
Mechanisms of Neuroendocrine Stress Response in Drosophila and Its Effect on Carbohydrate and Lipid Metabolism



{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Insulin/Insulin-Like Growth Factors Signaling Pathway in Drosophila melanogaster
3. Stress-Related Hormones in Drosophila melanogaster
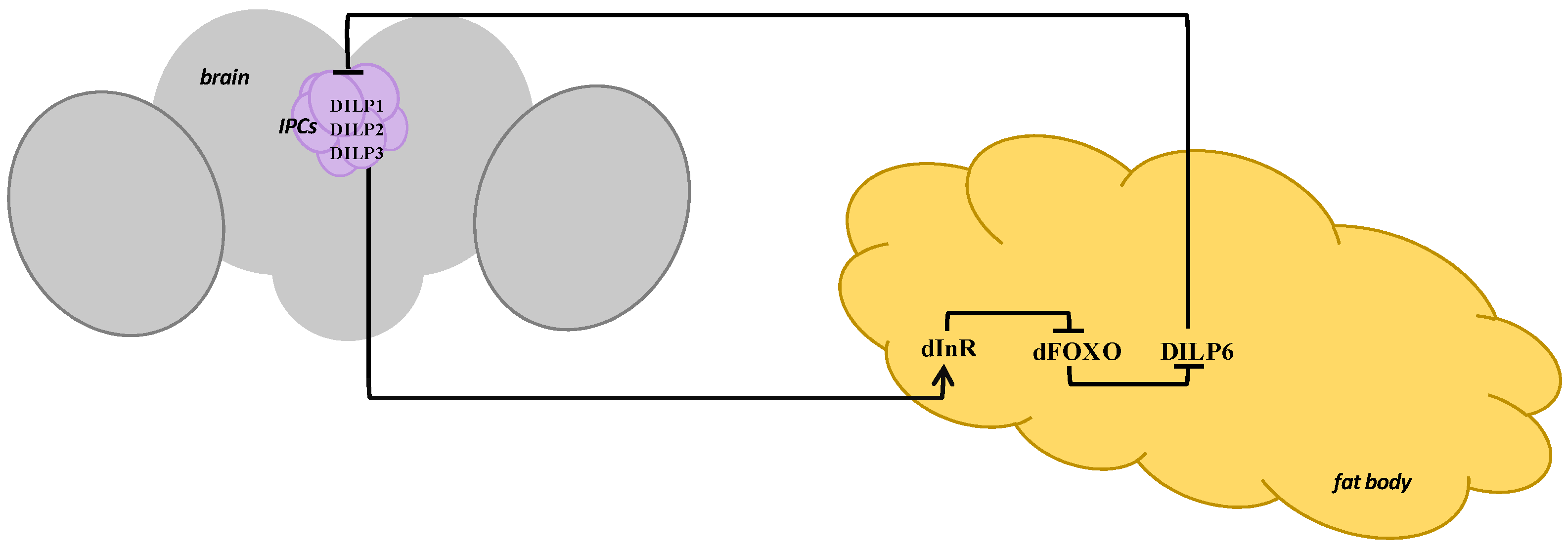
4. Drosophila IIS Pathway Regulation under Stress
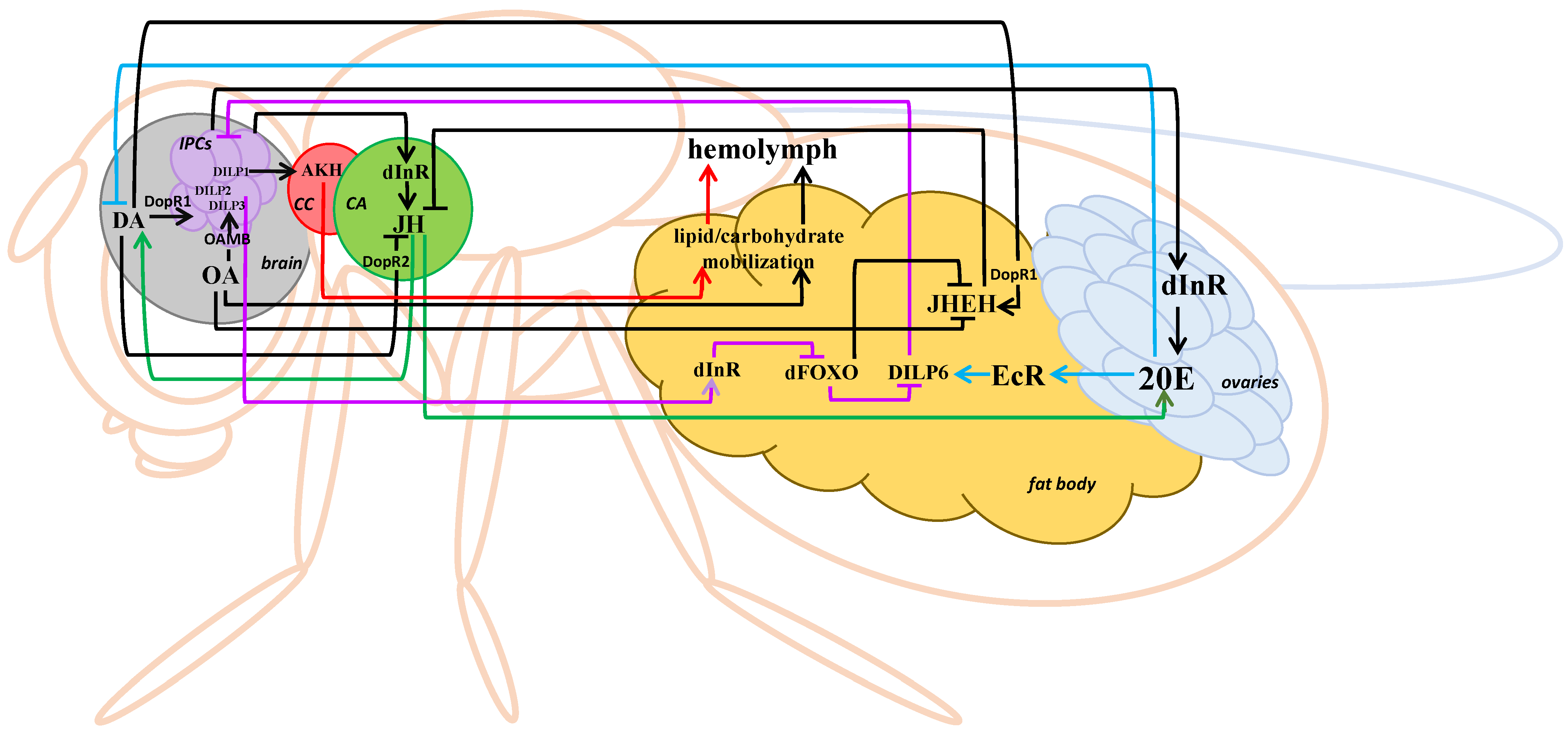
5. Interaction of the IIS Pathway and Stress-Related Hormones in Drosophila melanogaster
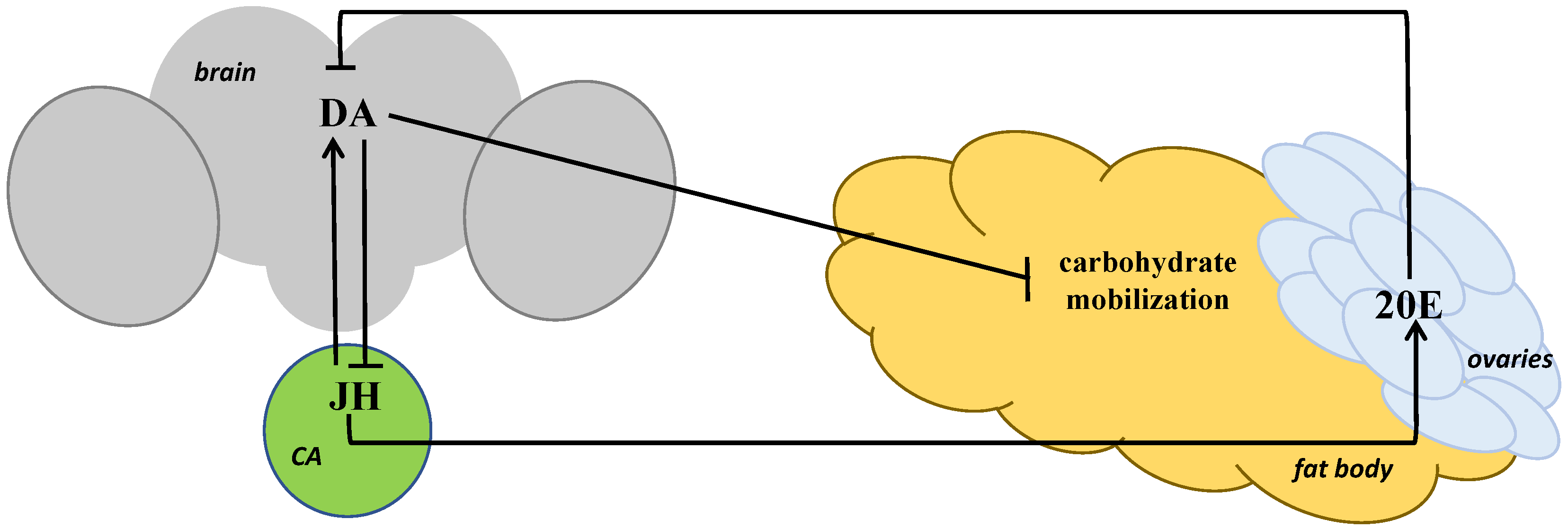
6. A Possible Mechanism of Short-Term Heat Stress Influence on Carbohydrate and Lipid Metabolism in Drosophila melanogaster
7. The Effect of Short-Term Heat Stress on Feeding Behavior in Drosophila melanogaster
8. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garofalo, R.S. 2 Genetic analysis of insulin signaling in Drosophila. Trends Endocrinol. Metab. 2002, 13, 156–162. [Google Scholar] [CrossRef]
- Chowański, S.; Walkowiak-Nowicka, K.; Winkiel, M.; Marciniak, P.; Urbański, A.; Pacholska-Bogalska, J. Insulin-like peptides and cross-talk with other factors in the regulation of insect metabolism. Front. Physiol. 2021, 12, 701203. [Google Scholar] [CrossRef] [PubMed]
- Brogiolo, W.; Stocker, H.; Ikeya, T.; Rintelen, F.; Fernandez, R.; Hafen, E. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr. Biol. 2001, 11, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Grönke, S.; Clarke, D.F.; Broughton, S.; Andrews, T.D.; Partridge, L. Molecular evolution and functional characterization of Drosophila insulin-like peptides. PLoS Genet. 2010, 6, e1000857. [Google Scholar] [CrossRef] [PubMed]
- Garelli, A.; Gontijo, A.M.; Miguela, V.; Caparros, E.; Dominguez, M. Imaginal discs secrete insulin-like peptide 8 to mediate plasticity of growth and maturation. Science 2012, 336, 579–582. [Google Scholar] [CrossRef]
- Colombani, J.; Andersen, D.S.; Leopold, P. Secreted peptide Dilp8 coordinates Drosophila tissue growth with developmental timing. Science 2012, 336, 582–585. [Google Scholar] [CrossRef]
- Álvarez-Rendón, J.P.; Salceda, R.; Riesgo-Escovar, J.R. Drosophila melanogaster as a model for diabetes type 2 progression. BioMed Res. Int. 2018, 2018, 1417528. [Google Scholar] [CrossRef]
- Miguel-Aliaga, I.; Thor, S.; Gould, A.P. Postmitotic specification of Drosophila insulinergic neurons from pioneer neurons. PLoS Biol. 2008, 6, e58. [Google Scholar] [CrossRef]
- Bai, H.; Kang, P.; Tatar, M. Drosophila insulin-like peptide-6 (dilp6) expression from fat body extends lifespan and represses secretion of Drosophila insulin-like peptide-2 from the brain. Aging Cell 2012, 11, 978–985. [Google Scholar] [CrossRef]
- Post, S.; Karashchuk, G.; Wade, J.D.; Sajid, W.; De Meyts, P.; Tatar, M. Drosophila insulin-like peptides DILP2 and DILP5 differentially stimulate cell signaling and glycogen phosphorylase to regulate longevity. Front. Endocrinol. 2018, 9, 245. [Google Scholar] [CrossRef]
- Post, S.; Liao, S.; Yamamoto, R.; Veenstra, J.A.; Nässel, D.R.; Tatar, M. Drosophila insulin-like peptide dilp1 increases lifespan and glucagon-like Akh expression epistatic to dilp2. Aging Cell 2019, 18, 12863. [Google Scholar] [CrossRef] [PubMed]
- Prince, E.; Kretzschmar, J.; Trautenberg, L.C.; Broschk, S.; Brankatschk, M. DIlp7-producing neurons regulate insulin-producing cells in Drosophila. Front Physiol. 2021, 12, 630390. [Google Scholar] [CrossRef] [PubMed]
- Rulifson, E.J.; Kim, S.K.; Nusse, R. Ablation of insulinproducing neurons in flies: Growth and diabetic phenotypes. Science 2002, 296, 1118–1120. [Google Scholar] [CrossRef] [PubMed]
- Géminard, C.; Rulifson, E.J.; Léopold, P. Remote control of insulin secretion by fat cells in Drosophila. Cell Metab. 2009, 10, 199–207. [Google Scholar] [CrossRef]
- Cognigni, P.; Bailey, A.P.; Miguel-Aliaga, I. Enteric neurons and systemic signals couple nutritional and reproductive status with intestinal homeostasis. Cell Metab. 2011, 13, 92–104. [Google Scholar] [CrossRef]
- Nässel, D.R.; Kubrak, O.I.; Liu, Y.; Luo, J.; Lushchak, O.V. Factors that regulate insulin producing cells and their output in Drosophila. Front. Physiol. 2013, 4, 252. [Google Scholar] [CrossRef]
- Gontijo, A.M.; Garelli, A. The biology and evolution of the Dilp8-Lgr3 pathway: A relaxin-like pathway coupling tissue growth and developmental timing control. Mech. Dev. 2018, 154, 44–50. [Google Scholar] [CrossRef]
- Imambocus, B.N.; Zhou, F.; Formozov, A.; Wittich, A.; Tenedini, F.M.; Hu, C.; Sauter, K.; Varela, E.M.; Herédia, F.; Casimiro, A.P.; et al. A neuropeptidergic circuit gates selective escape behavior of Drosophila larvae. Curr. Biol. 2022, 32, 149–163. [Google Scholar] [CrossRef]
- Belgacem, Y.H.; Martin, J.R. Hmgcr in the corpus allatum controls sexual dimorphism of locomotor activity and body size via the insulin pathway in Drosophila. PLoS ONE 2007, 2, e187. [Google Scholar] [CrossRef]
- Rauschenbach, I.Y.; Karpova, E.K.; Adonyeva, N.V.; Andreenkova, O.V.; Faddeeva, N.V.; Burdina, E.V.; Alekseev, A.A.; Menshanov, P.N.; Gruntenko, N.E. Disruption of insulin signalling affects the neuroendocrine stress reaction in Drosophila females. J. Exp. Biol. 2014, 217, 3733–3741. [Google Scholar] [CrossRef]
- Andreenkova, O.V.; Adonyeva, N.V.; Eremina, M.A.; Gruntenko, N.E.; Rauschenbach, I.Y. The Insulin-like receptor gene expression in the tissues synthesizing gonadotropic hormones at sexual maturation of Drosophila melanogaster females. Russ. J. Genet. 2016, 52, 1342–1344. [Google Scholar] [CrossRef]
- Gruntenko, N.E.; Rauschenbach, I.Y. The role of insulin signalling in the endocrine stress response in Drosophila melanogaster: A mini-review. Gen. Comp. Endocrinol. 2018, 258, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Yang, Z.; Yue, Z.; Alvarez, J.D.; Sehgal, A. FOXO and insulin signaling regulate sensitivity of the circadian clock to oxidative stress. Proc. Natl. Acad. Sci. USA 2007, 104, 15899–15904. [Google Scholar] [CrossRef] [PubMed]
- Puig, O.; Matilla, J. Understanding Forkhead box class O function: Lessons from Drosophila melanogaster. Antioxid. Redox Signal. 2011, 14, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yu, T.; Huang, P. Post-translational modifications of FOXO family proteins (Review). Mol. Med. Rep. 2016, 14, 4931–4941. [Google Scholar] [CrossRef]
- Gruntenko, N.E.; Adonyeva, N.V.; Burdina, E.V.; Karpova, E.K.; Andreenkova, O.V.; Gladkikh, D.V.; Ilinsky, Y.Y.; Rauschenbach, I.Y. The impact of FOXO on dopamine and octopamine metabolism in Drosophila under normal and heat stress conditions. Biol. Open. 2016, 5, 1706–1711. [Google Scholar] [CrossRef]
- Ding, K.; Barretto, E.C.; Johnston, M.; Lee, B.; Gallo, M.; Grewal, S.S. Transcriptome analysis of FOXO-dependent hypoxia gene expression identifies Hipk as a regulator of low oxygen tolerance in Drosophila. G3-Genes Genom. Genet. 2022, 12, jkac263. [Google Scholar] [CrossRef]
- Puig, O.; Marr, M.T.; Ruhf, M.L.; Tjian, R. Control of cell number by Drosophila FOXO: Downstream and feedback regulation of the insulin receptor pathway. Genes Dev. 2003, 17, 2006–2020. [Google Scholar] [CrossRef]
- Ikeya, T.; Galic, M.; Belawat, P.; Nairz, K.; Hafen, E. Nutrient dependent expression of insulin-like peptides from neuroendocrine cells in the CNS contributes to growth regulation in Drosophila. Curr. Biol. 2002, 12, 1293–1300. [Google Scholar] [CrossRef]
- Kim, S.K.; Rulifson, E.J. Conserved mechanisms of glucose sensing and regulation by Drosophila corpora cardiaca cells. Nature 2004, 431, 316–320. [Google Scholar] [CrossRef]
- Lee, G.; Park, J.H. Hemolymph sugar homeostasis and starvation-induced hyperactivity affected by genetic manipulations of the adipokinetic hormone-encoding gene in Drosophila melanogaster. Genetics 2004, 167, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Hughson, B.N. The glucagon-like adipokinetic hormone in Drosophila melanogaster—Biosynthesis and secretion. Front. Physiol. 2021, 12, 710652. [Google Scholar] [CrossRef] [PubMed]
- Isabel, G.; Martin, J.-R.; Chidami, S.; Veenstra, J.A.; Rosay, P. AKH-producing neuroendocrine cell ablation decreases trehalose and induces behavioral changes in Drosophila. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Sajwan, S.; Sidorov, R.; Stašková, T.; Žaloudíková, A.; Takasu, Y.; Kodrík, D.; Zurovec, M. Targeted mutagenesis and functional analysis of adipokinetic hormone-encoding gene in Drosophila. Insect Biochem. Mol. Biol. 2015, 61, 79–86. [Google Scholar] [CrossRef]
- Gáliková, M.; Diesner, M.; Klepsatel, P.; Hehlert, P.; Xu, Y.; Bickmeyer, I.; Predel, R.; Kühnlein, R.P. Energy homeostasis control in Drosophila adipokinetic hormone mutants. Genetics 2015, 201, 665–683. [Google Scholar] [CrossRef]
- Bednářová, A.; Tomčala, A.; Mochanová, M.; Kodrík, D.; Krishnan, N. Disruption of adipokinetic hormone mediated energy homeostasis has subtle effects on physiology, behavior and lipid status during aging in Drosophila. Front. Physiol. 2018, 9, 949. [Google Scholar] [CrossRef]
- Toivonen, J.M.; Partridge, L. Endocrine regulation of aging and reproduction in Drosophila. Mol. Cell Endocrinol. 2009, 299, 39–50. [Google Scholar] [CrossRef]
- Zhang, X.; Li, S.; Liu, S. Juvenile hormone studies in Drosophila melanogaster. Front. Physiol. 2022, 12, 785320. [Google Scholar] [CrossRef]
- Lafont, R.; Dauphin-Villemant, C.; Warren, J.T.; Rees, H.H. Ecdysteroid chemistry and biochemistry. Ref. Mod. Life Sci. 2017, 3, 125–195. [Google Scholar] [CrossRef]
- Sasaki, K.; Harano, K.-I. Multiple regulatory roles of dopamine in behavior and reproduction of social insects. Trends Entomol. 2010, 6, 1–13. [Google Scholar]
- Farooqui, T. Octopamine-mediated neuromodulation of insect senses. Neurochem. Res. 2007, 32, 1511–1529. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.A.; Krantz, D.E. Drosophila melanogaster as a genetic model system to study neurotransmitter transporters. Neurochem. Int. 2014, 73, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Gruntenko, N.E.; Rauschenbach, I.Y. Interplay of JH, 20E and biogenic amines under normal and stress conditions and its effects on reproduction. J. Insect Physiol. 2008, 54, 902–908. [Google Scholar] [CrossRef]
- Rauschenbach, I.Y. Stress response in insects: Mechanism, genetic control, and role in adaptation. Russ. J. Genet. 1997, 33, 942–949. [Google Scholar]
- Hirashima, A.; Nagano, T.; Eto, M. Stress-induced changes in the biogenic amine levels and larval growth of Tribolium castaneum Herbst. Biosci. Biotech. Biochem. 1993, 57, 2085–2089. [Google Scholar] [CrossRef]
- Jünger, M.A.; Rintelen, F.; Stocker, H.; Wasserman, J.D.; Végh, M.; Radimerski, T.; Greenberg, M.E.; Hafen, E. The Drosophila forkhead transcription factor FOXO mediates the reduction in cell number associated with reduced insulin signaling. J. Biol. Chem. 2003, 2, 20. [Google Scholar] [CrossRef]
- Hwangbo, D.S.; Gershman, B.; Tu, M.P.; Palmer, M.; Tatar, M. Drosophila dFOXO controls lifespan and regulates insulin signalling in brain and fat body. Nature 2004, 429, 562–566. [Google Scholar] [CrossRef]
- Eremina, M.A.; Menshanov, P.N.; Shishkina, O.D.; Gruntenko, N.E. The transcription factor dFOXO controls the expression of insulin pathway genes and lipids content under heat stress in Drosophila melanogaster. Vavilov J. Genet. Breed. 2021, 25, 465–471. [Google Scholar] [CrossRef]
- Zhang, Y.; Xi, Y. Fat body development and its function in energy storage and nutrient sensing in Drosophila melanogaster. J. Tissue Sci. Eng. 2014, 6, 1. [Google Scholar] [CrossRef]
- Slaidina, M.; Delanoue, R.; Grönke, S.; Partridge, L.; Leopold, P. A Drosophila insulin-like peptide promotes growth during nonfeeding states. Dev. Cell 2009, 17, 874–884. [Google Scholar] [CrossRef]
- Okamoto, N.; Yamanaka, N.; Yagi, Y.; Nishida, Y.; Kataoka, H.; O’Connor, M.B.; Mizoguchi, A. A fat body-derived IGF-like peptide regulates postfeeding growth in Drosophila. Dev. Cell 2009, 17, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Colombani, J.; Bianchini, L.; Layalle, S.; Pondeville, E.; Dauphin-Villemant, C.; Antoniewski, C.; Carré, C.; Noselli, S.; Léopold, P. Antagonistic actions of ecdysone and insulins determine final size in Drosophila. Science 2005, 310, 667–670. [Google Scholar] [CrossRef]
- Hossain, M.S.; Liu, Y.; Zhou, S.; Li, K.; Tian, L.; Li, S. 20-Hydroxyecdysone-induced transcriptional activity of FoxO upregulates brummer and acid lipase-1 and promotes lipolysis in Bombyx fat body. Insect Biochem. Mol. Biol. 2013, 43, 829–838. [Google Scholar] [CrossRef]
- Khlebodarova, T.M.; Gruntenko, N.E.; Grenback, L.G.; Sukhanova, M.Z.; Mazukov, M.M.; Rauschenbach, I.Y.; Tomas, B.A.; Hammock, B.D. A comparative analysis of juvenile hormone metabolizing enzymes in two species of Drosophila during development. Insect Biochem. Mol. Biol. 1996, 26, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Borovsky, D.; Breyssens, H.; Buytaert, E.; Peeters, T.; Laroye, C.; Stoffels, K.; Rougé, P. Cloning and characterization of Drosophila melanogaster juvenile hormone epoxide hydrolases (JHEH) and their promoters. Biomolecules 2022, 12, 991. [Google Scholar] [CrossRef]
- Raushenbach, I.Y.; Gruntenko, N.E.; Bownes, M.; Adonieva, N.V.; Terashima, J.; Karpova, E.K.; Faddeeva, N.V.; Chentsova, N.A. The role of juvenile hormone in the control of reproductive function in Drosophila virilis under nutritional stress. J. Insect Physiol. 2004, 50, 323–330. [Google Scholar] [CrossRef]
- Rauschenbach, I.Y.; Karpova, E.K.; Gruntenko, N.E. dFOXO transcription factor regulates juvenile hormone metabolism in Drosophila melanogaster females. Russ. J. Genet. 2015, 51, 932–934. [Google Scholar] [CrossRef]
- Mirth, C.K.; Tang, H.Y.; Makohon-Moore, S.C.; Salhadar, S.; Gokhale, R.H.; Warner, R.D.; Koyama, T.; Riddiford, L.M.; Shingleton, A.W. Juvenile hormone regulates body size and perturbs insulin signaling in Drosophila. Proc. Natl. Acad. Sci. USA 2014, 111, 7018–7023. [Google Scholar] [CrossRef]
- Terashima, J.; Takaki, K.; Sakurai, S.; Bownes, M. Nutritional status affects 20-hydroxyecdysone concentration and progression of oogenesis in Drosophila melanogaster. J Endocrinol. 2005, 187, 69–79. [Google Scholar] [CrossRef]
- Crocker, A.; Shahidullah, M.; Levitan, I.B.; Sehgal, A. Identification of a neural circuit that underlies the effects of octopamine on sleep: Wake behavior. Neuron 2010, 65, 670–681. [Google Scholar] [CrossRef]
- Luo, J.; Lushchak, O.V.; Goergen, P.; Williams, M.J.; Nässel, D.R. Drosophila insulin-producing cells are differentially modulated by serotonin and octopamine receptors and affect social behavior. PLoS ONE 2014, 9, e99732. [Google Scholar] [CrossRef]
- Gruntenko, N.E.; Wilson, T.G.; Monastirioti, M.; Rauschenbach, I.Y. Stress-reactivity and juvenile hormone degradation in Drosophila melanogaster strains having stress-related mutations. Insect Biochem. Mol. Biol. 2000, 30, 775–783. [Google Scholar] [CrossRef]
- Gruntenko, N.E.; Karpova, E.K.; Alekseev, A.A.; Chentsova, N.A.; Bogomolova, E.V.; Bownes, M.; Rauschenbach, I.Y. Effects of octopamine on reproduction, juvenile hormone metabolism, dopamine, and 20-hydroxyecdysone contents in Drosophila. Arch. Insect Biochem. Physiol. 2007, 65, 85–94. [Google Scholar] [CrossRef]
- Andreatta, G.; Kyriacou, C.P.; Flatt, T.; Costa, R. Aminergic signaling controls ovarian dormancy in Drosophila. Sci. Rep. 2018, 8, 2030. [Google Scholar] [CrossRef]
- Gruntenko, N.E.; Laukhina, O.V.; Rauschenbach, I.Y. Role of D1- and D2-like receptors in age-specific regulation of juvenile hormone and 20-hydroxyecdysone levels by dopamine in Drosophila. J. Insect Physiol. 2012, 58, 1534–1540. [Google Scholar] [CrossRef]
- Tatar, M.; Kopelman, A.; Epstein, D.; Tu, M.P.; Yin, C.M.; Garofalo, R.S. A mutant Drosophila insulin receptor homolog that extends lifespan and impairs neuroendocrine function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef]
- Tu, M.P.; Yin, C.M.; Tatar, M. Impaired ovarian ecdysone synthesis of Drosophila melanogaster insulin receptor mutants. Aging Cell 2002, 1, 158–160. [Google Scholar] [CrossRef]
- Tu, M.P.; Yin, C.M.; Tatar, M. Mutations in insulin signaling pathway alter juvenile hormone synthesis in Drosophila melanogaster. Gen. Comp. Endocrinol. 2005, 142, 347–356. [Google Scholar] [CrossRef]
- Bogomolova, E.V.; Adonyeva, N.V.; Karpova, E.K.; Gruntenko, N.E.; Rauschenbach, I.Y. Ubiquitous downregulation of InR gene expression affects stress associated hormone metabolism in Drosophila females. Russ. J. Genet. 2013, 49, 778–781. [Google Scholar] [CrossRef]
- Karpova, E.K.; Adonyeva, N.V.; Faddeeva, N.V.; Romanova, I.V.; Gruntenko, N.E.; Rauschenbach, I.Y. Insulin affects repro-duction and juvenile hormone metabolism under normal and stressful conditions in Drosophila females. Dokl. Biochem. Biophys. 2013, 452, 264–266. [Google Scholar] [CrossRef]
- Mattila, J.; Hietakangas, V. Regulation of carbohydrate energy metabolism in Drosophila melanogaster. Genetics 2017, 207, 1231–1253. [Google Scholar] [CrossRef]
- Murillo-Maldonado, J.M.; Sánchez-Chávez, G.; Salgado, L.M.; Salceda, R.; Riesgo-Escovar, J.R. Drosophila insulin pathway mutants affect visual physiology and brain function besides growth, lipid, and carbohydrate metabolism. Diabetes 2011, 60, 1632–1636. [Google Scholar] [CrossRef]
- Eremina, M.A.; Karpova, E.K.; Rauschenbach, I.Y.; Pirozhkova, D.S.; Andreenkova, O.V.; Gruntenko, N.E. Mutations in the insulin signaling pathway genes affect carbohydrate level under heat stress in Drosophila melanogaster females. Russ. J. Genet. 2019, 55, 519–522. [Google Scholar] [CrossRef]
- Karpova, E.K.; Eremina, M.A.; Pirozhkova, D.S.; Gruntenko, N.E. Stress-related hormones affect carbohydrate metabolism in Drosophila females. Arch. Insect Biochem. Physiol. 2019, 101, e21540. [Google Scholar] [CrossRef]
- Benoit, J.B.; Lopez-Martinez, G.; Elnitsky, M.A.; Lee, R.E., Jr.; Denlinger, D.L. Dehydration-induced cross tolerance of Belgica antarctica larvae to cold and heat is facilitated by trehalose accumulation. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 152, 518–523. [Google Scholar] [CrossRef]
- Klepsatel, P.; Gáliková, M.; Xu, Y.; Kühnlein, R.P. Thermal stress depletes energy reserves in Drosophila. Sci. Rep. 2016, 6, e33667. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Roeder, T. Tyramine and octopamine: Ruling behavior and metabolism. Annu. Rev. Entomol. 2005, 50, 447–477. [Google Scholar] [CrossRef]
- Farooqui, T. Review of octopamine in insect nervous systems. Open Access Insect Physiol. 2012, 4, 1–17. [Google Scholar] [CrossRef]
- Yu, J.; Wang, H.; Chen, W.; Song, H.; Wang, Y.; Liu, Z.; Xu, B. 20-Hydroxyecdysone and receptor interplay in the regulation of hemolymph glucose level in honeybee (Apis mellifera) larvae. Metabolites 2023, 13, 80. [Google Scholar] [CrossRef]
- Wang, X.P.; Huang, Z.; Li, Y.-L.; Jin, K.Y.; Dong, D.J.; Wang, J.X.; Zhao, X.F. Krüppel-like factor 15 integrated autophagy and gluconeogenesis to maintain glucose homeostasis under 20-hydroxyecdysone regulation. PLoS Genet. 2022, 18, e1010229. [Google Scholar] [CrossRef]
- Li, Y.N.; Lio, Y.-B.; Xie, X.-Q.; Zhang, J.N.; Li, W.L. The modulation of trehalose metabolism by 20-hydroxyecdysone in Antheraea pernyi (Lepidoptera: Saturniidae) during its diapause termination and post-termination period. J. Insect Sci. 2020, 20, 5. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Gao, H.; Askar, A.; Li, X.P.; Zhang, G.C.; Jing, T.Z.; Zou, H.; Guan, H.; Zhao, Y.H.; Zou, C.S. Steroid hormone 20-hydroxyecdysone disturbs fat body lipid metabolism and negatively regulates gluconeogenesis in Hyphantria cunea larvae. J. Insect Sci. 2022. Epub ahead of print. [Google Scholar] [CrossRef]
- Bobrovskikh, M.A.; Gruntenko, N.E. The role of 20-hydroxyecdysone in the control of carbohydrate levels in Drosophila melanogaster under short-term heat stress. Dokl. Biochem. Biophys. 2023, in press. [Google Scholar]
- Xu, J.; Sheng, Z.; Palli, S.R. Juvenile hormone and insulin regulate trehalose homeostasis in the red flour beetle, Tribolium castaneum. PLoS Genet. 2013, 9, e1003535. [Google Scholar] [CrossRef]
- Ugrankar, R.; Theodoropoulos, P.; Akdemir, F.; Henne, W.M.; Graff, J.M. Circulating glucose levels inversely correlate with Drosophila larval feeding through insulin signaling and SLC5A11. Commun. Biol. 2018, 1, 110. [Google Scholar] [CrossRef]
- Pascual, A.; Préat, T. Localization of long-term memory within the Drosophila mushroom body. Science 2001, 294, 1115–1117. [Google Scholar] [CrossRef]
- Plaçais, P.-Y.; Tredern, É.; Scheunemann, L.; Trannoy, S.; Goguel, V.; Han, K.; Isabel, G.; Preat, T. Upregulated energy metabolism in the Drosophila mushroom body is the trigger for long-term memory. Nat. Commun. 2017, 8, 15510. [Google Scholar] [CrossRef]
- Chia, J.; Scott, K. Activation of specific mushroom body output neurons inhibits proboscis extension and sucrose consumption. PLoS ONE 2020, 15, e0223034. [Google Scholar] [CrossRef]
- Rauschenbach, I.Y.; Serova, L.I.; Timochina, I.S.; Chentsova, N.A.; Schumnaja, L.V. Analysis of differences in dopamine content between two lines of Drosophila virilis in response to heat stress. J. Insect Physiol. 1993, 39, 761–767. [Google Scholar] [CrossRef]
- Hirashima, A.; Sukhanova, M.J.; Rauschenbach, I.Y. Biogenic amines in Drosophila virilis under stress conditions. Biosci. Biotechnol. Biochem. 2000, 64, 2625–2630. [Google Scholar] [CrossRef]
- Hirashima, A.; Rauschenbach, I.Y.; Sukhanova, M.J. Ecdysteroids in stress responsive and nonresponsive Drosophila virilis lines under stress conditions. Biosci. Biotechnol. Biochem. 2000, 64, 2657–2662. [Google Scholar] [CrossRef]
- Gruntenko, N.E.; Khlebodarova, T.M.; Sukhanova, M.J.; Vasenkova, I.A.; Kaidanov, L.Z.; Rauschenbach, I.Y. Prolonged negative selection of Drosophila melanogaster for a character of adaptive significance disturbs stress reactivity. Insect Biochem. Mol. Biol. 1999, 29, 445–452. [Google Scholar] [CrossRef]
- Chentsova, N.A.; Gruntenko, N.E.; Rauschenbach, I.Y. Ecdysone 20-monooxygenase activity in Drosophila virilis strains varying in ecdysteroid response to heat stress. Russ. J. Genet. 2007, 43, 829–830. [Google Scholar] [CrossRef]
- Sukhanova, M.Z.; Grenback, L.G.; Gruntenko, N.E.; Khlebodarova, T.M.; Rauschenbach, I.Y. Alkaline phosphatase in Drosophila under heat stress. J. Insect Physiol. 1995, 42, 161–165. [Google Scholar] [CrossRef]
- Sukhanova, M.Z.; Grenback, L.G.; Gruntenko, N.E.; Khlebodarova, T.M.; Rauschenbach, I.Y. Tyrosine decarboxylase and dopa decarboxylase in Drosophila virilis under heat stress. Biochem. Genet. 1997, 35, 91–103. [Google Scholar] [CrossRef]
- Rauschenbach, I.Y.; Sukhanova, M.J.; Shumnaya, L.V.; Gruntenko, N.E.; Grenback, L.G.; Khlebodarova, T.M.; Chentsova, N.A. Role of DOPA decarboxylase and N-acetyltransferase in regulation of dopamine content in Drosophila virilis under normal and stress conditions. Insect Biochem. Molec. Biol. 1997, 27, 729–734. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bobrovskikh, M.A.; Gruntenko, N.E. Mechanisms of Neuroendocrine Stress Response in Drosophila and Its Effect on Carbohydrate and Lipid Metabolism. Insects 2023, 14, 474. https://doi.org/10.3390/insects14050474
Bobrovskikh MA, Gruntenko NE. Mechanisms of Neuroendocrine Stress Response in Drosophila and Its Effect on Carbohydrate and Lipid Metabolism. Insects. 2023; 14(5):474. https://doi.org/10.3390/insects14050474
Chicago/Turabian StyleBobrovskikh, Margarita A., and Nataly E. Gruntenko. 2023. "Mechanisms of Neuroendocrine Stress Response in Drosophila and Its Effect on Carbohydrate and Lipid Metabolism" Insects 14, no. 5: 474. https://doi.org/10.3390/insects14050474
APA StyleBobrovskikh, M. A., & Gruntenko, N. E. (2023). Mechanisms of Neuroendocrine Stress Response in Drosophila and Its Effect on Carbohydrate and Lipid Metabolism. Insects, 14(5), 474. https://doi.org/10.3390/insects14050474