016, 11, e0155433. [Google Scholar] [CrossRef] [PubMed]
- Grinsted, L.; Schou, M.F.; Settepani, V.; Holm, C.; Bird, T.L.; Bilde, T. Prey to predator body size ratio in the evolution of cooperative hunting—A social spider test case. Dev. Genes Evol. 2020, 230, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Ersts, P.J. DotDotGoose, Version 1.5.3; American Museum of Natural History: New York, NY, USA, 2022. Available online: https://biodiversityinformatics.amnh.org/open_source/dotdotgoose(accessed on 17 October 2022).
- RStudio Team. RStudio: Integrated Development Environment for R; PBC: Boston, MA, USA, 2022. [Google Scholar]
- Branthôme, F.-X. Spain: The Other European Giant. Available online: https://www.tomatonews.com/en/spain-the-other-european-giant_2_264.html (accessed on 1 February 2022).
- Visser, D.; Uys, V.M.; Nieuwenhuis, R.J.; Pieterse, W. First records of the tomato leaf miner Tuta absoluta (Meyrick, 1917) (lepidoptera: Gelechiidae) in South Africa. BioInvasions Rec. 2017, 6, 301–305. [Google Scholar] [CrossRef]
- Tellier, A.; Brown, J.K.; Boots, M.; John, S. Theory of host-parasite coevolution: From ecology to genomics. eLS 2021, 2, 1–10. [Google Scholar] [CrossRef]
- Riechert, S.E. The hows and whys of successful pest suppression by spiders: Insights from case studies. J. Arachnol. 1999, 27, 387–396. [Google Scholar]
- Michalko, R.; Pekár, S.; Dul’a, M.; Entling, M.H. Global patterns in the biocontrol efficacy of spiders: A meta-analysis. Glob. Ecol. Biogeogr. 2019, 28, 1366–1378. [Google Scholar] [CrossRef]
- Edwards, G.B. Cyrtophora citricola (Araneae: Araneidae): A colonial tentweb orbweaver established in Florida. Entomol. Circ. Fla. Dept. Agric. Consum. Serv. 2006, 411, 1–5. [Google Scholar]
- Stange, L.A. A colonial tentweb orbweaver Cyrtophora citricola. Edis 2013, 2013, 1–6. [Google Scholar] [CrossRef]
- Diaz, C.; Maksuta, D.; Amarpuri, G.; Tanikawa, A.; Miyashita, T.; Dhinojwala, A.; Blackledge, T.A. The moth specialist spider Cyrtarachne akirai uses prey scales to increase adhesion. J. R. Soc. Interface 2020, 17, 20190792. [Google Scholar] [CrossRef] [Green Version]
- Ben Husin, T.O.; Port, G.R. Efficacy of entomopathogenic nematodes against Tuta absoluta. Biol. Control 2021, 160, 104699. [Google Scholar] [CrossRef]
- Mishra, A.; Rastogi, N. Unraveling the roles of solitary and social web-making spiders in perennial ecosystems: Influence on pests and beneficials. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 567–576. [Google Scholar] [CrossRef]
- Grinsted, L.; Agnarsson, I.; Bilde, T. Subsocial behaviour and brood adoption in mixed-species colonies of two theridiid spiders. Naturwissenschaften 2012, 99, 1021–1030. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roberts-McEwen, T.A.; Deutsch, E.K.; Mowery, M.A.; Grinsted, L. Group-Living Spider Cyrtophora citricola as a Potential Novel Biological Control Agent of the Tomato Pest Tuta absoluta. Insects 2023, 14, 34. https://doi.org/10.3390/insects14010034
Roberts-McEwen TA, Deutsch EK, Mowery MA, Grinsted L. Group-Living Spider Cyrtophora citricola as a Potential Novel Biological Control Agent of the Tomato Pest Tuta absoluta. Insects. 2023; 14(1):34. https://doi.org/10.3390/insects14010034
Chicago/Turabian StyleRoberts-McEwen, Thomas A., Ella K. Deutsch, Monica A. Mowery, and Lena Grinsted. 2023. "Group-Living Spider Cyrtophora citricola as a Potential Novel Biological Control Agent of the Tomato Pest Tuta absoluta" Insects 14, no. 1: 34. https://doi.org/10.3390/insects14010034
APA StyleRoberts-McEwen, T. A., Deutsch, E. K., Mowery, M. A., & Grinsted, L. (2023). Group-Living Spider Cyrtophora citricola as a Potential Novel Biological Control Agent of the Tomato Pest Tuta absoluta. Insects, 14(1), 34. https://doi.org/10.3390/insects14010034