
Improvement in the Identification Technology for Asian Spongy Moth, Lymantria dispar Linnaeus, 1758 (Lepidoptera: Erebidae) Based on SS-COI

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Preparation of Template DNA
2.3. DNA Barcoding and Phylogenetic Relationships of Lymantria
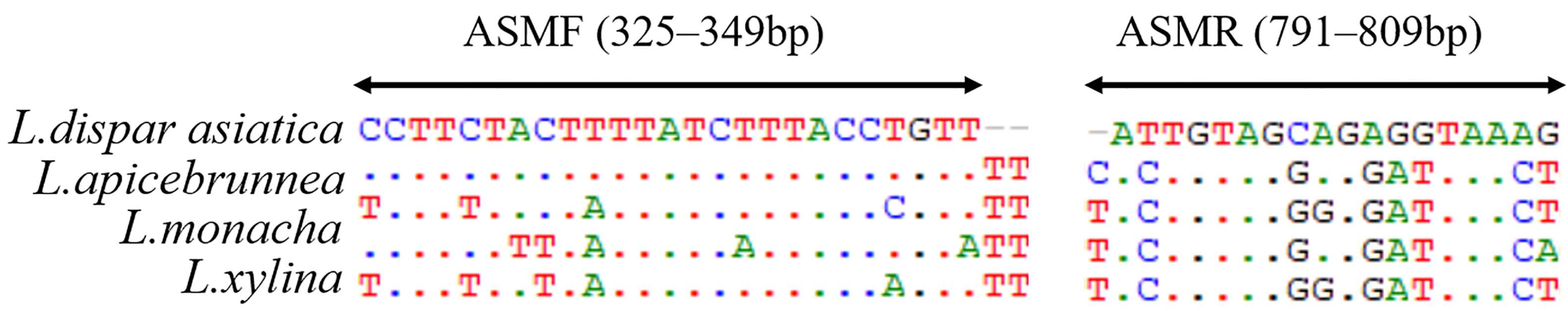
2.4. Amplification of COI and the Design of Primers Specific for ASMs
2.5. Assessment of the Specificity and Sensitivity of the SS-COI PCR Assay
2.6. Random Detection of Moths in the Wild with ASMF/ASMR
3. Results
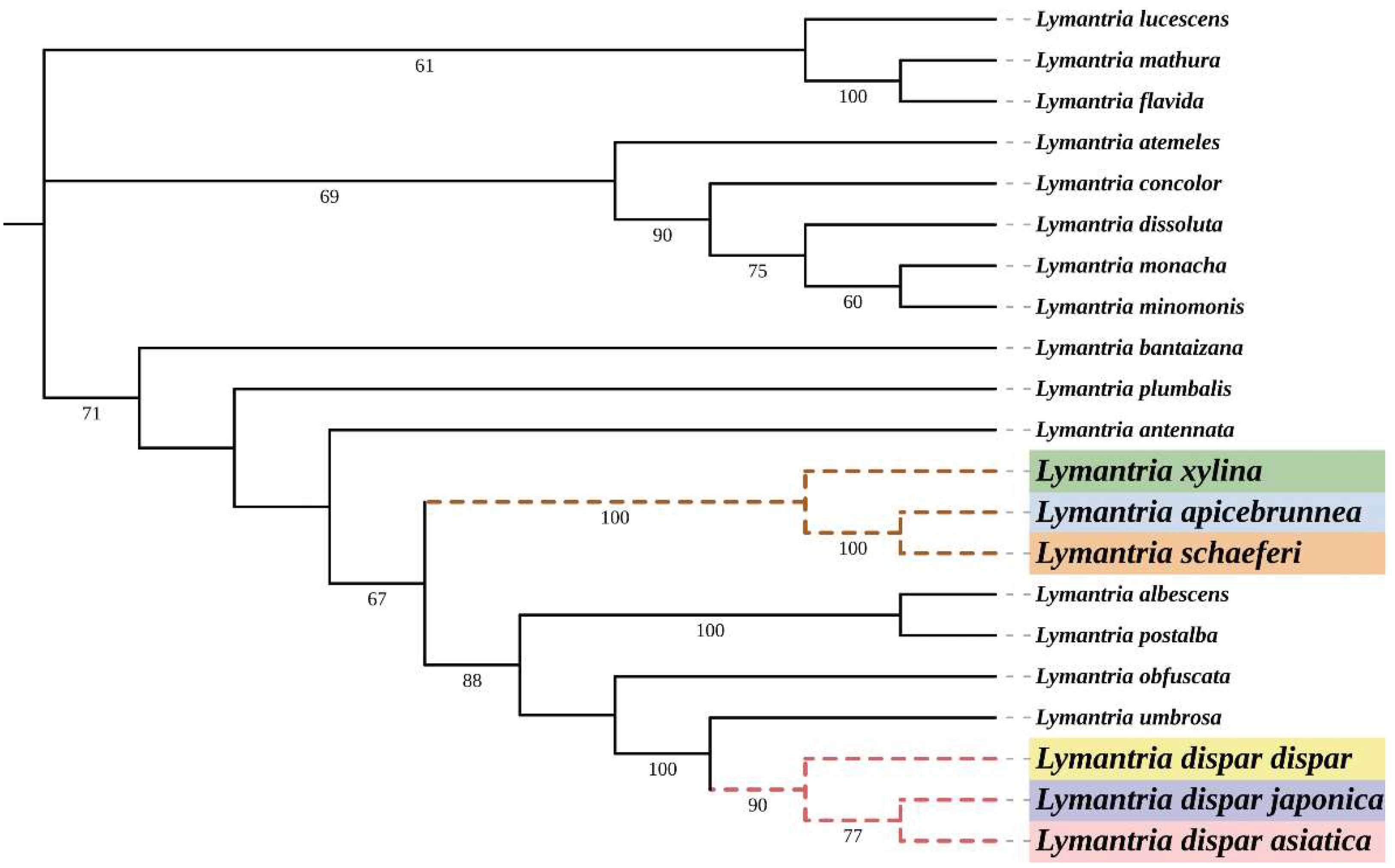
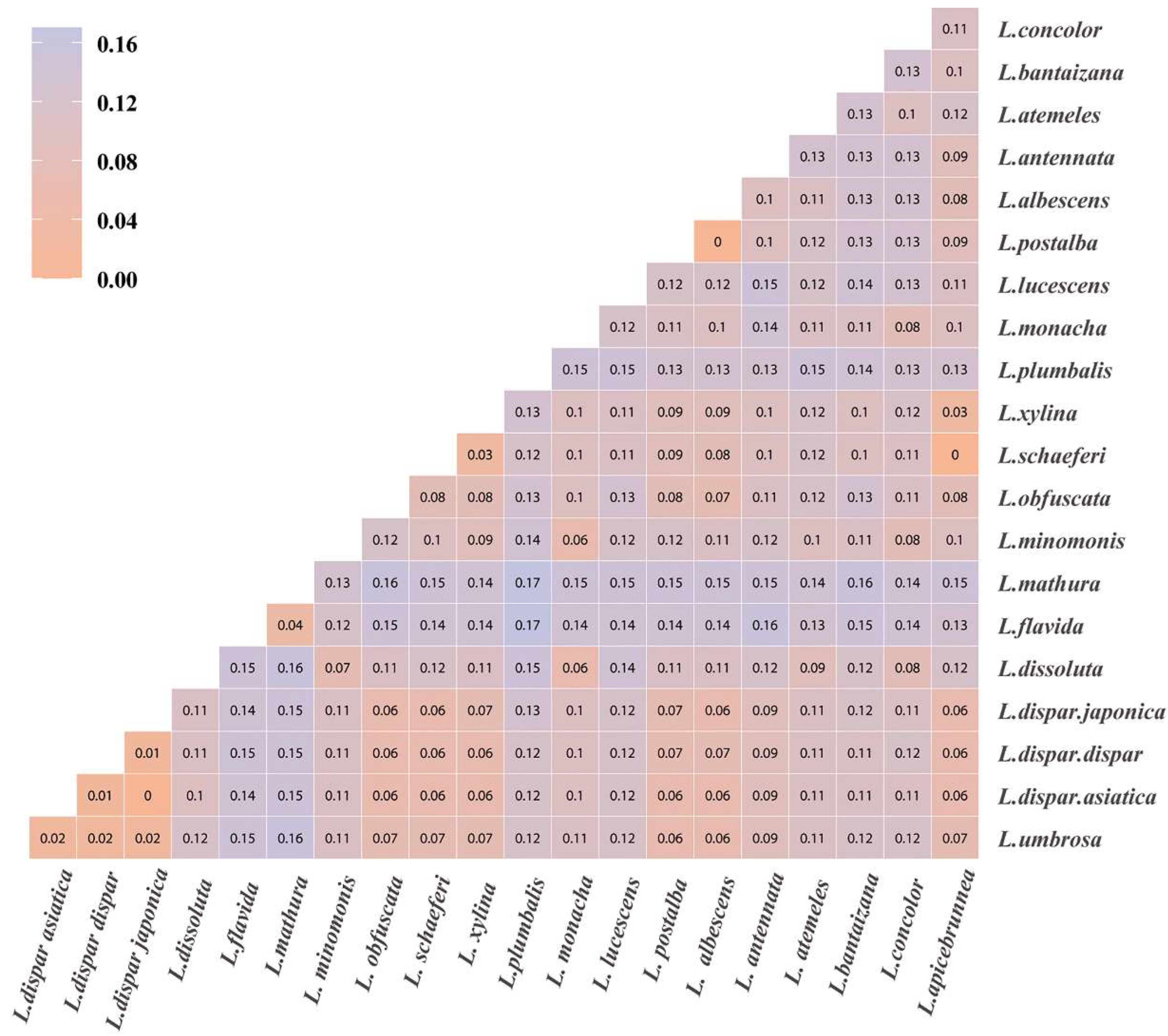
3.1. DNA Barcoding and Phylogenetic Analysis of Lymantria
3.2. Amplification of the COI Gene Using Generic Primers
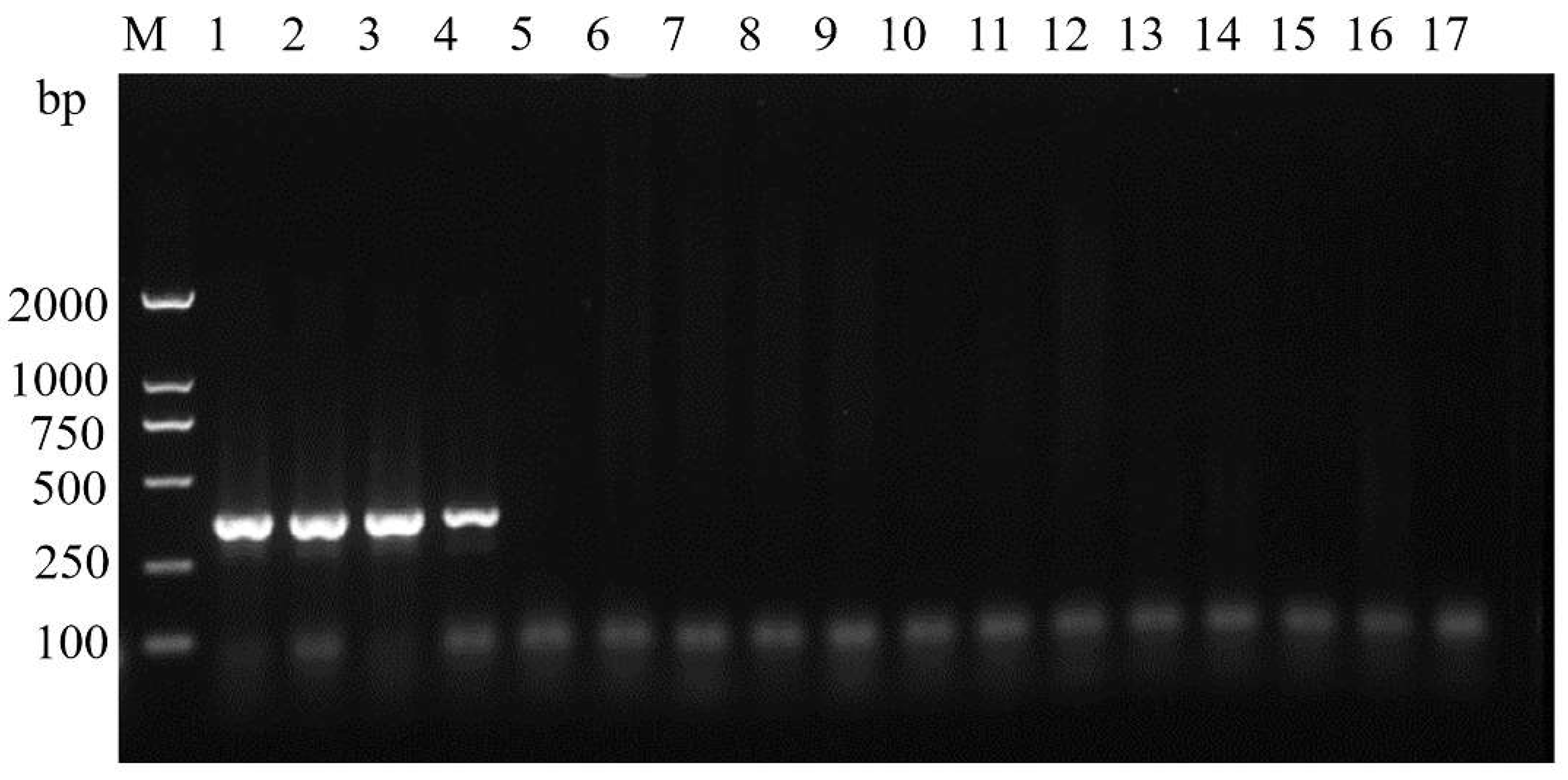
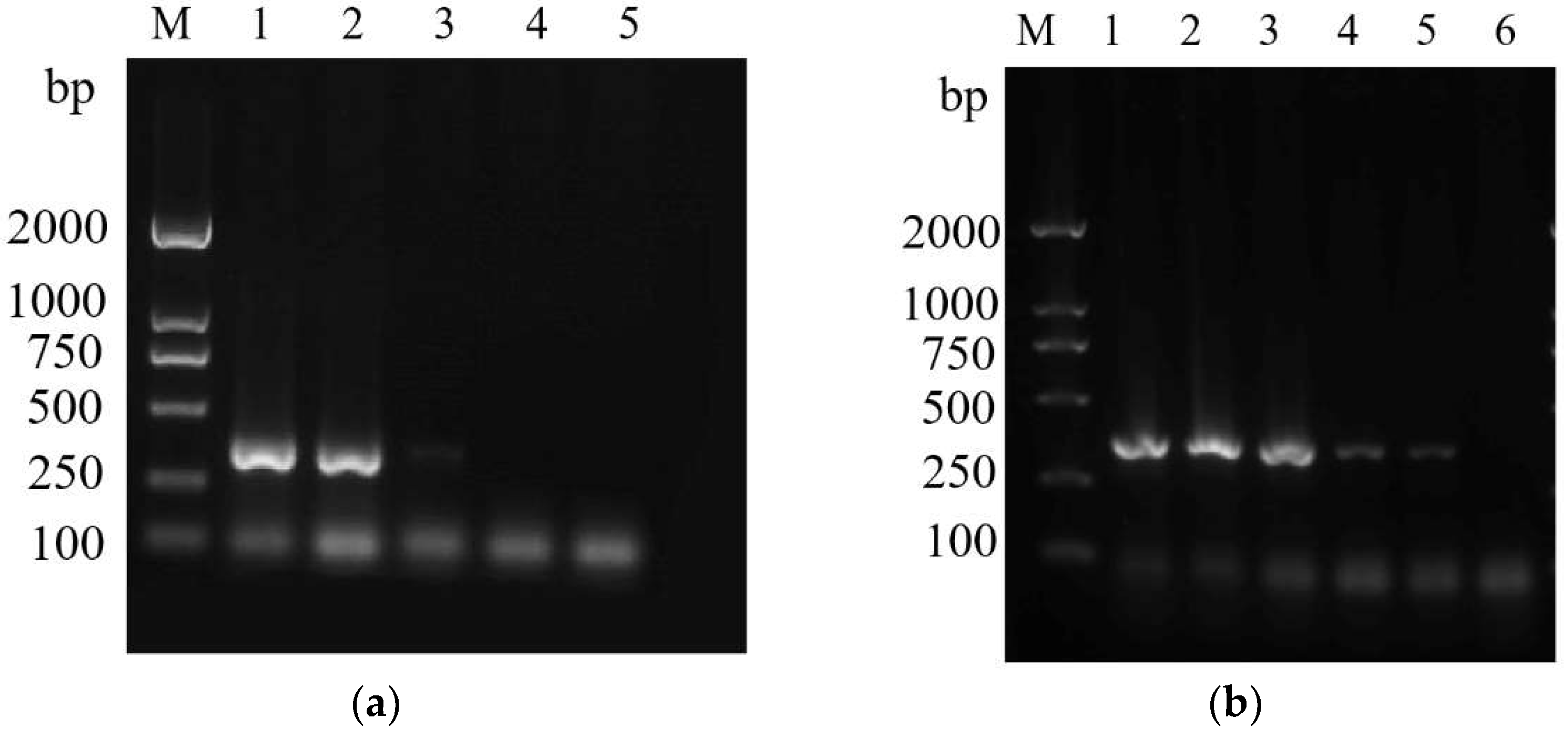
3.3. Primer Specificity Test
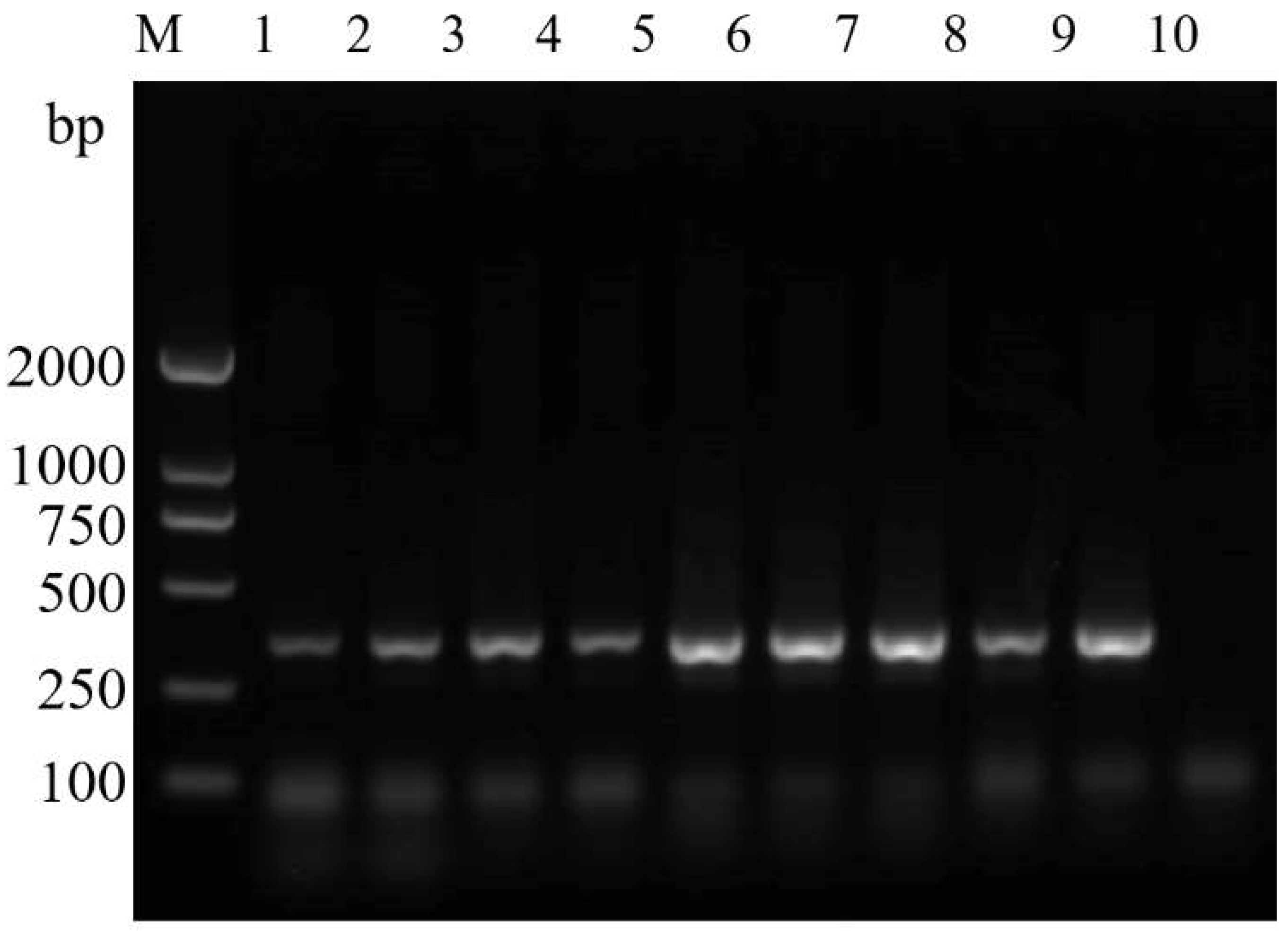
3.4. Sensitivity Test of the ASM-Specific Primers
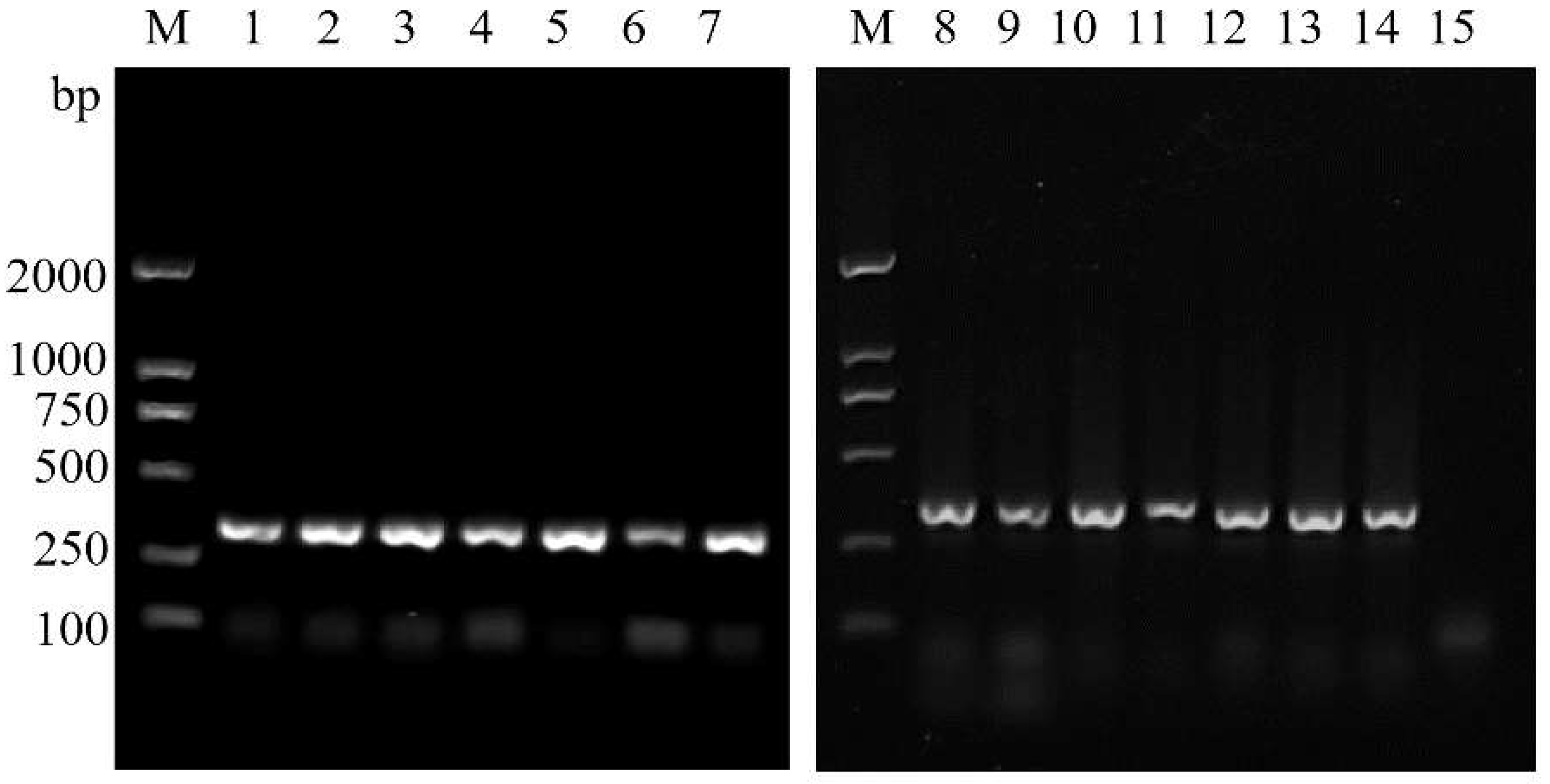
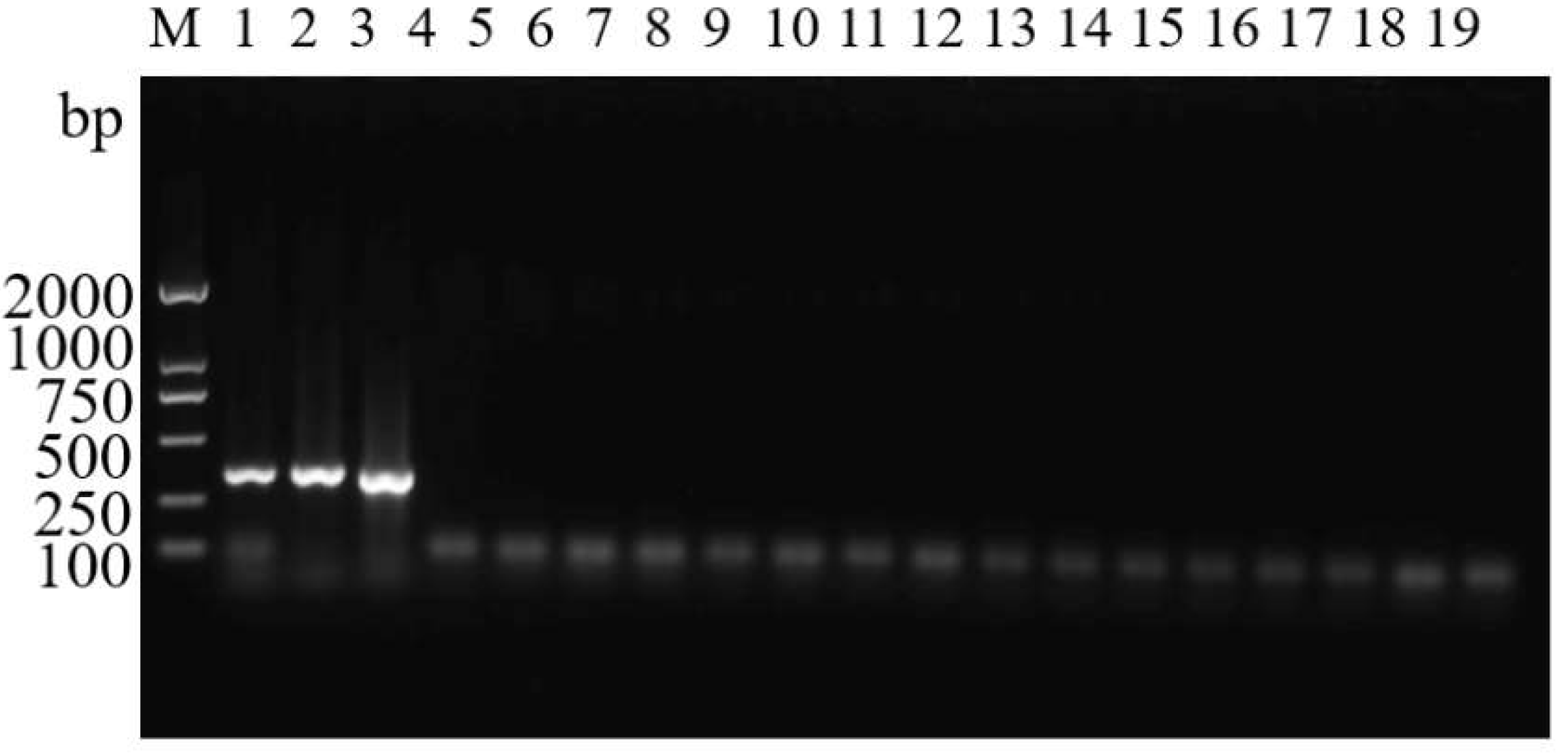
3.5. Random Detection of Moths in the Wild with ASMF/ASMR
4. Discussion
4.1. Development of Rapid and Accurate Molecular Detection Techniques Is a Necessity
4.2. For Practical Reasons, SS-COI PCR Is a Highly Efficient and Suitable Method for Ports
4.3. The Updated ASM Detection Method has Enough Specificity and Sensitivity for Application
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simberloff, D.; Martin, J.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; GarcíaBerthou, E.; Pascal, M. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacon, S.J.; Bacher, S.; Aebi, A. Gaps in border controls are related to quarantine alien insect invasions in Europe. PLoS ONE 2012, 7, e47689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Dobbs, T.T.; Brodel, C.F. Cargo aircraft as a pathway for the entry of nonindigenous pests into south Florida. Fla. Entomol. 2004, 87, 65–78. [Google Scholar] [CrossRef]
- Bradshaw, C.J.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; BarbetMassin, M.; Salles, J.; Simard, F.; Courchamp, F. Massive yet grossly underestimated global costs of invasive insects. Nat. Commun. 2016, 7, 12986. [Google Scholar] [CrossRef] [Green Version]
- Liao, M.L. Species Invasion: Ecological War without Smoke; Beijing Science Popularization Development Center: Beijing, China, 2021; p. 6. [Google Scholar]
- Simberloff, D.; Souza, L.; Nuñez, M.A.; Barrios-Garcia, M.N.; Bunn, W. The natives are restless, but not often and mostly when disturbed. Ecology 2012, 93, 598–607. [Google Scholar] [CrossRef]
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.M. Economic impacts of non-native forest insects in the continental United States. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef] [Green Version]
- Brockerhoff, E.G.; Liebhold, A.M.; Jactel, H. The ecology of forest insect invasions and advances in their management. Can. J. For. Res. 2006, 36, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Wan, F.-H.; Yang, N.-W. Invasion and management of agricultural alien insects in China. Annu. Rev. Entomol. 2016, 61, 77–98. [Google Scholar] [CrossRef]
- Zahiri, R.; Holloway, J.D.; Kitching, I.J.; Lafontaine, J.D.; Mutanen, M.; Wahlberg, N. Molecular phylogenetics of Erebidae (Lepidoptera, noctuoidea). Syst Entomol. 2012, 37, 102–124. [Google Scholar] [CrossRef]
- Pogue, M.; Schaefer, P.W. A Review of Selected Species of Lymantria (Hubner [1819]) (Lepidoptera: Noctuidae: Lymantriinae) from Subtropical and Temperate Regions of Asia Including the Description of Three New Speces, Some Potentially Invasive to North America; U.S. Department of Agriculture, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2007. [Google Scholar]
- Luque, G.M.; Bellard, C.; Bertelsmeier, C.; Bonnaud, E.; Genovesi, P.; Simberloff, D.; Courchamp, F. The 100th of the world’s worst invasive alien species. Biol. Invasions 2014, 16, 981–985. [Google Scholar] [CrossRef]
- GISD. 100 of the World’s Worst Invasive Alien Species. Available online: http://www.iucngisd.org/gisd/100_worst.php (accessed on 27 April 2011).
- Feng, W.J. Risk analysis and control measures of gypsy moth in Shanxi. Shanxi For. Sci. Technol. 2021, 50, 12–13+24. [Google Scholar]
- Xiao, G.R. Chinese Forest Insects; China Forestry Publishing House: Beijing, China, 1992. [Google Scholar]
- Wen, L.R.; Zhao, C.Z.; Zhang, H.Y.; Chen, C.T. Observation on Biological Characteristics of Gypsy Moth. Jilin For. Technol. 1989, 34-37+21. [Google Scholar]
- Turner, N. The Gypsy Moth Problem; Connecticut Agricultural Experiment Station: New Haven, CT, USA, 1963. [Google Scholar]
- USDA. Department of Agriculture. Available online: https://www.aphis.usda.gov/aphis/ourfocus/planthealth/plant-pest-and-disease-programs/pests-and-diseases/gypsy-moth/ct_gypsy_moth (accessed on 15 December 2021).
- Forbush, E.H.; Fernald, C.H. The Gypsy Moth: Porthetria dispar (Linn.). A Report of the Work of Destroying the Insect in the Commonwealth of Massachusetts, Together with an Account of Its History and Habits Both in Massachusetts and Europe; Wright & Potter Printing Company: Boston, MA, USA, 1896. [Google Scholar]
- Headlee, T.J. The Present Status of the Gypsy Moth in New Jersey. J. Econ. Entomol. 1921, 14, 172–178. [Google Scholar] [CrossRef]
- Montgomery, M.E.; Wallner, W.E. The gypsy moth. In Dynamics of Forest Insect Populations; Springer: Boston, MA, USA, 1988; pp. 353–375. [Google Scholar]
- Wallner, W. Invasive pests (‘biological pollutants’) and US forests: Whose problem, who pays? Eppo Bull. 1996, 26, 167–180. [Google Scholar] [CrossRef]
- Wei, J.; Luo, Y.Q.; Shi, J.; Wang, D.P.; Shen, S.W. Impact of temperature on postdiapause and diapause of the Asian gypsy moth, Lymantria dispar asiatica. J. Insect Sci. 2014, 14, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keena, M.A. Comparison of the hatch of Lymantria dispar (Lepidoptera: Lymantriidae) eggs from Russia and the United States after exposure to different temperatures and durations of low temperature. Ann. Entomol. Soc. Am. 1996, 89, 564–572. [Google Scholar] [CrossRef]
- CFIA. List of Pests Regulated by Canada. AGM (Lymantria albescens, Lymantria umbrosa, Lymantria postalba, Lymantria dispar japonica and Lymantria dispar asiatica)—Canadian Food Inspection Agency (Canada.Ca). Available online: https://inspection.canada.ca/plant-health/invasive-species/insects/spongy-moth/agm/eng/1330353359964/1330353499535 (accessed on 15 February 2022).
- Cao, J. Quarantine Regulations of Asian gypsy moth on Entry Ships. China Customs. 2021, 6, 44. [Google Scholar]
- Chen, F.; Luo, Y.; Keena, M.A.; Wu, Y.; Wu, P.; Shi, J. DNA barcoding of gypsy moths from China (Lepidoptera: Erebidae) reveals new haplotypes and divergence patterns within gypsy moth subspecies. J. Econ. Entomol. 2016, 109, 366–374. [Google Scholar] [CrossRef]
- Chen, F.; Shi, J.; Luo, Y.-q.; Sun, S.-y.; Pu, M. Genetic characterization of the gypsy moth from China (Lepidoptera, Lymantriidae) using inter simple sequence repeats markers. PLoS ONE 2013, 8, e73017. [Google Scholar] [CrossRef]
- Wu, Y.; Du, Q.; Qin, H.; Shi, J.; Wu, Z.; Shao, W. Rapid identification of the Asian gypsy moth and its related species based on mitochondrial DNA. Ecol. Evol. 2018, 8, 2320–2325. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Wu, Y.; Kurenshchikov, D.K.; Ilyinykh, A.V.; Shi, J. Underestimated mitochondrial diversity in gypsy moth Lymantria dispar from Asia. Agr. Forest Entomol. 2019, 21, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Kurenshchikov, D.; Yu, J.; Zou, Y.; Wang, Y.; Wang, Y.; Shi, J. Microsatellite and morphological analyses reveal unexpected diversity in Lymantria dispar in China. Forests 2019, 10, 1100. [Google Scholar] [CrossRef]
- Wang, Y.; Harrison, R.L.; Shi, J. Effects of Rearing Density on Developmental Traits of Two Different Biotypes of the Gypsy Moth, Lymantria dispar L., from China and the USA. Insects 2021, 12, 175. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Luo, Y.; Shi, J. The influence of geographic population, age, and mating status on the flight activity of the Asian gypsy moth Lymantria dispar (Lepidoptera: Erebidae) in China. Appl. Entomol. Zool. 2017, 52, 265–270. [Google Scholar] [CrossRef]
- Shi, J.; Chen, F.; Keena, M.A. Differences in Wing Morphometrics of Lymantria dispar (Lepidoptera: Erebidae) Between Populations That Vary in Female Flight Capability. Ann. Entomol. Soc. Am. 2015, 108, 528–535. [Google Scholar] [CrossRef] [Green Version]
- Ji, W.; Shi, J. Beijing Key Laboratory for Forest Pest Control and Sino-French Joint Laboratory for Invasive Forest Pests in Eurasia; Beijing Forestry University: Beijing, China, 2023; manuscript in preparation. [Google Scholar]
- Xu, Y.; Zhang, S.; Wang, H.; Wang, M.; Li, G. Mitochondrial gene sequence (COI) reveals the genetic structure and demographic history of Lymantria dispar (Lepidoptera: Erebidae: Lymantriinae) in and around China. Insects 2019, 10, 146. [Google Scholar] [CrossRef] [Green Version]
- Donald, S.; Zahiri, R.; Djoumad, A.; Freschi, L.; Lamarche, J.; Holden, D.; Cervantes, S.; Ojeda, D.I.; Potvin, A.; Nisole, A. A multi-species TaqMan PCR assay for the identification of Asian gypsy moths (Lymantria spp.) and other invasive lymantriines of biosecurity concern to North America. PLoS ONE 2016, 11, e0160878. [Google Scholar]
- Hosseini, R.; Keller, M.A.; Schmidt, O.; Framenau, V.W. Molecular identification of wolf spiders (Araneae: Lycosidae) by multiplex polymerase chain reaction. Biol. Control 2007, 40, 128–135. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, X.; Kong, L.; Li, Q. Molecular identification of dried shellfish products sold on the market using DNA barcoding. J. Ocean. Univ. China 2021, 20, 931–938. [Google Scholar] [CrossRef]
- Shi, F.; Ge, S.; Hou, Z.; Xu, Y.; Tao, J.; Wu, H.; Zong, S. Species-specific primers for rapid detection of Monochamus saltuarius, an effective vector of Bursaphelenchus xylophilus in China. J. Appl. Entomol. 2022, 146, 636–647. [Google Scholar] [CrossRef]
- Song, X.; Wang, J.; Wang, X. Species-specific COI primers for rapid identification of Bemisia tabaci Mediterranean (MED) species. J. Appl. Entomol. 2021, 145, 1029–1038. [Google Scholar] [CrossRef]
- Jiang, F.; Li, Z.; Deng, Y.; Wu, J.; Liu, R.; Buahom, N. Rapid diagnosis of the economically important fruit fly, Bactrocera correcta (Diptera: Tephritidae) based on a species-specific barcoding cytochrome oxidase I marker. Bull. Entomol. Res. 2013, 103, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Xiao-Feng, L.I.; Wan, F.H.; Zhang, G.F.; Zhang, J.L. Identification of Phenacoccus solenopsis Tinsley (Hemiptera:Pseudococcidae) with species-specific COI (SS-COI) primers. Acta Entomol. Sin. 2013, 56, 689–696. [Google Scholar]
- Zhongling, Z. Lepidoptera, Lymantriidae Fauna Sinica: Insecta; Science Press: Beijing, China, 2003; Volume 30. [Google Scholar]
- Folmer, O.; Black, M.; Wr, H.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial Cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Simon, C.; Buckley, T.R.; Frati, F.; Stewart, J.B.; Beckenbach, A.T. Incorporating molecular evolution into phylogenetic analysis, and a new compilation of conserved polymerase chain reaction primers for animal mitochondrial DNA. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 545–579. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Proceedings of the Nucleic Acids Symposium Series, London, UK; 1999; pp. 95–98. [Google Scholar]
- Landry, J.F. Taxonomic review of the leek moth genus Acrolepiopsis (Lepidoptera: Acrolepiidae) in North America. Can. Entomol. 2007, 139, 319–353. [Google Scholar] [CrossRef]
- Nice, C.; Shapiro, A. Molecular and morphological divergence in the butterfly genus Lycaeides (Lepidoptera: Lycaenidae) in North America: Evidence of recent speciation. J. Evol. Biol. 1999, 12, 936–950. [Google Scholar] [CrossRef] [Green Version]
- Gajardo, G.; Crespo, J.; Triantafyllidis, A.; Tzika, A.; Baxevanis, A.D.; Kappas, I.; Abatzopoulos, T.J. Species identification of Chilean Artemia populations based on mitochondrial DNA RFLP analysis. J. Biogeogr. 2004, 31, 547–555. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Goto, A.; Nishida, M. Mitochondrial DNA sequence divergence between two cryptic species of Lethenteron, with reference to an improved identification technique. J. Fish Biol. 2003, 62, 591–609. [Google Scholar] [CrossRef]
- Dinh, N.T. The dna barcoding data and genetic distance of leaf beetles (Coleoptera, Chrysomelidae) in Vietnam. Aca. J. Biol. 2020, 42, 25–31. [Google Scholar] [CrossRef]
- Hebert, P.D.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. London. Ser. B Biol. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [Green Version]
- Kress, W.J.; García-Robledo, C.; Uriarte, M.; Erickson, D.L. DNA barcodes for ecology, evolution, and conservation. Trends Ecol. Evol. 2015, 30, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Mir, R.A.; Bhat, K.A.; Rashid, G.; Ebinezer, L.B.; Masi, A.; Rakwal, R.; Shah, A.; Zargar, S.M. DNA barcoding: A way forward to obtain deep insights about the realistic diversity of living organisms. Nucleus 2021, 64, 157–165. [Google Scholar] [CrossRef]
- Costa, F.O.; DeWaard, J.R.; Boutillier, J.; Ratnasingham, S.; Dooh, R.T.; Hajibabaei, M.; Hebert, P.D. Biological identifications through DNA barcodes: The case of the Crustacea. Can. J. Fish Aquat. Sci. 2007, 64, 272–295. [Google Scholar] [CrossRef]
- Kang, T.H.; Lee, K.S.; Lee, H.S. DNA barcoding of the Korean Lymantria Hübner, 1819 (Lepidoptera: Erebidae: Lymantriinae) for quarantine inspection. J. Econ. Entomol. 2015, 108, 1596–1611. [Google Scholar] [CrossRef]
- Boykin, L.M.; Armstrong, K.; Kubatko, L.; Barro, P.D. DNA barcoding invasive insects: Database roadblocks. Invertebr. Syst. 2012, 26, 506–514. [Google Scholar] [CrossRef]
- Hu, Z.; Xu, Z.; Lu, Y. Analysis of Common Problems and Research Progress of Loop Mediated Isothermal Amplification. Chin. J. Virol. 2016, 32, 659–665. [Google Scholar]
- Djoumad, A.; Nisole, A.; Stewart, D.; Holden, D.; Zahiri, R.; Inoue, M.N.; Martemyanov, V.V.; Levesque, R.C.; Hamelin, R.C.; Cusson, M. Reassessment of the status of Lymantria albescens and Lymantria postalba (Lepidoptera: Erebidae: Lymantriinae) as distinct ‘Asian gypsy moth’ species, using both mitochondrial and nuclear sequence data. Syst. Entomol. 2020, 45, 493–504. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
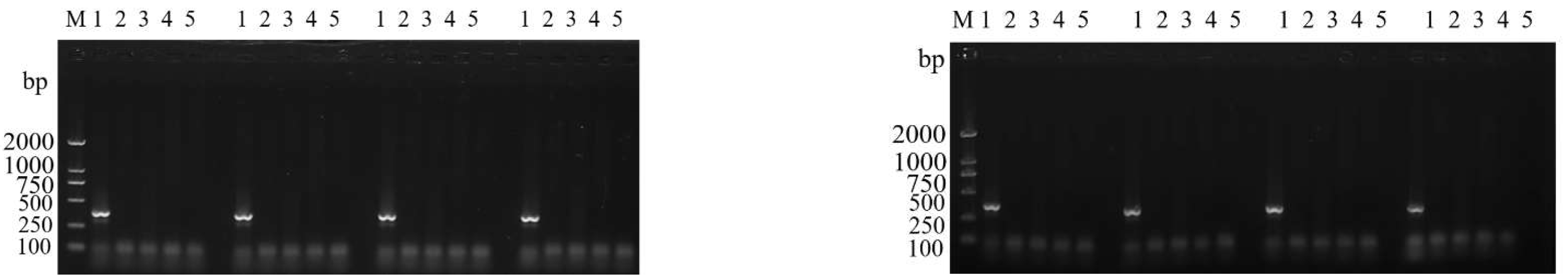{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species No. | Species | Location | Developmental Stage | Date |
|---|---|---|---|---|
| 1 | Lymantria dispar asiatica | Tian Jing | Adult and eggs | August, 2021 |
| 2 | Lymantria dispar asiatica | He Gang | Adult and eggs | August, 2021 |
| 3 | Lymantria dispar asiatica | Shan Dong | Adult and eggs | July, 2021 |
| 4 | Lymantria dispar asiatica | Chi Feng | Adult and eggs | July, 2021 |
| 5 | Lymantria dispar asiatica | Cheng De | Adult and eggs | July, 2021 |
| 6 | Lymantria dispar asiatica | He Bei | Adult and eggs | June, 2020 |
| 7 | Lymantria dispar asiatica | Cha Ri Su | Adult and eggs | September, 2020 |
| 8 | Lymantria dispar asiatica | Tong Liao | Adult and eggs | August, 2021 |
| 9 | Lymantria dispar asiatica | Bei Jing | Adult and eggs | August, 2020 |
| 10 | Lymantria dispar asiatica | Liao Ning | Adult and eggs | September, 2021 |
| 11 | Lymantria dispar asiatica | Shaan Xi | Adult and eggs | July, 2021 |
| 12 | Lymantria dispar asiatica | Shuo Zhou | Adult and eggs | August, 2021 |
| 13 | Lymantria dispar asiatica | Yun Cheng | Adult and eggs | July, 2021 |
| 14 | Lymantria dispar asiatica | Inner Mongolia | Adult | September , 2021 |
| 15 | Lymantria dispar asiatica | Gui zhou | Adult | June, 2020 |
| 16 | Lymantria dispar asiatica | Chengdu | Adult | July, 2020 |
| 17 | Lymantria dispar asiatica | Ulanhot | Adult | September, 2020 |
| 18 | Lymantria dispar japonica | Japan | Adult | September, 2017 |
| 19 | Lymantria monacha | Inner Mongolia | Adult and eggs | August, 2019 |
| 20 | Lymantria xylina | Fu Jian | Adult | August, 2019 |
| 21 | Lymantria apicebrunnea | Yun Nan | Adult and eggs | July, 2021 |
| 22 | Lymantria fumida | Gui Zhou | Adult | July, 2022 |
| 23 | Lymantria mathura | Si chuan | Adult | August, 2022 |
| 24 | Lymantria nebulosa | Yun Nan | Adult | August, 2022 |
| 25 | Lymantria marginata | Yun Nan | Adult | August, 2022 |
| 26 | Lymantria similis | Yun Nan | Adult | August, 2022 |
| No. | Species | Accession No. |
|---|---|---|
| 1 | Lymantria umbrosa | HM775854 |
| 2 | Lymantria dissoluta | HM775756 |
| 3 | Lymantria flavida | HM775761 |
| 4 | Lymantria mathura | HM775782 |
| 5 | Lymantria minomonis | HM775790 |
| 6 | Lymantria obfuscata | HM775826 |
| 7 | Lymantria schaeferi | HM775840 |
| 8 | Lymantria xylina | MW085568 |
| 9 | Lymantria plumbalis | HM775836 |
| 10 | Lymantria monacha | HM875343 |
| 11 | Lymantria lucescens | LC406220 |
| 12 | Lymantria postalba | LC406195 |
| 13 | Lymantria albescens | LC406184 |
| 14 | Lymantria antennata | HQ921458 |
| 15 | Lymantria atemeles | KP759547 |
| 16 | Lymantria bantaizana | KX436537 |
| 17 | Lymantria concolor | KX436392 |
| 18 | Lymantria dispar dispar | HM418030 |
| 19 | Lymantria dispar japonica | KY923061 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, W.; Dou, F.; Zhang, C.; Xiao, Y.; Yin, W.; Yu, J.; Kurenshchikov, D.K.; Zhu, X.; Shi, J. Improvement in the Identification Technology for Asian Spongy Moth, Lymantria dispar Linnaeus, 1758 (Lepidoptera: Erebidae) Based on SS-COI. Insects 2023, 14, 94. https://doi.org/10.3390/insects14010094
Ji W, Dou F, Zhang C, Xiao Y, Yin W, Yu J, Kurenshchikov DK, Zhu X, Shi J. Improvement in the Identification Technology for Asian Spongy Moth, Lymantria dispar Linnaeus, 1758 (Lepidoptera: Erebidae) Based on SS-COI. Insects. 2023; 14(1):94. https://doi.org/10.3390/insects14010094
Chicago/Turabian StyleJi, Wenzhuai, Fengrui Dou, Chunhua Zhang, Yuqian Xiao, Wenqi Yin, Jinyong Yu, D. K. Kurenshchikov, Xiue Zhu, and Juan Shi. 2023. "Improvement in the Identification Technology for Asian Spongy Moth, Lymantria dispar Linnaeus, 1758 (Lepidoptera: Erebidae) Based on SS-COI" Insects 14, no. 1: 94. https://doi.org/10.3390/insects14010094
APA StyleJi, W., Dou, F., Zhang, C., Xiao, Y., Yin, W., Yu, J., Kurenshchikov, D. K., Zhu, X., & Shi, J. (2023). Improvement in the Identification Technology for Asian Spongy Moth, Lymantria dispar Linnaeus, 1758 (Lepidoptera: Erebidae) Based on SS-COI. Insects, 14(1), 94. https://doi.org/10.3390/insects14010094