


Light Pollution Disrupts Seasonal Differences in the Daily Activity and Metabolic Profiles of the Northern House Mosquito, Culex pipiens


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Fly Box Construction
2.3. Measuring Locomotor Activity
2.4. Metabolic Assays
2.5. Data Analysis
3. Results
3.1. Survival
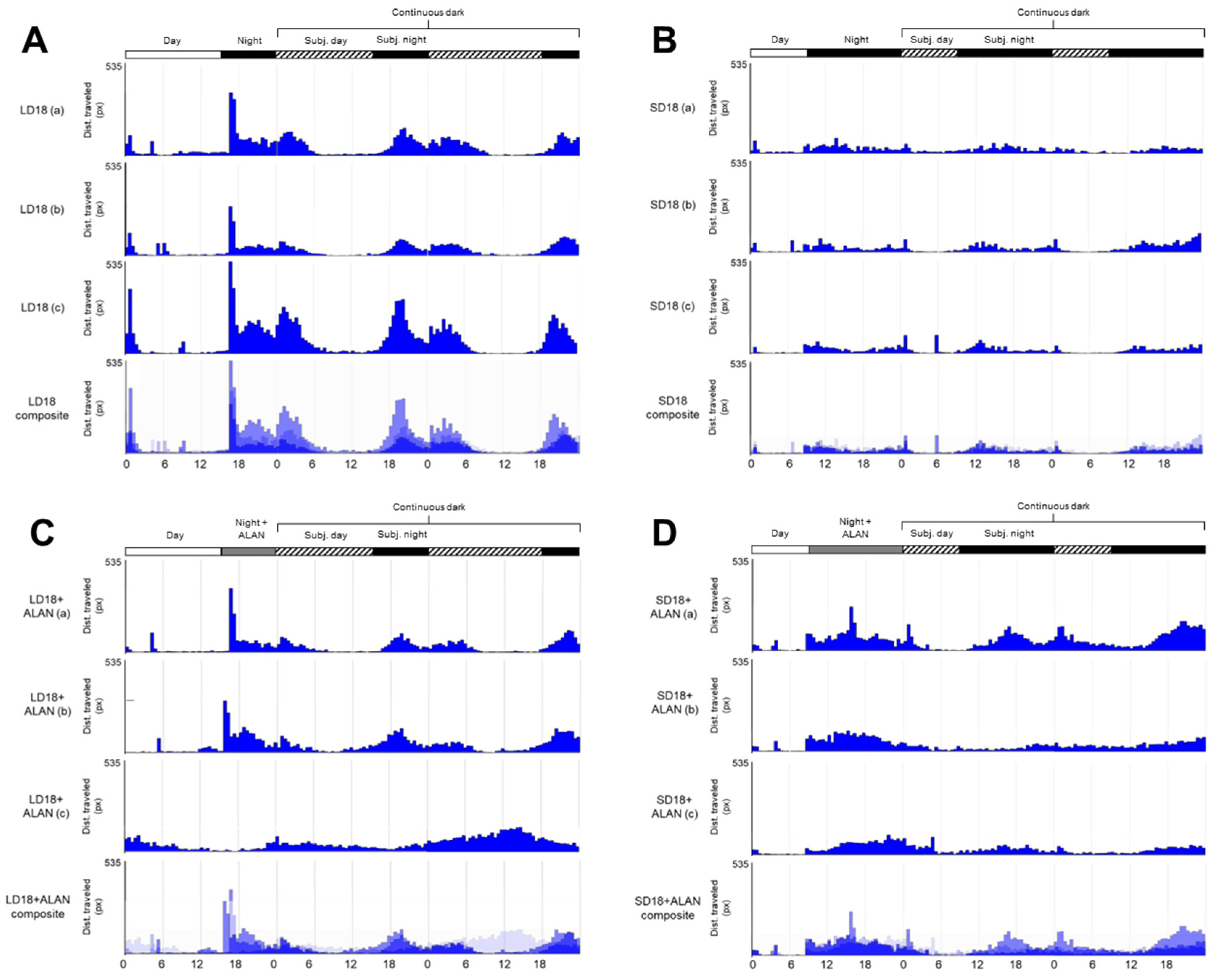
3.2. Activity Analysis
3.3. Circadian Periodicity
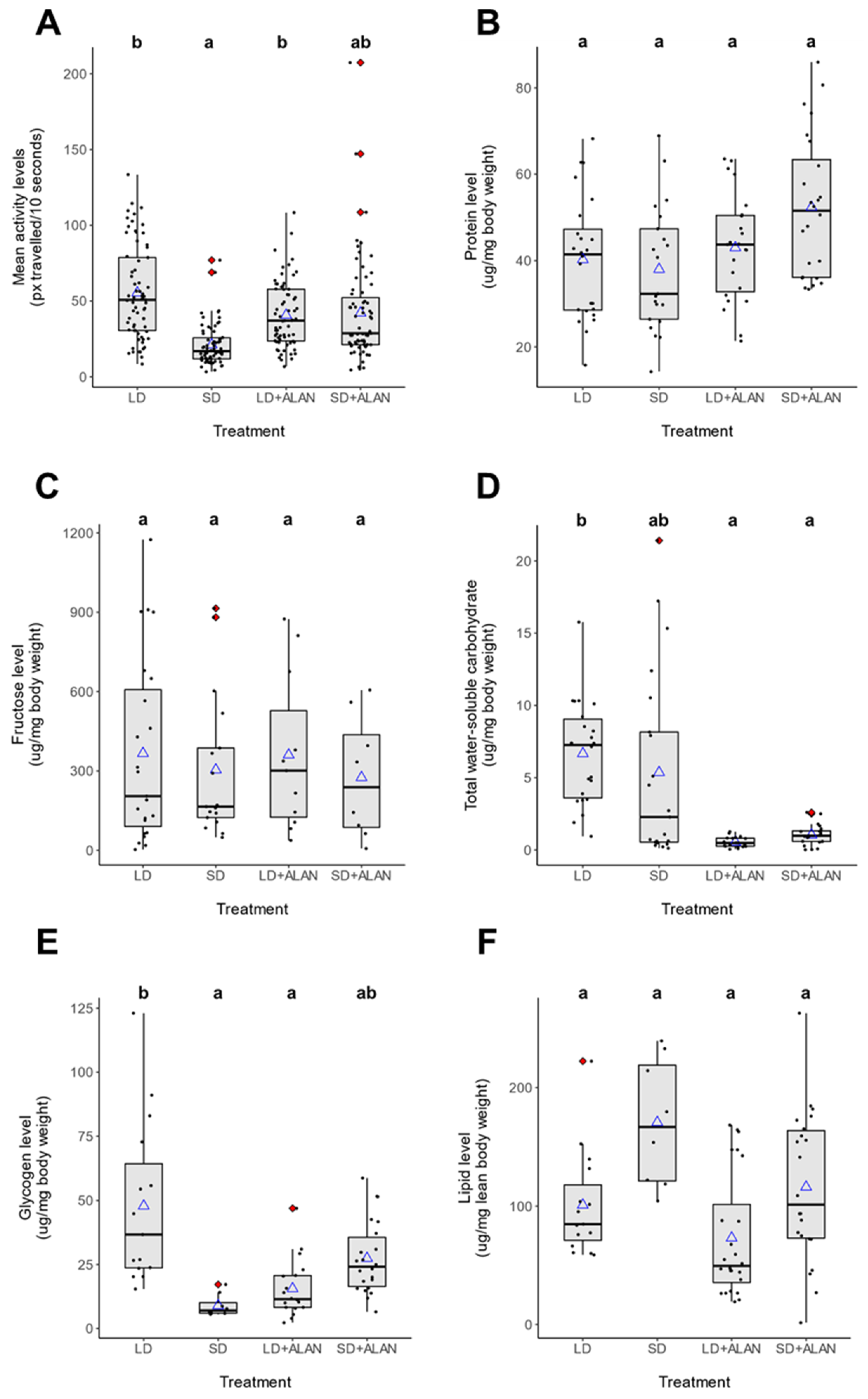
3.4. Metabolic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meireles-Filho, A.C.A.; Kyriacou, C.P. Circadian Rhythms in Insect Disease Vectors. Mem. Inst. Oswaldo Cruz 2013, 108, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glossop, N.R.; Lyons, L.C.; Hardin, P.E. Interlocked Feedback Loops within the Drosophila Circadian Oscillator. Science 1999, 286, 766–768. [Google Scholar] [CrossRef] [PubMed]
- Hardin, P.E.; Hall, J.C.; Rosbash, M. Feedback of the Drosophila Period Gene Product on Circadian Cycling of Its Messenger RNA Levels. Nature 1990, 343, 536–540. [Google Scholar] [CrossRef]
- Nitabach, M.N.; Taghert, P.H. Organization of the Drosophila Circadian Control Circuit. Curr. Biol. CB 2008, 18, R84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vosshall, L.B.; Young, M.W. Circadian Rhythms in Drosophila Can Be Driven by Period Expression in a Restricted Group of Central Brain Cells. Neuron 1995, 15, 345–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grima, B.; Chélot, E.; Xia, R.; Rouyer, F. Morning and Evening Peaks of Activity Rely on Different Clock Neurons of the Drosophila Brain. Nature 2004, 431, 869–873. [Google Scholar] [CrossRef]
- Stoleru, D.; Peng, Y.; Agosto, J.; Rosbash, M. Coupled Oscillators Control Morning and Evening Locomotor Behaviour of Drosophila. Nature 2004, 431, 862–868. [Google Scholar] [CrossRef]
- Miyasako, Y.; Umezaki, Y.; Tomioka, K. Separate Sets of Cerebral Clock Neurons Are Responsible for Light and Temperature Entrainment of Drosophila Circadian Locomotor Rhythms. J. Biol. Rhythms 2007, 22, 115–126. [Google Scholar] [CrossRef]
- Murad, A.; Emery-Le, M.; Emery, P. A subset of dorsal neurons modulates circadian behavior and light responses in Drosophila. Neuron 2007, 53, 689–701. [Google Scholar] [CrossRef] [Green Version]
- Veleri, S.; Brandes, C.; Helfrich-Förster, C.; Hall, J.C.; Stanewsky, R. A Self-Sustaining, Light-Entrainable Circadian Oscillator in the Drosophila Brain. Curr. Biol. CB 2003, 13, 1758–1767. [Google Scholar] [CrossRef]
- Helfrich-Förster, C.; Nitabach, M.; Holmes, T. Insect Circadian Clock Outputs. Essays Biochem. 2011, 49, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Shafer, O.T.; Yao, Z. Pigment-Dispersing Factor Signaling and Circadian Rhythms in Insect Locomotor Activity. Curr. Opin. Insect Sci. 2014, 1, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Závodská, R.; von Wowern, G.; Löfstedt, C.; Rosén, W.; Sauman, I. The Release of a Pheromonotropic Neuropeptide, PBAN, in the Turnip Moth Agrotis segetum, Exhibits a Circadian Rhythm. J. Insect Physiol. 2009, 55, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Mark, B.; Bustos-González, L.; Cascallares, G.; Conejera, F.; Ewer, J. The Circadian Clock Gates Drosophila Adult Emergence by Controlling the Timecourse of Metamorphosis. Proc. Natl. Acad. Sci. USA 2021, 118, e2023249118. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Zheng, X.; Sehgal, A. Regulation of Feeding and Metabolism by Neuronal and Peripheral Clocks in Drosophila. Cell Metab. 2008, 8, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Dimopoulos, G. Molecular Analysis of Photic Inhibition of Blood-Feeding in Anopheles gambiae. BMC Physiol. 2008, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Danks, H.V. Insect Dormancy: An Ecological Perspective; Biological Survey of Canada (Terrestrial Arthropods): Ottawa, ON, Canada, 1987. [Google Scholar]
- Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press: Oxford, UK; New York, NY, USA, 1985; ISBN 978-0-19-503635-0. [Google Scholar]
- Eldridge, B.F. The Effect of Temperature and Photoperiod on Blood-Feeding and Ovarian Development in Mosquitoes of the Culex pipiens Complex. Am. J. Trop. Med. Hyg. 1968, 17, 133–140. [Google Scholar] [CrossRef]
- Spielman, A.; Wong, J. Environmental Control of Ovarian Diapause in Culex pipiens. Ann. Entomol. Soc. Am. 1973, 66, 905–907. [Google Scholar] [CrossRef]
- Sanburg, L.L.; Larsen, J.R. Effect of Photoperiod and Temperature on Ovarian Development in Culex pipiens pipiens. J. Insect Physiol. 1973, 19, 1173–1190. [Google Scholar] [CrossRef]
- Spielman, A. Effect of Synthetic Juvenile Hormone on Ovarian Diapause of Culex pipiens Mosquitoes. J. Med. Entomol. 1974, 11, 223–225. [Google Scholar] [CrossRef]
- Sim, C.; Denlinger, D.L. Insulin Signaling and FOXO Regulate the Overwintering Diapause of the Mosquito Culex pipiens. Proc. Natl. Acad. Sci. USA 2008, 105, 6777–6781. [Google Scholar] [CrossRef] [Green Version]
- Sim, C.; Kang, D.S.; Kim, S.; Bai, X.; Denlinger, D.L. Identification of FOXO Targets That Generate Diverse Features of the Diapause Phenotype in the Mosquito Culex pipiens. Proc. Natl. Acad. Sci. USA 2015, 112, 3811–3816. [Google Scholar] [CrossRef] [Green Version]
- Sim, C.; Denlinger, D.L. Transcription Profiling and Regulation of Fat Metabolism Genes in Diapausing Adults of the Mosquito Culex pipiens. Physiol. Genom. 2009, 39, 202–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowen, M.F.; Davis, E.E.; Haggart, D.A. A Behavioural and Sensory Analysis of Host-Seeking Behaviour in the Diapausing Mosquito Culex pipiens. J. Insect Physiol. 1988, 34, 805–813. [Google Scholar] [CrossRef]
- Bowen, M.F. Patterns of Sugar Feeding in Diapausing and Nondiapausing Culex pipiens (Diptera: Culicidae) Females. J. Med. Entomol. 1992, 29, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Meuti, M.E.; Stone, M.; Ikeno, T.; Denlinger, D.L. Functional Circadian Clock Genes Are Essential for the Overwintering Diapause of the Northern House Mosquito, Culex pipiens. J. Exp. Biol. 2015, 218, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Bunning, E.J. Die Endonome Tagesrhythmik Als Grundlage Der Photoperiodischen Reaktion. Ber. Deut. Bot. Ges. 1937, 54, 590–607. [Google Scholar]
- Pittendrigh, C.S.; Minis, D.H. The Entrainment of Circadian Oscillations by Light and Their Role as Photoperiodic Clocks. Am. Nat. 1964, 98, 261–294. [Google Scholar] [CrossRef]
- Meuti, M.E.; Denlinger, D.L. Evolutionary Links Between Circadian Clocks and Photoperiodic Diapause in Insects. Integr. Comp. Biol. 2013, 53, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Lees, A.D. The Physiology of Diapause in Arthropods. Camb. Monogr. Exp. Biol. 1955, 4, 1–51. [Google Scholar]
- Ragland, G.J.; Keep, E. Comparative Transcriptomics Support Evolutionary Convergence of Diapause Responses across Insecta. Physiol. Entomol. 2017, 42, 246–256. [Google Scholar] [CrossRef] [Green Version]
- Kostál, V. Eco-Physiological Phases of Insect Diapause. J. Insect Physiol. 2006, 52, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Refinetti, R.; Lissen, G.C.; Halber, F. Procedures for Numerical Analysis of Circadian Rhythms. Biol. Rhythm Res. 2007, 38, 275–325. [Google Scholar] [CrossRef] [PubMed]
- Socha, R.; Zemek, R. Locomotor Activity in Adult Pyrrhocoris apterus (Heteroptera) in Relation to Sex, Physiological Status and Wing Dimorphism. Physiol. Entomol. 2000, 25, 383–389. [Google Scholar] [CrossRef]
- Matsunaga, K.; Takahashi, H.; Yoshida, T.; Kimura, M.T. Feeding, Reproductive and Locomotor Activities in Diapausing and Non-Diapausing Adults of Drosophila. Ecol. Res. 1995, 10, 87–93. [Google Scholar] [CrossRef]
- Veronesi, R.; Gentile, G.; Carrieri, M.; Maccagnani, B.; Stermieri, L.; Bellini, R. Seasonal Pattern of Daily Activity of Aedes caspius, Aedes detritus, Culex modestus, and Culex pipiens in the Po Delta of Northern Italy and Significance for Vector-Borne Disease Risk Assessment. J. Vector Ecol. 2012, 37, 49–61. [Google Scholar] [CrossRef]
- Mitchell, C.J. Differentiation of Host-Seeking Behavior from Blood-Feeding Behavior in Overwintering Culex pipiens (Diptera: Culicidae) and Observations on Gonotrophic Dissociation. J. Med. Entomol. 1983, 20, 157–163. [Google Scholar] [CrossRef]
- Andreadis, T.G. The Contribution of Culex pipiens Complex Mosquitoes to Transmission and Persistence of West Nile Virus in North America. J. Am. Mosq. Control Assoc. 2012, 28, 137–151. [Google Scholar] [CrossRef]
- West Nile Virus |West Nile Virus| CDC. Available online: https://www.cdc.gov/westnile/index.html (accessed on 7 November 2022).
- Bugbee, L.M.; Forte, L.R. The Discovery of West Nile Virus in Overwintering Culex pipiens (Diptera: Culicidae) Mosquitoes in Lehigh County, Pennsylvania. J. Am. Mosq. Control Assoc. 2004, 20, 326–327. [Google Scholar]
- Farajollahi, A.; Crans, W.J.; Bryant, P.; Wolf, B.; Burkhalter, K.L.; Godsey, M.S.; Aspen, S.E.; Nasci, R.S. Detection of West Nile Viral RNA from an Overwintering Pool of Culex pipiens pipiens(Diptera: Culicidae) in New Jersey, 2003. J. Med. Entomol. 2005, 42, 490–494. [Google Scholar] [CrossRef] [Green Version]
- Andreadis, T.G.; Armstrong, P.M.; Bajwa, W.I. Studies on Hibernating Populations of Culex Pipiens from a West Nile Virus Endemic Focus in New York City: Parity Rates and Isolation of West Nile Virus. J. Am. Mosq. Control Assoc. 2010, 26, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Bumgarner, J.R.; Nelson, R.J. Light at Night and Disrupted Circadian Rhythms Alter Physiology and Behavior. Integr. Comp. Biol. 2021, 61, 1160–1169. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.J.; Visser, M.E.; Hölker, F. The Biological Impacts of Artificial Light at Night: The Research Challenge. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140133. [Google Scholar] [CrossRef] [PubMed]
- Falchi, F.; Cinzano, P.; Duriscoe, D.; Kyba, C.C.M.; Elvidge, C.D.; Baugh, K.; Portnov, B.A.; Rybnikova, N.A.; Furgoni, R. The New World Atlas of Artificial Night Sky Brightness. Sci. Adv. 2016, 2, e1600377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaston, K.J.; Davies, T.W.; Nedelec, S.L.; Holt, L.A. Impacts of Artificial Light at Night on Biological Timings. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 49–68. [Google Scholar] [CrossRef]
- Dominoni, D.M.; Borniger, J.C.; Nelson, R.J. Light at Night, Clocks and Health: From Humans to Wild Organisms. Biol. Lett. 2016, 12, 20160015. [Google Scholar] [CrossRef] [Green Version]
- Saunders, D. Dormancy, Diapause, and the Role of the Circadian System in Insect Photoperiodism. Annu. Rev. Entomol. 2019, 65, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Gomes, E.; Rey, B.; Débias, F.; Amat, I.; Desouhant, E. Dealing with Host and Food Searching in a Diurnal Parasitoid: Consequences of Light at Night at Intra- and Trans-Generational Levels. Insect Conserv. Divers. 2021, 14, 235–246. [Google Scholar] [CrossRef]
- Rund, S.S.C.; Labb, L.F.; Benefiel, O.M.; Duffield, G.E. Artificial Light at Night Increases Aedes aegypti Mosquito Biting Behavior with Implications for Arboviral Disease Transmission. Am. J. Trop. Med. Hyg. 2020, 103, 2450–2452. [Google Scholar] [CrossRef]
- Owens, A.C.S.; Lewis, S.M. The Impact of Artificial Light at Night on Nocturnal Insects: A Review and Synthesis. Ecol. Evol. 2018, 8, 11337–11358. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.; Quintanilla-Ahumada, D.; Anguita, C.; Manríquez, P.H.; Widdicombe, S.; Pulgar, J.; Silva-Rodríguez, E.A.; Miranda, C.; Manríquez, K.; Quijón, P.A. Artificial Light Pollution at Night (ALAN) Disrupts the Distribution and Circadian Rhythm of a Sandy Beach Isopod. Environ. Pollut. 2019, 248, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Levy, K.; Wegrzyn, Y.; Efronny, R.; Barnea, A.; Ayali, A. Lifelong Exposure to Artificial Light at Night Impacts Stridulation and Locomotion Activity Patterns in the Cricket Gryllus bimaculatus. Proc. R. Soc. B Biol. Sci. 2021, 288, 20211626. [Google Scholar] [CrossRef] [PubMed]
- Luarte, T.; Bonta, C.C.; Silva-Rodriguez, E.A.; Quijón, P.A.; Miranda, C.; Farias, A.A.; Duarte, C. Light Pollution Reduces Activity, Food Consumption and Growth Rates in a Sandy Beach Invertebrate. Environ. Pollut. Barking Essex 1987 2016, 218, 1147–1153. [Google Scholar] [CrossRef]
- Honnen, A.-C.; Kypke, J.L.; Hölker, F.; Monaghan, M.T. Artificial Light at Night Influences Clock-Gene Expression, Activity, and Fecundity in the Mosquito Culex pipiens f. molestus. Sustainability 2019, 11, 6220. [Google Scholar] [CrossRef] [Green Version]
- Mukai, A.; Yamaguchi, K.; Goto, S.G. Urban Warming and Artificial Light Alter Dormancy in the Flesh Fly. R. Soc. Open Sci. 2021, 8, 210866. [Google Scholar] [CrossRef] [PubMed]
- van Geffen, K.G.; van Grunsven, R.H.A.; van Ruijven, J.; Berendse, F.; Veenendaal, E.M. Artificial Light at Night Causes Diapause Inhibition and Sex-Specific Life History Changes in a Moth. Ecol. Evol. 2014, 4, 2082–2089. [Google Scholar] [CrossRef]
- Westby, K.M.; Medley, K.A. Cold Nights, City Lights: Artificial Light at Night Reduces Photoperiodically Induced Diapause in Urban and Rural Populations of Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2020, 57, 1694–1699. [Google Scholar] [CrossRef] [PubMed]
- Fyie, L.R.; Gardiner, M.M.; Meuti, M.E. Artificial Light at Night Alters the Seasonal Responses of Biting Mosquitoes. J. Insect Physiol. 2021, 129, 104194. [Google Scholar] [CrossRef] [PubMed]
- Robich, R.M.; Denlinger, D.L. Diapause in the Mosquito Culex pipiens Evokes a Metabolic Switch from Blood Feeding to Sugar Gluttony. Proc. Natl. Acad. Sci. USA 2005, 102, 15912–15917. [Google Scholar] [CrossRef] [Green Version]
- Loetti, V.; Schweigmann, N.; Burroni, N. Development rates, larval survivorship and wing length of Culex pipiens (Diptera: Culicidae) at constant temperatures. J. Nat. Hist. 2011, 45, 2203–2213. [Google Scholar] [CrossRef]
- Gaston, K.J.; Bennie, J.; Davies, T.W.; Hopkins, J. The ecological impacts of nighttime light pollution: A mechanistic appraisal. Biol. Rev. 2013, 88, 912–927. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, S.; Hossler, K.; Meyer, L. Artificial lighting at night alters aquatic-riparian invertebrate food webs. Ecol. Appl. 2019, 29, e01821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva Araujo, M.; Guo, F.; Rosbash, M. Video Recording Can Conveniently Assay Mosquito Locomotor Activity. Sci. Rep. 2020, 10, 4994. [Google Scholar] [CrossRef]
- Gilestro, G.F.; Cirelli, C. PySolo: A Complete Suite for Sleep Analysis in Drosophila. Bioinforma. Oxf. Engl. 2009, 25, 1466–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecsey, C. Research Tools: Sleep and Circadian Analysis MATLAB Program. Available online: https://academics.skidmore.edu/blogs/cvecsey/?page_id=57 (accessed on 13 June 2022).
- Foray, V.; Pelisson, P.F.; Bel-Venner, M.C.; Desouhant, E.; Venner, S.; Menu, F.; Giron, D.; Rey, B. A handbook for uncovering the complete energetic budget in insects: The Van Handel’s method (1985) revisited. Physiol. Entomol. 2012, 37, 295–302. [Google Scholar] [CrossRef]
- Rosato, E.; Kyriacou, C. Analysis of locomotor activity rhythms in Drosophila. Nat. Protoc. 2006, 1, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Statistics and Computing; Springer: New York, NY, USA, 2002; ISBN 978-1-4419-3008-8. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 97, 1–48. [Google Scholar] [CrossRef]
- Ludecke, D. Sjstats: Statistical Functions for Regression Models (Version 0.18.2). Zenodo 2022. [Google Scholar] [CrossRef]
- Huck, D.T.; Klein, M.S.; Meuti, M.E. Determining the effects of nutrition on the reproductive physiology of male mosquitoes. J. Insect Physiol. 2021, 129, 104191. [Google Scholar] [CrossRef]
- Fox, J. Effect displays in R for generalised linear models. J. Stat. Softw. 2003, 8, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage Publications Inc.: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Kim, M.; Denlinger, D.L. Decrease in Expression of Beta-Tubulin and Microtubule Abundance in Flight Muscles during Diapause in Adults of Culex pipiens. Insect Mol. Biol. 2009, 18, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Stegwee, D.; Kimmel, E.C.; de Boer, J.A.; Henstra, S. Hormonal control of reversible degeneration of flight muscle in the Colorado potato beetle, Leptinotarsa decemlineata Say (Coleoptera). J. Cell Biol. 1963, 19, 519–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolda, H.; Denlinger, D.L. Diapause in a Large Aggregation of a Tropical Beetle. Ecol. Entomol. 1984, 9, 217–230. [Google Scholar] [CrossRef]
- Hunt, J.H.; Kensinger, B.J.; Kossuth, J.A.; Henshaw, M.T.; Norberg, K.; Wolschin, F.; Amdam, G.V. A Diapause Pathway Underlies the Gyne Phenotype in Polistes Wasps, Revealing an Evolutionary Route to Caste-Containing Insect Societies. Proc. Natl. Acad. Sci. USA 2007, 104, 14020–14025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koopmanschap, A.B.; Lammers, J.H.M.; de Kort, C.A.D. The Structure of the Gene Encoding Diapause Protein 1 of the Colorado Potato Beetle (Leptinotarsa decemlineata). J. Insect Physiol. 1995, 41, 509–518. [Google Scholar] [CrossRef]
- Lewis, D.K.; Spurgeon, D.; Sappington, T.W.; Keeley, L.L. A Hexamerin Protein, AgSP-1, Is Associated with Diapause in the Boll Weevil. J. Insect Physiol. 2002, 48, 887–901. [Google Scholar] [CrossRef]
- Kang, D.S.; Cotten, M.A.; Denlinger, D.L.; Sim, C. Comparative Transcriptomics Reveals Key Gene Expression Differences between Diapausing and Non-Diapausing Adults of Culex pipiens. PLoS ONE 2016, 11, e0154892. [Google Scholar] [CrossRef] [Green Version]
- Deak, I.I. Use of Drosophila Mutants to Investigate the Effect of Disuse on the Maintenance of Muscle. J. Insect Physiol. 1976, 22, 1159–1165. [Google Scholar] [CrossRef]
- Anderson, M. Ultrastructural Evidence for Disuse Atrophy in Insect Skeletal Muscle. Cell Tissue Res. 1979, 203, 503–506. [Google Scholar] [CrossRef]
- Piazza, N.; Gosangi, B.; Devilla, S.; Arking, R.; Wessells, R. Exercise-Training in Young Drosophila Melanogaster Reduces Age-Related Decline in Mobility and Cardiac Performance. PLoS ONE 2009, 4, e5886. [Google Scholar] [CrossRef]
- Zhou, G.; Miesfeld, R.L. Energy Metabolism during Diapause in Culex pipiens Mosquitoes. J. Insect Physiol. 2009, 55, 40–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, B.J. Linking Energetics and Overwintering in Temperate Insects. J. Therm. Biol. 2015, 54, 5–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; He, L.; Sang, W.; Wang, L.; Huang, Q.; Lei, C. Potential Role of Lysine Succinylation in the Response of Moths to Artificial Light at Night Stress. Ecotoxicol. Environ. Saf. 2021, 220, 112334. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-R.; Wei, W.-L.; Tzeng, D.T.W.; Owens, A.C.S.; Tang, H.-C.; Wu, C.-S.; Lin, S.-S.; Zhong, S.; Yang, E.-C. Effects of Artificial Light at Night (ALAN) on Gene Expression of Aquatica ficta Firefly Larvae. Environ. Pollut. 2021, 281, 116944. [Google Scholar] [CrossRef]
- van Langevelde, F.; van Grunsven, R.H.A.; Veenendaal, E.M.; Fijen, T.P.M. Artificial Night Lighting Inhibits Feeding in Moths. Biol. Lett. 2017, 13, 20160874. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, C.J.; Briegel, H. Inability of Diapausing Culex pipiens (Diptera: Culicidae) to Use Blood for Producing Lipid Reserves for Overwinter Survival. J. Med. Entomol. 1989, 26, 318–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, V.; Meuti, M.E. Circadian Transcription Factors Differentially Regulate Features of the Adult Overwintering Diapause in the Northern House Mosquito, Culex pipiens. Insect Biochem. Mol. Biol. 2020, 121, 103365. [Google Scholar] [CrossRef]
- Timmerman, S.E.; Briegel, H. Larval growth and biosynthesis of reserves in mosquitoes. J. Insect Physiol. 1999, 45, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Singh, J.; Kim, S.; Hockaday, W.C.; Sim, C.; Kim, S.J. Solid-state NMR reveals differential carbohydrate utilization in diapausing Culex pipiens. Sci. Rep. 2016, 6, 37350. [Google Scholar] [CrossRef] [Green Version]
- Nayar, J.K.; Sauerman, D.M., Jr. The Effects of Nutrition on Survival and Fecundity in Florida Mosquitoes Part 3. Utilization of Blood and Sugar for Fecundity1. J. Med. Entomol. 1975, 12, 220–225. [Google Scholar] [CrossRef]
- Owens, A.C.S.; Cochard, P.; Durrant, J.; Farnworth, B.; Perkin, E.K.; Seymoure, B. Light Pollution Is a Driver of Insect Declines. Biol. Conserv. 2020, 241, 108259. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolkoff, M.; Fyie, L.; Meuti, M. Light Pollution Disrupts Seasonal Differences in the Daily Activity and Metabolic Profiles of the Northern House Mosquito, Culex pipiens. Insects 2023, 14, 64. https://doi.org/10.3390/insects14010064
Wolkoff M, Fyie L, Meuti M. Light Pollution Disrupts Seasonal Differences in the Daily Activity and Metabolic Profiles of the Northern House Mosquito, Culex pipiens. Insects. 2023; 14(1):64. https://doi.org/10.3390/insects14010064
Chicago/Turabian StyleWolkoff, Matthew, Lydia Fyie, and Megan Meuti. 2023. "Light Pollution Disrupts Seasonal Differences in the Daily Activity and Metabolic Profiles of the Northern House Mosquito, Culex pipiens" Insects 14, no. 1: 64. https://doi.org/10.3390/insects14010064
APA StyleWolkoff, M., Fyie, L., & Meuti, M. (2023). Light Pollution Disrupts Seasonal Differences in the Daily Activity and Metabolic Profiles of the Northern House Mosquito, Culex pipiens. Insects, 14(1), 64. https://doi.org/10.3390/insects14010064