

Research Progress on the Regulation of Autophagy and Apoptosis in Insects by Sterol Hormone 20-Hydroxyecdysone


Abstract
:Simple Summary
Abstract
1. Introduction
2. Sterol Hormone 20E Biosynthesis
3. 20E Signal Transduction

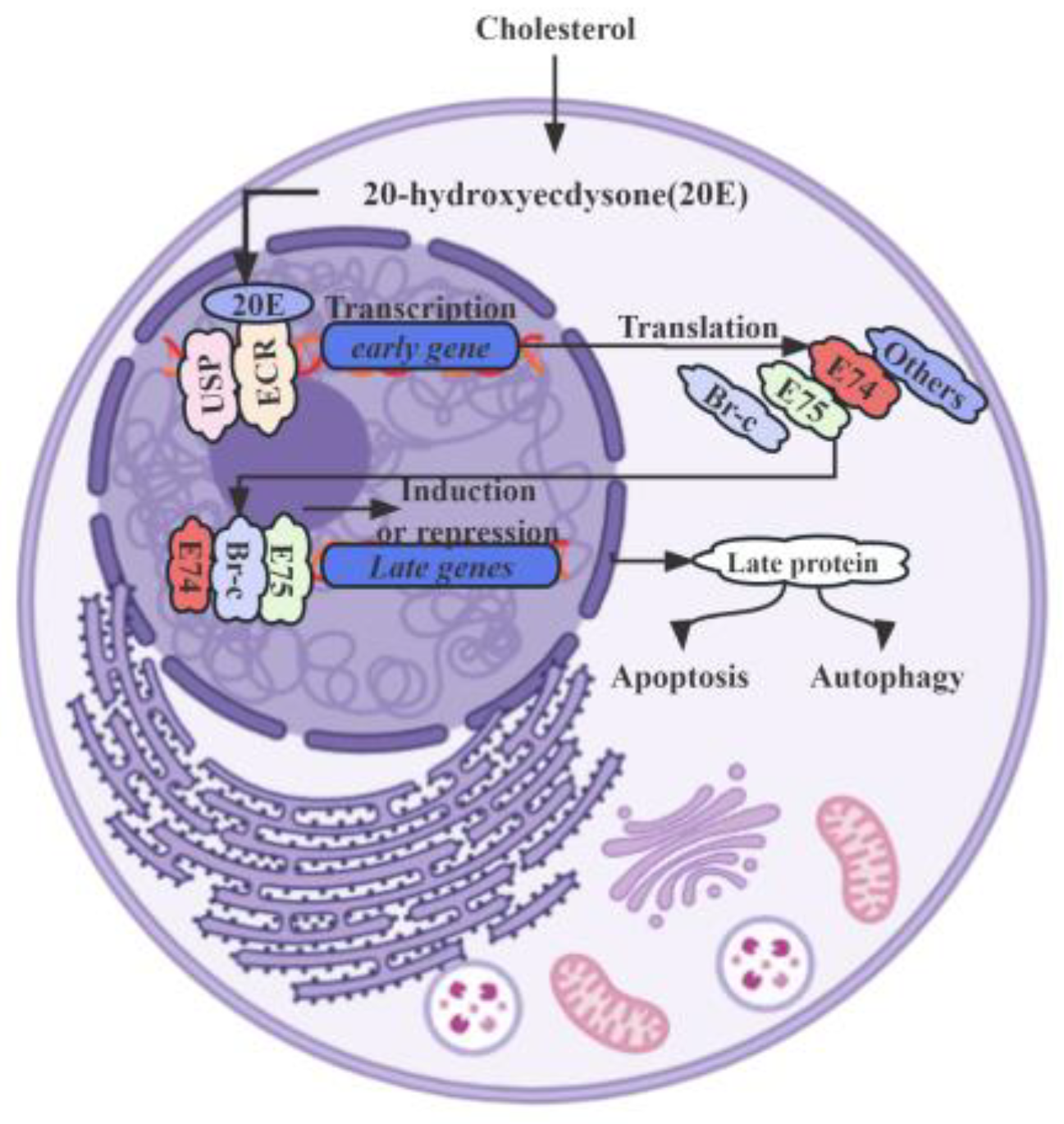{kind=link}
{kind=link}
| Species | Cytochrome P450 Enzymes | The Primary-Response Genes |
|---|---|---|
| D. melanogaster | DmCYP18a1 [41] DmCyp6a8/DmCyp6a2 [64] DmCyp6g1 [65] DmCYP302A1, DmCYP315A1 and DmCYP314A1 [66] | DmE75A [67] DmE75B, DmE75C and DmE75D [68] DmE93, DmE74, Br-C, and β-FTZ-F1 [69] |
| B. mori | BmCyp450, BmCYP314a1, BmCYP315a1, BmCYP302a1 and BmCYP306a1 [70] BmCYP4 and CYP6 [71] BmCYP18A1 [72] | BmE75A, BmE75B and BmE75C [73] BmE93 and BR-C [74] EcR-A and BHR3 [75] |
| H. armigera | HarmCYP302A1, HarmCYP306A1, HarmCYP314A1, and HarmCYP315A1 [53] HarmCYP6b2, HarmCYP18a1 and HarmCYP18b1 [76] | Br-C, E74, Ha-Ntf2, Ha-Ran, E75B, HHR3, and Ha-eIF5c [25] |
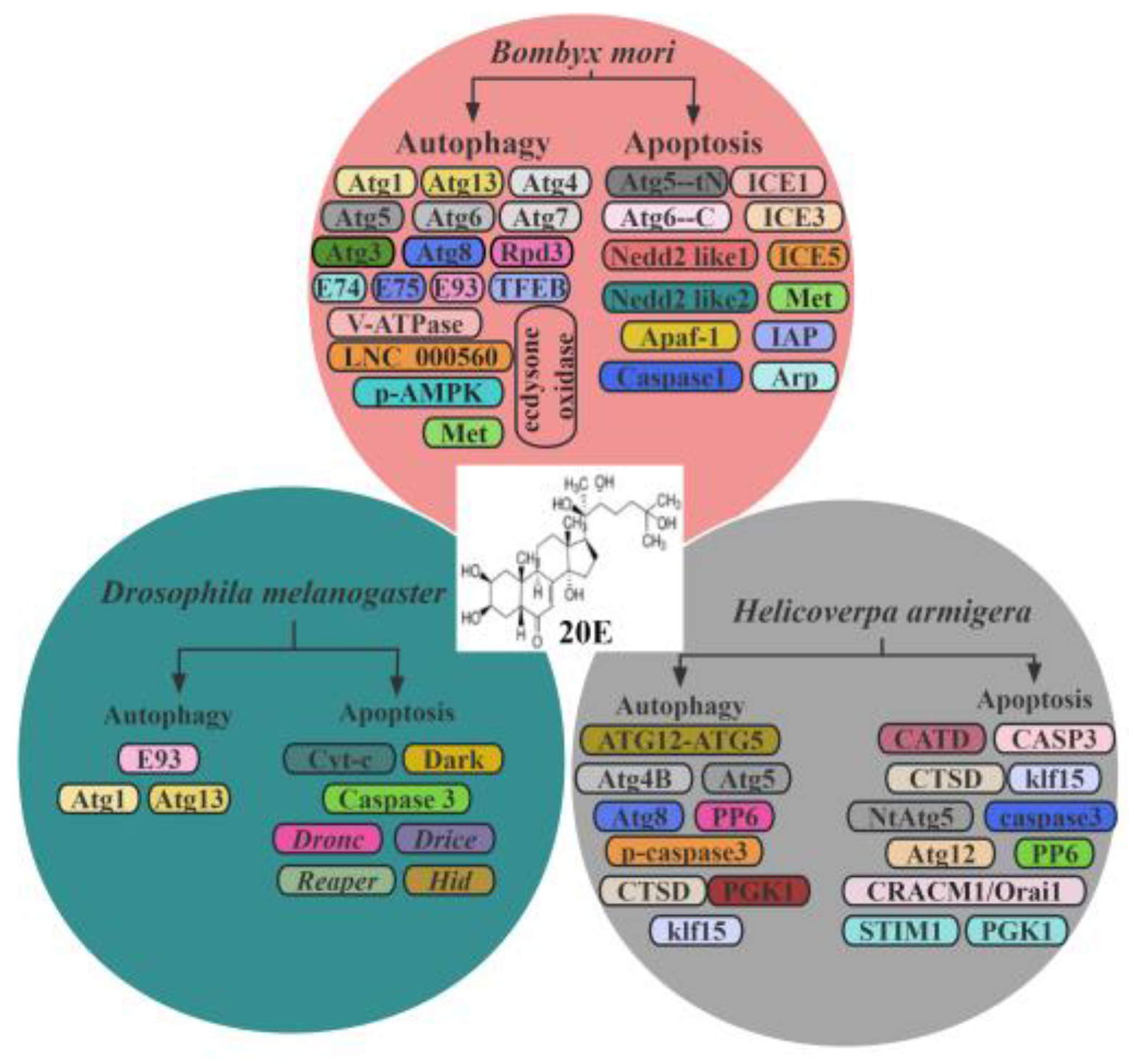
4. 20E Regulates Autophagy and Apoptosis in Insects
4.1. Regulation in D. melanogaster
4.2. Regulation in B. mori
4.3. Regulation in H. armigera
4.4. Regulation in the Others
| PCD-Related Enzymes | Species | Autophagy/Apoptosis | Up/Downregulation | Pathway | Associated Protein/Factors |
|---|---|---|---|---|---|
| Caspase | D. melanogaster | Apoptosis | Up [54] | PI3K-TORC1 | Caspase-9, Caspase 3, Caspase-6 |
| AMPK-PP2A axis | D. melanogaster | Autophagy | Down [15] | Insulin/IGF | PP2A, AKT |
| ATG1/ATG13 complex | D. melanogaster | Autophagy | Down [90] | PI3K/mTOR | PTEN |
| BmADK | B. mori | Autophagy and apoptosis | Down [61] | \ | ATG-8, Caspase-9, Ec-R, E74A, Br-C |
| BmV-ATPase | B. mori | Autophagy and apoptosis | Up [92] | mTOR | TFEB, BmATG8, BmATG5, BmATG6 |
| 20E-EcR-USP complex | B. mori | Autophagy and apoptosis | Down [94] | BECN1/ATG6-PIK3C3/Vps34 (catalytic subunit of class III PtdIns3K) | Br-C, E74, E75, E93 |
| BIGFLP | B. mori | Autophagy | Up [60] | Insulin/IGF | Insulin-like peptides |
| HDACs | B. mori | Autophagy | Up [98] | BmRpd3/HsHDAC1 | mTORC1 |
| HaP60 | H. armigera | Apoptosis | Up [62] | Insulin/IGF | ILPs, HaFOXO |
| PDK1 and FoxO | H. armigera | Apoptosis | Up [103] | \ | Insulin, calcium channel modulator 1 |
| CTSD/CathD/CATD | H. armigera | Apoptosis | Up [101] | Activation of MAPK1/ERK2 -MAPK3/ERK1 and AKT/protein kinase B | Ccaspase 3 and Caspase 7 |
| PGK1 | H. armigera | Autophagy | Up [80] | \ | ARD1, PTEN |
| PKCδ | H. armigera | Apoptosis | Up [105] | EcRB1/USP1 | Brz-7, Caspase-3, Caspase-6 |
5. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| 20E | 20-Hydroxyecdysone |
| PCD | Programmed cell death |
| E | Ecdysone |
| D. melanogaster | Drosophila melanogaster |
| B. mori | Bombyx mori |
| H. Armigera | Helicoverpa armigera |
| B. germanica | Blattella germanica |
| EnR | ecdysone receptor |
| JH | juvenile hormone |
| Hemimetabola | Dictyoptera to Hemiptera |
| USP | ultraspiracle protein |
| EcR-B1 | ecdysone receptor B1 isoform |
| USP1 | ubiquitin specific peptidase 1 |
| PTTH | Prothoracicotropic Hormone |
| Insulin/IIS | Insulin-like growth factor receptor signaling |
| IAP | inhibitor of apoptosis |
| AMPK | AMP-activated protein kinase |
| PP2A | protein phosphatase 2A |
| AKT | protein kinase B |
| Br-C | Broad-Complex |
| E74 | Ecdysone-induced protein 74 |
| E75 | Ecdysone-induced protein 75 |
| E93 | Ecdysone-induced protein 93 |
| BIGFLP | Bombyx imp-L2 gene-enhancer flippase protein |
| HaP60 | H. armigera P60 |
| Klf15 | Krüppel-like factor |
| PTEN | phosphatase and tensin homolog |
| Cyt-c | cytochrome C |
| lncRNAs | Long non-coding RNAs |
| Hid | head involution defective |
| Fkh | Fork head |
| CTSD | cathepsin D |
| FoxO | Forkhead Box O |
| Atgs | autophagy related genes |
| PE | phosphatidylethanolamine |
| α-KG | alpha-ketoglutarate |
| Glu | glutamate |
| STIM1 | stromal interacting molecule 1 |
| PKC | protein kinase C |
| GDH | glutamate dehydrogenase |
| PDK1 | phosphoinositide-dependent kinase-1 |
References
- Kaplanis, J.N.; Thompson, M.J.; Robbins, W.E.; Bryce, B.M. Insect hormones: Alpha ecdysone and 20-hydroxyecdysone in bracken fern. Science 1967, 157, 1436–1438. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Tian, L.; Guo, X.; Li, K.; Li, J.; Deng, X.; Li, Q.; Xia, Q.; Zhong, Y.; Huang, Z.; et al. BmATG5 and BmATG6 mediate apoptosis following autophagy induced by 20-hydroxyecdysone or starvation. Autophagy 2016, 12, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Werling, K.; Shaw, W.R.; Itoe, M.A.; Westervelt, K.A.; Marcenac, P.; Paton, D.G.; Peng, D.; Singh, N.; Smidler, A.L.; South, A.; et al. Steroid Hormone Function Controls Non-competitive Plasmodium Development in Anopheles. Cell 2019, 177, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Chen, Z.; Wen, J.; Jia, Q. Sterol Regulation of Development and 20-Hydroxyecdysone Biosynthetic and Signaling Genes in Drosophila melanogaster. Cells 2023, 12, 1739. [Google Scholar] [CrossRef] [PubMed]
- Sonobe, H.; Yamada, R. Ecdysteroids during early embryonic development in silkworm Bombyx mori: Metabolism and functions. Zool. Sci. 2004, 21, 503–516. [Google Scholar] [CrossRef]
- Wang, X.P.; Sun, S.P.; Li, Y.X.; Wang, L.; Dong, D.J.; Wang, J.X.; Zhao, X.F. 20-hydroxyecdysone reprograms amino acid metabolism to support the metamorphic development of Helicoverpa armigera. Cell Rep. 2023, 42, 112644. [Google Scholar] [CrossRef]
- Denton, D.; Aung-Htut, M.T.; Kumar, S. Developmentally programmed cell death in Drosophila. Biochim. Biophys. Acta 2013, 1833, 3499–3506. [Google Scholar] [CrossRef]
- He, H.J.; Hou, L.; Wang, J.X.; Zhao, X.F. The apoptosis inhibitor survivin prevents insect midgut from cell death during postembryonic development. Mol. Biol. Rep. 2012, 39, 1691–1699. [Google Scholar] [CrossRef] [PubMed]
- Tower, J. Programmed cell death in aging. Ageing Res. Rev. 2015, 23, 90–100. [Google Scholar] [CrossRef]
- Yamamoto, H.; Matsui, T. Molecular mechanisms of macroautophagy, microautophagy, and chaperone-mediated autophagy. J. Nippon Med. Sch. 2023. [Google Scholar] [CrossRef] [PubMed]
- Sorice, M. Crosstalk of Autophagy and Apoptosis. Cells 2022, 11, 1479. [Google Scholar] [CrossRef]
- Hsu, H.W.; Chiu, M.C.; Shih, C.J.; Matsuura, K.; Yang, C.S. Apoptosis as a primary defense mechanism in response to viral infection in invasive fire ant Solenopsis invicta. Virology 2019, 531, 255–259. [Google Scholar] [CrossRef]
- Wang, L.; Xiao, Q.; Zhou, X.L.; Zhu, Y.; Dong, Z.Q.; Chen, P.; Pan, M.H.; Lu, C. Bombyx mori Nuclear Polyhedrosis Virus (BmNPV) Induces Host Cell Autophagy to Benefit Infection. Viruses 2017, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Rost-Roszkowska, M.M.; Poprawa, I.; Chachulska-Zymełka, A. Apoptosis and autophagy in the midgut epithelium of Acheta domesticus (Insecta, Orthoptera, Gryllidae). Zool. Sci. 2010, 27, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Zhou, S.; Liu, S.; Li, K.; Zhao, H.; Long, S.; Liu, H.; Xie, Y.; Su, Y.; Yu, F.; et al. The AMPK-PP2A axis in insect fat body is activated by 20-hydroxyecdysone to antagonize insulin/IGF signaling and restrict growth rate. Proc. Natl. Acad. Sci. USA 2020, 117, 9292–9301. [Google Scholar] [CrossRef]
- Taylor, D.M.; Olds, C.L.; Haney, R.S.; Torrevillas, B.K.; Luckhart, S. Comprehensive and Durable Modulation of Growth, Development, Lifespan and Fecundity in Anopheles stephensi Following Larval Treatment with the Stress Signaling Molecule and Novel Antimalarial Abscisic Acid. Front. Microbiol. 2019, 10, 3024. [Google Scholar] [CrossRef]
- Li, Y.B.; Li, X.R.; Yang, T.; Wang, J.X.; Zhao, X.F. The steroid hormone 20-hydroxyecdysone promotes switching from autophagy to apoptosis by increasing intracellular calcium levels. Insect Biochem. Mol. Biol. 2016, 79, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Wu, X.; Zhou, L.; He, T.; Yin, Q.; Liu, S. 20-Hydroxyecdysone-responsive microRNAs of insects. RNA Biol. 2020, 17, 1454–1471. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Saha, T.T.; Zou, Z.; Raikhel, A.S. Regulatory Pathways Controlling Female Insect Reproduction. Annu. Rev. Entomol. 2018, 63, 489–511. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Fu, B.; Wang, C.; Liu, J.; Tang, Y.; Zhang, W.; Zhu, J.; Li, Y.; Pan, Q.; Liu, F. The role of 20E biosynthesis relative gene Shadow in the reproduction of the predatory mirid bug, Cyrtorhinus lividipennis (Hemiptera: Miridae). Arch. Insect Biochem. Physiol. 2022, 109, e21854. [Google Scholar] [CrossRef] [PubMed]
- Yoo, B.; Kim, H.Y.; Chen, X.; Shen, W.; Jang, J.S.; Stein, S.N.; Cormier, O.; Pereira, L.; Shih, C.; Krieger, C.; et al. 20-hydroxyecdysone (20E) signaling regulates amnioserosa morphogenesis during Drosophila dorsal closure: EcR modulates gene expression in a complex with the AP-1 subunit, Jun. Biol. Open 2021, 10, bio058605. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Sun, X.; Jin, X.Y. 20E-regulated USP expression and phosphorylation in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2003, 33, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.; Shimizu, S.; Fujiwara, H.; Sakurai, S.; Iwami, M. EcR expression in the prothoracicotropic hormone-producing neurosecretory cells of the Bombyx mori brain. FEBS J. 2006, 273, 3861–3868. [Google Scholar] [CrossRef] [PubMed]
- Lan, Q.; Hiruma, K.; Hu, X.; Jindra, M.; Riddiford, L.M. Activation of a delayed-early gene encoding MHR3 by the ecdysone receptor heterodimer EcR-B1-USP-1 but not by EcR-B1-USP-2. Mol. Cell. Biol. 1999, 19, 4897–4906. [Google Scholar] [CrossRef] [PubMed]
- He, H.J.; Wang, Q.; Zheng, W.W.; Wang, J.X.; Song, Q.S.; Zhao, X.F. Function of nuclear transport factor 2 and Ran in the 20E signal transduction pathway in the cotton bollworm, Helicoverpa armigera. BMC Mol. Cell Biol. 2010, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Hiruma, K.; Jindra, M.; Shinoda, T.; Segraves, W.A.; Malone, F.; Riddiford, L.M. Regulation of the transcription factor E75 by 20-hydroxyecdysone and juvenile hormone in the epidermis of the tobacco hornworm, Manduca sexta, during larval molting and metamorphosis. Dev. Biol. 1998, 193, 127–138. [Google Scholar] [CrossRef]
- Dong, D.J.; Wang, J.X.; Zhao, X.F. A eukaryotic initiation factor 5C is upregulated during metamorphosis in the cotton bollworm, Helicoverpa armigera. BMC Dev. Biol. 2009, 9, 19. [Google Scholar] [CrossRef]
- Kefi, M.; Balabanidou, V.; Douris, V.; Lycett, G.; Feyereisen, R.; Vontas, J. Two functionally distinct CYP4G genes of Anopheles gambiae contribute to cuticular hydrocarbon biosynthesis. Insect Biochem. Mol. Biol. 2019, 110, 52–59. [Google Scholar] [CrossRef]
- Cruz, J.; Martín, D.; Franch-Marro, X. Egfr Signaling Is a Major Regulator of Ecdysone Biosynthesis in the Drosophila Prothoracic Gland. Curr. Biol. 2020, 30, 1547–1554. [Google Scholar] [CrossRef]
- Engelmann, F. 20-hydroxyecdysone, what it can do. Science 1971, 174, 1041. [Google Scholar] [CrossRef]
- Temirgaziyev, B.S.; Kučáková, K.; Baizhigit, Y.A.; Jurášek, M.; Džubák, P.; Hajdúch, M.; Dolenský, B.; Drašar, P.B.; Tuleuov, B.I.; Adekenov, S.M. Bioavailability and structural study of 20-hydroxyecdysone complexes with cyclodextrins. Steroids 2019, 147, 37–41. [Google Scholar] [CrossRef]
- Horike, N.; Sonobe, H. Ecdysone 20-monooxygenase in eggs of the silkworm, Bombyx mori: Enzymatic properties and developmental changes. Arch. Insect Biochem. Physiol. 1999, 41, 9–17. [Google Scholar] [CrossRef]
- Rewitz, K.F.; Yamanaka, N.; Gilbert, L.I.; O’Connor, M.B. The insect neuropeptide PTTH activates receptor tyrosine kinase torso to initiate metamorphosis. Science 2009, 326, 1403–1405. [Google Scholar] [CrossRef] [PubMed]
- Ou, Q.; King-Jones, K. What goes up must come down: Transcription factors have their say in making ecdysone pulses. Curr. Top. Dev. Biol. 2013, 103, 35–71. [Google Scholar]
- Erion, R.; Sehgal, A. Regulation of insect behavior via the insulin-signaling pathway. Front. Physiol. 2013, 4, 353. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, G.; Yu, L.; Du, S.; Jiang, D.; Chu, X.; Wang, K.; Wu, S.; Wang, R.; Zhang, F.; et al. Temperature and metal ions regulate larval diapause termination via the 20-hydroxyecdysone and juvenile hormone pathways in Monochamus alternatus. Pest Manag. Sci. 2023, 79, 437–446. [Google Scholar] [CrossRef]
- Guo, S.; Tian, Z.; Wu, Q.W.; King-Jones, K.; Liu, W.; Zhu, F.; Wang, X.P. Steroid hormone ecdysone deficiency stimulates preparation for photoperiodic reproductive diapause. PLoS Genet. 2021, 17, e1009352. [Google Scholar] [CrossRef]
- Rauschenbach, I.Y.; Chentsova, N.A.; Alekseev, A.A.; Gruntenko, N.E.; Adonyeva, N.V.; Karpova, E.K.; Komarova, T.N.; Vasiliev, V.G.; Bownes, M. Dopamine and octopamine regulate 20-hydroxyecdysone level in vivo in Drosophila. Arch. Insect Biochem. Physiol. 2007, 65, 95–102. [Google Scholar] [CrossRef]
- Satake, S.; Masumura, M.; Ishizaki, H.; Nagata, K.; Kataoka, H.; Suzuki, A.; Mizoguchi, A. Bombyxin, an insulin-related peptide of insects, reduces the major storage carbohydrates in the silkworm Bombyx mori. Comp. Biochem. Phys. B 1997, 118, 349–357. [Google Scholar] [CrossRef]
- Iga, M.; Kataoka, H. Recent studies on insect hormone metabolic pathways mediated by cytochrome P450 enzymes. Biol. Pharm. Bull. 2012, 35, 838–843. [Google Scholar] [CrossRef]
- Guittard, E.; Blais, C.; Maria, A.; Parvy, J.P.; Pasricha, S.; Lumb, C.; Lafont, R.; Daborn, P.J.; Dauphin-Villemant, C. CYP18A1, a key enzyme of Drosophila steroid hormone inactivation, is essential for metamorphosis. Dev. Biol. 2011, 349, 35–45. [Google Scholar] [CrossRef]
- Shimell, M.; O’Connor, M.B. The cytochrome P450 Cyp6t3 is not required for ecdysone biosynthesis in Drosophila melanogaster. MicroPubl. Biol. 2022, 2022. [Google Scholar] [CrossRef]
- Rewitz, K.F.; Gilbert, L.I. Daphnia Halloween genes that encode cytochrome P450s mediating the synthesis of the arthropod molting hormone: Evolutionary implications. BMC Evol. Biol. 2008, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, S.; Jia, Q.; Wu, L.; Yuan, D.; Li, E.Y.; Feng, Q.; Wang, G.; Palli, S.R.; Wang, J.; et al. Juvenile hormone membrane signaling phosphorylates USP and thus potentiates 20-hydroxyecdysone action in Drosophila. Sci. Bull. 2022, 67, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Michalik, J.; Chojnicka, B.; Cymborowski, B. Vitellogenesis in virgin and mated females of the mealworm beetle, Tenebrio molitor. Acta Biochim. Pol. 1996, 43, 623–631. [Google Scholar] [CrossRef]
- Panaitof, S.C.; Scott, M.P. Effect of juvenile hormone on vitellogenin gene expression in the fat body of burying beetles, Nicrophorus orbicollis. Arch. Insect Biochem. Physiol. 2006, 63, 82–91. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; He, Q.; Zhou, S. Regulatory Mechanisms of Vitellogenesis in Insects. Front. Cell Dev. Biol. 2020, 8, 593613. [Google Scholar] [CrossRef]
- Zhu, L.; Yin, T.Y.; Sun, D.; Liu, W.; Zhu, F.; Lei, C.L.; Wang, X.P. Juvenile hormone regulates the differential expression of putative juvenile hormone esterases via methoprene-tolerant in non-diapause-destined and diapause-destined adult female beetle. Gene 2017, 627, 373–378. [Google Scholar] [CrossRef]
- Wyatt, G.R.; Davey, K.G. Cellular and Molecular Actions of Juvenile Hormone. II. Roles of Juvenile Hormone in Adult Insects. In Advances in Insect Physiology; Elsevier: Amsterdam, The Netherlands, 1996; Volume 26, pp. 1–155. [Google Scholar]
- Mpakou, V.E.; Velentzas, A.D.; Velentzas, P.D.; Margaritis, L.H.; Stravopodis, D.J.; Papassideri, I.S. Programmed cell death of the ovarian nurse cells during oogenesis of the ladybird beetle Adalia bipunctata (Coleoptera: Coccinellidae). Dev. Growth Differ. 2011, 53, 804–815. [Google Scholar] [CrossRef]
- Liu, S.; Li, K.; Gao, Y.; Liu, X.; Chen, W.; Ge, W.; Feng, Q.; Palli, S.R.; Li, S. Antagonistic actions of juvenile hormone and 20-hydroxyecdysone within the ring gland determine developmental transitions in Drosophila. Proc. Natl. Acad. Sci. USA 2018, 115, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Petryk, A.; Warren, J.T.; Marqués, G.; Jarcho, M.P.; Gilbert, L.I.; Kahler, J.; Parvy, J.P.; Li, Y.; Dauphin-Villemant, C.; O’Connor, M.B. Shade is the Drosophila P450 enzyme that mediates the hydroxylation of ecdysone to the steroid insect molting hormone 20-hydroxyecdysone. Proc. Natl. Acad. Sci. USA 2003, 100, 13773–13778. [Google Scholar] [CrossRef]
- Zheng, J.; Tian, K.; Yuan, Y.; Li, M.; Qiu, X. Identification and expression patterns of Halloween genes encoding cytochrome P450s involved in ecdysteroid biosynthesis in the cotton bollworm Helicoverpa armigera. B Entomol. Res. 2017, 107, 85–95. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Li, S. E93 predominantly transduces 20-hydroxyecdysone signaling to induce autophagy and caspase activity in Drosophila fat body. Insect Biochem. Mol. Biol. 2014, 45, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Tian, L.; Guo, Z.; Guo, S.; Zhang, J.; Gu, S.H.; Palli, S.R.; Cao, Y.; Li, S. 20-Hydroxyecdysone (20E) Primary Response Gene E75 Isoforms Mediate Steroidogenesis Autoregulation and Regulate Developmental Timing in Bombyx. J. Biol. Chem. 2016, 291, 18163–18175. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Dai, F.; Guo, E.; Li, K.; Ma, L.; Tian, L.; Cao, Y.; Zhang, G.; Palli, S.R.; Li, S. 20-Hydroxyecdysone (20E) Primary Response Gene E93 Modulates 20E Signaling to Promote Bombyx Larval-Pupal Metamorphosis. J Biol. Chem. 2015, 290, 27370–27383. [Google Scholar] [CrossRef] [PubMed]
- Tohidi-Esfahani, D.; Graham, L.D.; Hannan, G.N.; Simpson, A.M.; Hill, R.J. An ecdysone receptor from the pentatomomorphan, Nezara viridula, shows similar affinities for moulting hormones makisterone A and 20-hydroxyecdysone. Insect Biochem. Mol. Biol. 2011, 41, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jia, Q.; Tettamanti, G.; Li, S. Balancing crosstalk between 20-hydroxyecdysone-induced autophagy and caspase activity in the fat body during Drosophila larval-prepupal transition. Insect Biochem. Mol. Biol. 2013, 43, 1068–1078. [Google Scholar] [CrossRef]
- Tian, L.; Guo, E.; Wang, S.; Liu, S.; Jiang, R.J.; Cao, Y.; Ling, E.; Li, S. Developmental regulation of glycolysis by 20-hydroxyecdysone and juvenile hormone in fat body tissues of the silkworm, Bombyx mori. J. Mol. Cell Biol. 2010, 2, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Fujinaga, D.; Kohmura, Y.; Okamoto, N.; Kataoka, H.; Mizoguchi, A. Insulin-like growth factor (IGF)-like peptide and 20-hydroxyecdysone regulate the growth and development of the male genital disk through different mechanisms in the silkmoth, Bombyx mori. Insect Biochem. Mol. Biol. 2017, 87, 35–44. [Google Scholar] [CrossRef]
- Song, K.; Li, Y.; He, H.; Liu, L.; Zhao, P.; Xia, Q.; Wang, Y. A Novel Adenosine Kinase from Bombyx mori: Enzymatic Activity, Structure, and Biological Function. Int. J. Mol. Sci. 2019, 20, 3732. [Google Scholar] [CrossRef]
- Di, Y.Q.; Zhao, Y.M.; Jin, K.Y.; Zhao, X.F. Subunit P60 of phosphatidylinositol 3-kinase promotes cell proliferation or apoptosis depending on its phosphorylation status. PLoS Genet. 2021, 17, e1009514. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Xu, Q.Y.; Fu, K.Y.; Guo, W.C.; Li, G.Q. RNA interference against the putative insulin receptor substrate gene chico affects metamorphosis in Leptinotarsa decemlineata. Insect Biochem. Mol. Biol. 2018, 103, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bhaskara, S.; Chandrasekharan, M.B.; Ganguly, R. Caffeine induction of Cyp6a2 and Cyp6a8 genes of Drosophila melanogaster is modulated by cAMP and D-JUN protein levels. Gene 2008, 415, 49–59. [Google Scholar] [CrossRef]
- Jones, R.T.; Bakker, S.E.; Stone, D.; Shuttleworth, S.N.; Boundy, S.; McCart, C.; Daborn, P.J.; Ffrench-Constant, R.H.; van den Elsen, J.M. Homology modelling of Drosophila cytochrome P450 enzymes associated with insecticide resistance. Pest Manag. Sci. 2010, 66, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.I. Halloween genes encode P450 enzymes that mediate steroid hormone biosynthesis in Drosophila melanogaster. Mol. Cell. Endocrinol. 2004, 215, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Segraves, W.A.; Hogness, D.S. The E75 ecdysone-inducible gene responsible for the 75B early puff in Drosophila encodes two new members of the steroid receptor superfamily. Gene Dev. 1990, 4, 204–219. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, T.J.; Dubrovskaya, V.A.; Jannat, H.; Maughan, B.; Dubrovsky, E.B. Hormonal regulation of the E75 gene in Drosophila: Identifying functional regulatory elements through computational and biological analysis. J. Mol. Biol. 2009, 387, 794–808. [Google Scholar] [CrossRef]
- Rewitz, K.F.; Yamanaka, N.; O’Connor, M.B. Steroid hormone inactivation is required during the juvenile-adult transition in Drosophila. Dev. Cell 2010, 19, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mao, T.; Wang, H.; Lu, Z.; Qu, J.; Fang, Y.; Chen, J.; Li, M.; Cheng, X.; Hu, J.; et al. The CncC/keap1 pathway is activated in high temperature-induced metamorphosis and mediates the expression of Cyp450 genes in silkworm, Bombyx mori. Biochem. Biophys. Res. Commun. 2019, 514, 1045–1050. [Google Scholar] [CrossRef]
- Li, B.; Zhang, H.; Ni, M.; Wang, B.B.; Li, F.C.; Xu, K.Z.; Shen, W.D.; Xia, Q.Y.; Zhao, P. Identification and characterization of six cytochrome P450 genes belonging to CYP4 and CYP6 gene families in the silkworm, Bombyx mori. Mol. Biol. Rep. 2014, 41, 5135–5146. [Google Scholar] [CrossRef]
- Li, Z.; Ge, X.; Ling, L.; Zeng, B.; Xu, J.; Aslam, A.F.; You, L.; Palli, S.R.; Huang, Y.; Tan, A. CYP18A1 regulates tissue-specific steroid hormone inactivation in Bombyx mori. Insect Biochem. Mol. Biol. 2014, 54, 33–41. [Google Scholar] [CrossRef]
- Li, K.; Guo, E.; Hossain, M.S.; Li, Q.; Cao, Y.; Tian, L.; Deng, X.; Li, S. Bombyx E75 isoforms display stage- and tissue-specific responses to 20-hydroxyecdysone. Sci. Rep. 2015, 5, 12114. [Google Scholar] [CrossRef] [PubMed]
- Reza, A.M.; Kanamori, Y.; Shinoda, T.; Shimura, S.; Mita, K.; Nakahara, Y.; Kiuchi, M.; Kamimura, M. Hormonal control of a metamorphosis-specific transcriptional factor Broad-Complex in silkworm. Comp. Biochem. Phys. B 2004, 139, 753–761. [Google Scholar] [CrossRef]
- Sekimoto, T.; Iwami, M.; Sakurai, S. Coordinate responses of transcription factors to ecdysone during programmed cell death in the anterior silk gland of the silkworm, Bombyx mori. Insect Mol. Biol. 2006, 15, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Yang, X.; Bu, Z.; Li, X.; Zhang, Z.; Sun, W. Identification and Characterization of Genes Involved in Ecdysteroid Esterification Pathway Contributing to the High 20-Hydroxyecdysone Resistance of Helicoverpa armigera. Front. Physiol. 2020, 11, 508. [Google Scholar] [CrossRef] [PubMed]
- Winbush, A.; Weeks, J.C. Steroid-triggered, cell-autonomous death of a Drosophila motoneuron during metamorphosis. Neural Dev. 2011, 6, 15. [Google Scholar] [CrossRef]
- Li, Y.B.; Yang, T.; Wang, J.X.; Zhao, X.F. The Steroid Hormone 20-Hydroxyecdysone Regulates the Conjugation of Autophagy-Related Proteins 12 and 5 in a Concentration and Time-Dependent Manner to Promote Insect Midgut Programmed Cell Death. Front. Endocrinol. 2018, 9, 28. [Google Scholar] [CrossRef]
- Wang, X.P.; Huang, Z.; Li, Y.L.; Jin, K.Y.; Dong, D.J.; Wang, J.X.; Zhao, X.F. Krüppel-like factor 15 integrated autophagy and gluconeogenesis to maintain glucose homeostasis under 20-hydroxyecdysone regulation. PLoS Genet. 2022, 18, e1010229. [Google Scholar] [CrossRef]
- Kang, X.L.; Li, Y.X.; Dong, D.J.; Wang, J.X.; Zhao, X.F. 20-Hydroxyecdysone counteracts insulin to promote programmed cell death by modifying phosphoglycerate kinase 1. BMC Biol. 2023, 21, 119. [Google Scholar] [CrossRef]
- Tennessen, J.M. Ecdysone and 20-hydroxyecdysone are not required to activate glycolytic gene expression in Drosophila melanogaster embryos. MicroPubl. Biol. 2021, 2021. [Google Scholar] [CrossRef]
- Tettamanti, G.; Casartelli, M. Cell death during complete metamorphosis. Philos. Trans. R. Soc. B 2019, 374, 20190065. [Google Scholar] [CrossRef] [PubMed]
- Rost-Roszkowska, M.M. Degeneration of the midgut epithelium in Allacma fusca L. (Insecta, Collembola, Symphypleona): Apoptosis and necrosis. Zool. Sci. 2008, 25, 753–759. [Google Scholar] [CrossRef]
- Song, X.F.; Tian, H.; Zhang, P.; Zhang, Z.X. Expression of Cyt-c-Mediated Mitochondrial Apoptosis-Related Proteins in Rat Renal Proximal Tubules during Development. Nephron 2017, 135, 77–86. [Google Scholar] [CrossRef]
- Nicolson, S.; Denton, D.; Kumar, S. Ecdysone-mediated programmed cell death in Drosophila. Int. J. Dev. Biol. 2015, 59, 23–32. [Google Scholar] [CrossRef]
- Wang, J.; Wang, S.; Li, S. Sumoylation modulates 20-hydroxyecdysone signaling by maintaining USP protein levels in Drosophila. Insect Biochem. Mol. Biol. 2014, 54, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Long, S.; Cao, W.; Qiu, Y.; Deng, R.; Liu, J.; Zhang, L.; Dong, R.; Liu, F.; Li, S.; Zhao, H.; et al. The appearance of cytoplasmic cytochrome C precedes apoptosis during Drosophila salivary gland degradation. Insect Sci. 2023. [Google Scholar] [CrossRef] [PubMed]
- Draizen, T.A.; Ewer, J.; Robinow, S. Genetic and hormonal regulation of the death of peptidergic neurons in the Drosophila central nervous system. J. Neurobiol. 1999, 38, 455–465. [Google Scholar] [CrossRef]
- Cao, C.; Liu, Y.; Lehmann, M. Fork head controls the timing and tissue selectivity of steroid-induced developmental cell death. J. Cell Biol. 2007, 176, 843–852. [Google Scholar] [CrossRef]
- Rusten, T.E.; Lindmo, K.; Juhász, G.; Sass, M.; Seglen, P.O.; Brech, A.; Stenmark, H. Programmed autophagy in the Drosophila fat body is induced by ecdysone through regulation of the PI3K pathway. Dev. Cell 2004, 7, 179–192. [Google Scholar] [CrossRef]
- Yin, V.P.; Thummel, C.S. Mechanisms of steroid-triggered programmed cell death in Drosophila. Semin. Cell Dev. Biol. 2005, 16, 237–243. [Google Scholar] [CrossRef]
- Dai, Y.; Li, K.; Wu, W.; Wu, K.; Yi, H.; Li, W.; Xiao, Y.; Zhong, Y.; Cao, Y.; Tian, L. Steroid hormone 20-hydroxyecdysone induces the transcription and complex assembly of V-ATPases to facilitate autophagy in Bombyx mori. Insect Biochem. Mol. Biol. 2020, 116, 103255. [Google Scholar] [CrossRef]
- Tian, L.; Liu, S.; Liu, H.; Li, S. 20-hydroxyecdysone upregulates apoptotic genes and induces apoptosis in the Bombyx fat body. Arch. Insect Biochem. Physiol. 2012, 79, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Ma, L.; Guo, E.; Deng, X.; Ma, S.; Xia, Q.; Cao, Y.; Li, S. 20-Hydroxyecdysone upregulates Atg genes to induce autophagy in the Bombyx fat body. Autophagy 2013, 9, 1172–1187. [Google Scholar] [CrossRef]
- Wang, X.; Ding, Y.; Lu, X.; Geng, D.; Li, S.; Raikhel, A.S.; Zou, Z. The ecdysone-induced protein 93 is a key factor regulating gonadotrophic cycles in the adult female mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2021, 118, e2021910118. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, D.; Casartelli, M.; Cappellozza, S.; de Eguileor, M.; Tettamanti, G. Roles and regulation of autophagy and apoptosis in the remodelling of the lepidopteran midgut epithelium during metamorphosis. Sci. Rep. 2016, 6, 32939. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xu, J.; Xu, X.; Ma, W.; Tian, L.; Li, K. Functional identification of Bombyx mori Atg13 in autophagy. Arch. Insect Biochem. Physiol. 2020, 105, e21718. [Google Scholar] [CrossRef]
- Wu, W.; Luo, M.; Li, K.; Dai, Y.; Yi, H.; Zhong, Y.; Cao, Y.; Tettamanti, G.; Tian, L. Cholesterol derivatives induce dephosphorylation of the histone deacetylases Rpd3/HDAC1 to upregulate autophagy. Autophagy 2021, 17, 512–528. [Google Scholar] [CrossRef]
- Xu, X.; Cui, L.; Zhong, W.; Cai, Y. Autophagy-Associated lncRNAs: Promising Targets for Neurological Disease Diagnosis and Therapy. Neural Plast. 2020, 2020, 8881687. [Google Scholar] [CrossRef]
- Qiao, H.; Wang, J.; Wang, Y.; Yang, J.; Wei, B.; Li, M.; Wang, B.; Li, X.; Cao, Y.; Tian, L.; et al. Transcriptome analysis reveals potential function of long non-coding RNAs in 20-hydroxyecdysone regulated autophagy in Bombyx mori. BMC Genom. 2021, 22, 374. [Google Scholar] [CrossRef]
- Di, Y.Q.; Han, X.L.; Kang, X.L.; Wang, D.; Chen, C.H.; Wang, J.X.; Zhao, X.F. Autophagy triggers CTSD (cathepsin D) maturation and localization inside cells to promote apoptosis. Autophagy 2021, 17, 1170–1192. [Google Scholar] [CrossRef]
- Pan, J.; Di, Y.Q.; Li, Y.B.; Chen, C.H.; Wang, J.X.; Zhao, X.F. Insulin and 20-hydroxyecdysone oppose each other in the regulation of phosphoinositide-dependent kinase-1 expression during insect pupation. J. Biol. Chem. 2018, 293, 18613–18623. [Google Scholar] [CrossRef]
- Li, Y.B.; Pei, X.Y.; Wang, D.; Chen, C.H.; Cai, M.J.; Wang, J.X.; Zhao, X.F. The steroid hormone 20-hydroxyecdysone upregulates calcium release-activated calcium channel modulator 1 expression to induce apoptosis in the midgut of Helicoverpa armigera. Cell Calcium 2017, 68, 24–33. [Google Scholar] [CrossRef]
- Chen, C.H.; Di, Y.Q.; Shen, Q.Y.; Wang, J.X.; Zhao, X.F. The steroid hormone 20-hydroxyecdysone induces phosphorylation and aggregation of stromal interacting molecule 1 for store-operated calcium entry. J. Biol. Chem. 2019, 294, 14922–14936. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Pan, J.; Di, Y.Q.; Liu, W.; Hou, L.; Wang, J.X.; Zhao, X.F. Protein kinase C delta phosphorylates ecdysone receptor B1 to promote gene expression and apoptosis under 20-hydroxyecdysone regulation. Proc. Natl. Acad. Sci. USA 2017, 114, E7121–E7130. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Liu, W.; Zhao, W.L.; Wang, J.X.; Zhao, X.F. Upregulation of the expression of prodeath serine/threonine protein kinase for programmed cell death by steroid hormone 20-hydroxyecdysone. Apoptosis 2013, 18, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Mané-Padrós, D.; Cruz, J.; Vilaplana, L.; Nieva, C.; Ureña, E.; Bellés, X.; Martín, D. The hormonal pathway controlling cell death during metamorphosis in a hemimetabolous insect. Dev. Biol. 2010, 346, 150–160. [Google Scholar] [CrossRef]
- Yu, J.; Song, H.; Wang, Y.; Liu, Z.; Wang, H.; Xu, B. 20-hydroxyecdysone Upregulates Ecdysone Receptor (ECR) Gene to Promote Pupation in the Honeybee, Apis mellifera Ligustica. Integr. Comp. Biol. 2023, 63, 288–303. [Google Scholar] [CrossRef]
- Li, Z.; Lyu, Z.; Ye, Q.; Cheng, J.; Wang, C.; Lin, T. Cloning, Expression Analysis, 20-Hydroxyecdysone Induction, and RNA Interference Study of Autophagy-Related Gene 8 from Heortia vitessoides Moore. Insects 2020, 11, 245. [Google Scholar] [CrossRef]
- Poyraz, T.E.; Goncu, E.; Koc, K. Apoptotic and autophagic characteristics of perivisceral fat body remodeling of the greater wax moth Galleria mellonella and effects of juvenile hormone analog, fenoxycarb, on these processes. Arch. Insect Biochem. Physiol. 2021, 107, e21780. [Google Scholar]
- Ekoka, E.; Maharaj, S.; Nardini, L.; Dahan-Moss, Y.; Koekemoer, L.L. 20-Hydroxyecdysone (20E) signaling as a promising target for the chemical control of malaria vectors. Parasite Vector 2021, 14, 86. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, L.; Li, H.; Zheng, Q.; Hu, J.; Wu, W. Research Progress on the Regulation of Autophagy and Apoptosis in Insects by Sterol Hormone 20-Hydroxyecdysone. Insects 2023, 14, 871. https://doi.org/10.3390/insects14110871
Lin L, Li H, Zheng Q, Hu J, Wu W. Research Progress on the Regulation of Autophagy and Apoptosis in Insects by Sterol Hormone 20-Hydroxyecdysone. Insects. 2023; 14(11):871. https://doi.org/10.3390/insects14110871
Chicago/Turabian StyleLin, Luobin, Huaqin Li, Qinzhou Zheng, Jiaxuan Hu, and Wenmei Wu. 2023. "Research Progress on the Regulation of Autophagy and Apoptosis in Insects by Sterol Hormone 20-Hydroxyecdysone" Insects 14, no. 11: 871. https://doi.org/10.3390/insects14110871
APA StyleLin, L., Li, H., Zheng, Q., Hu, J., & Wu, W. (2023). Research Progress on the Regulation of Autophagy and Apoptosis in Insects by Sterol Hormone 20-Hydroxyecdysone. Insects, 14(11), 871. https://doi.org/10.3390/insects14110871