
Monitoring and Prediction of Siberian Silk Moth Dendrolimus sibiricus Tschetv. (Lepidoptera: Lasiocampidae) Outbreaks Using Remote Sensing Techniques

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
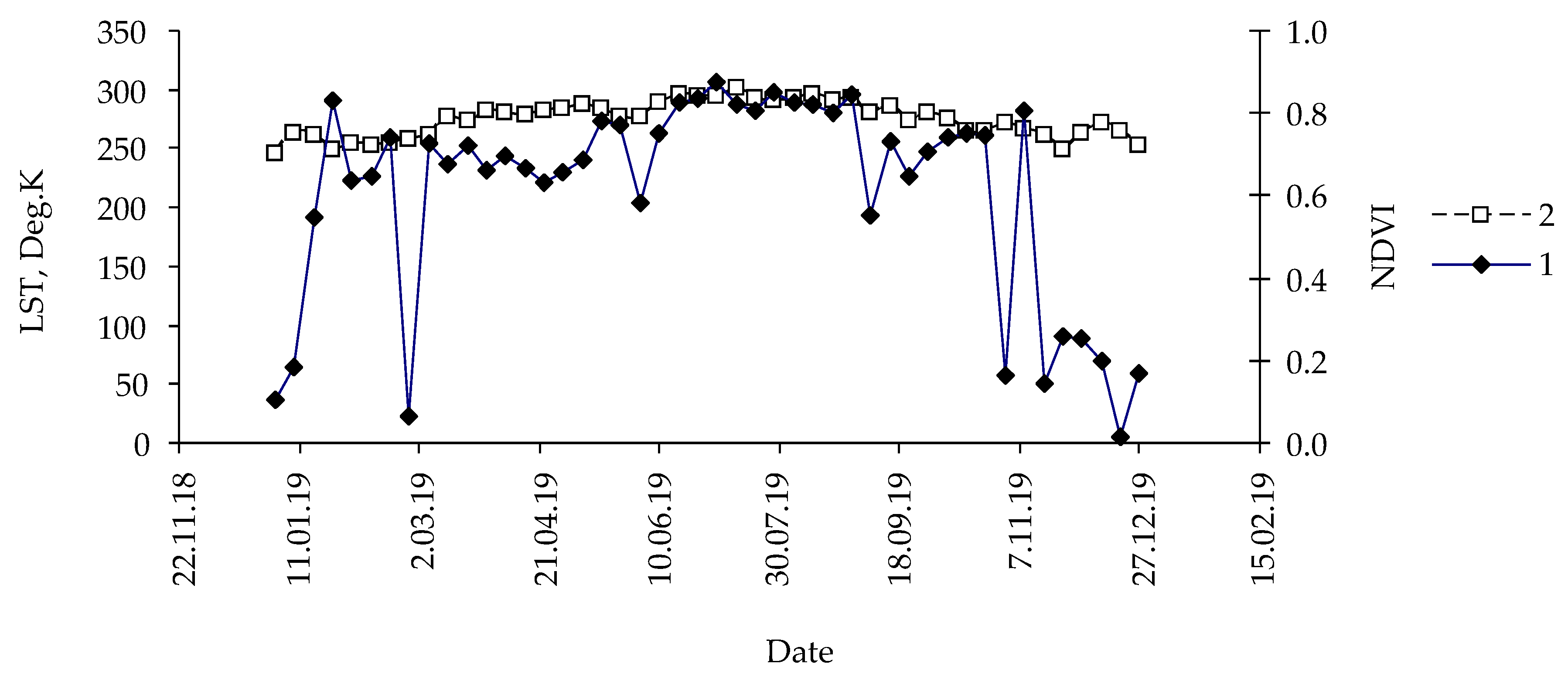
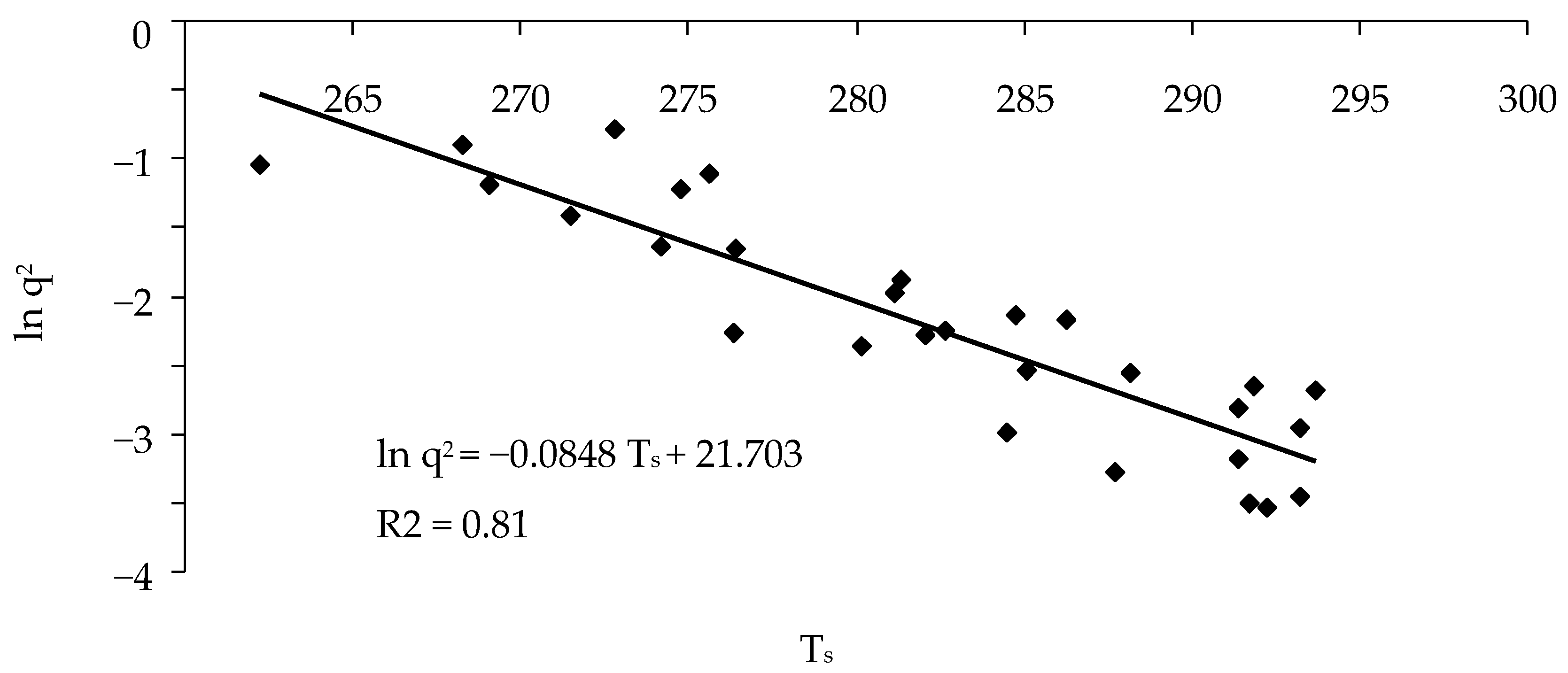
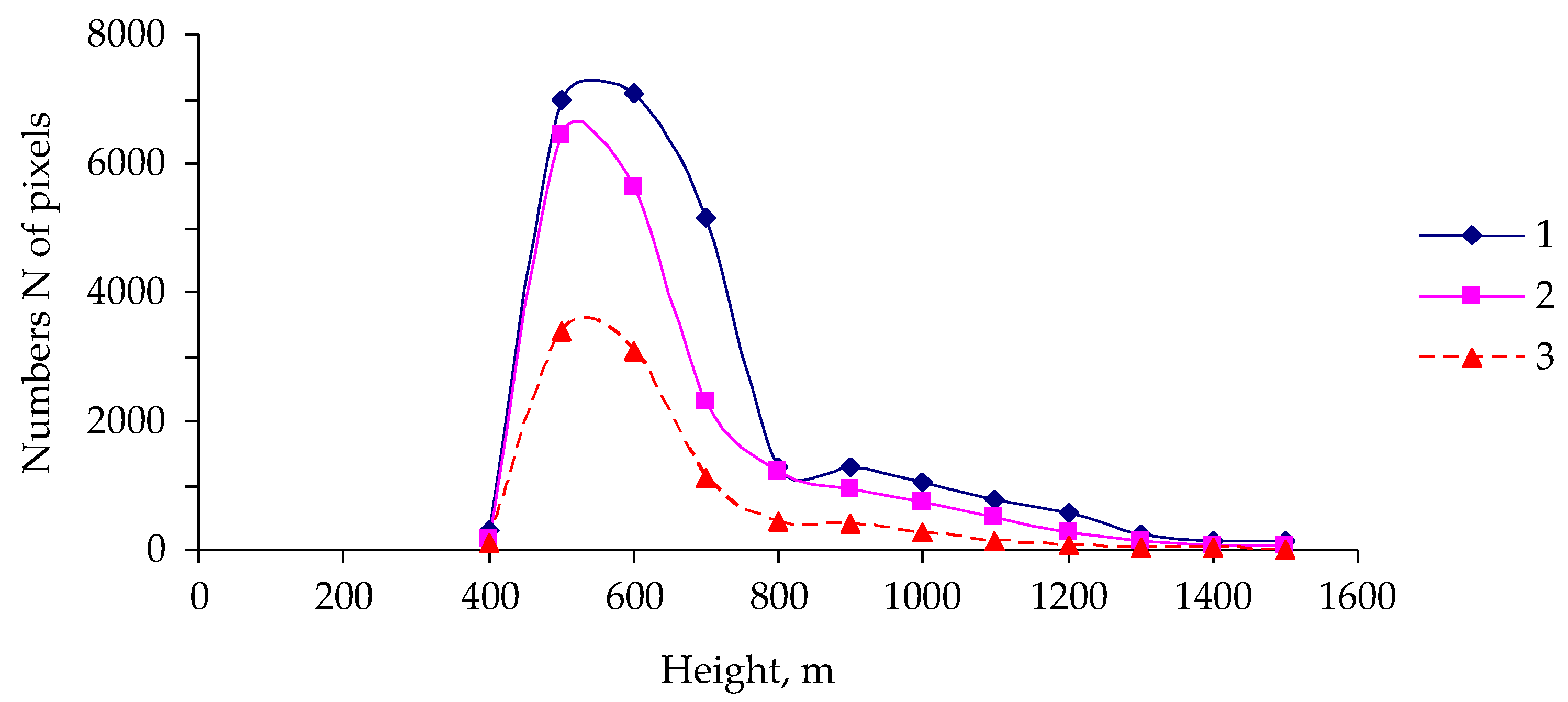
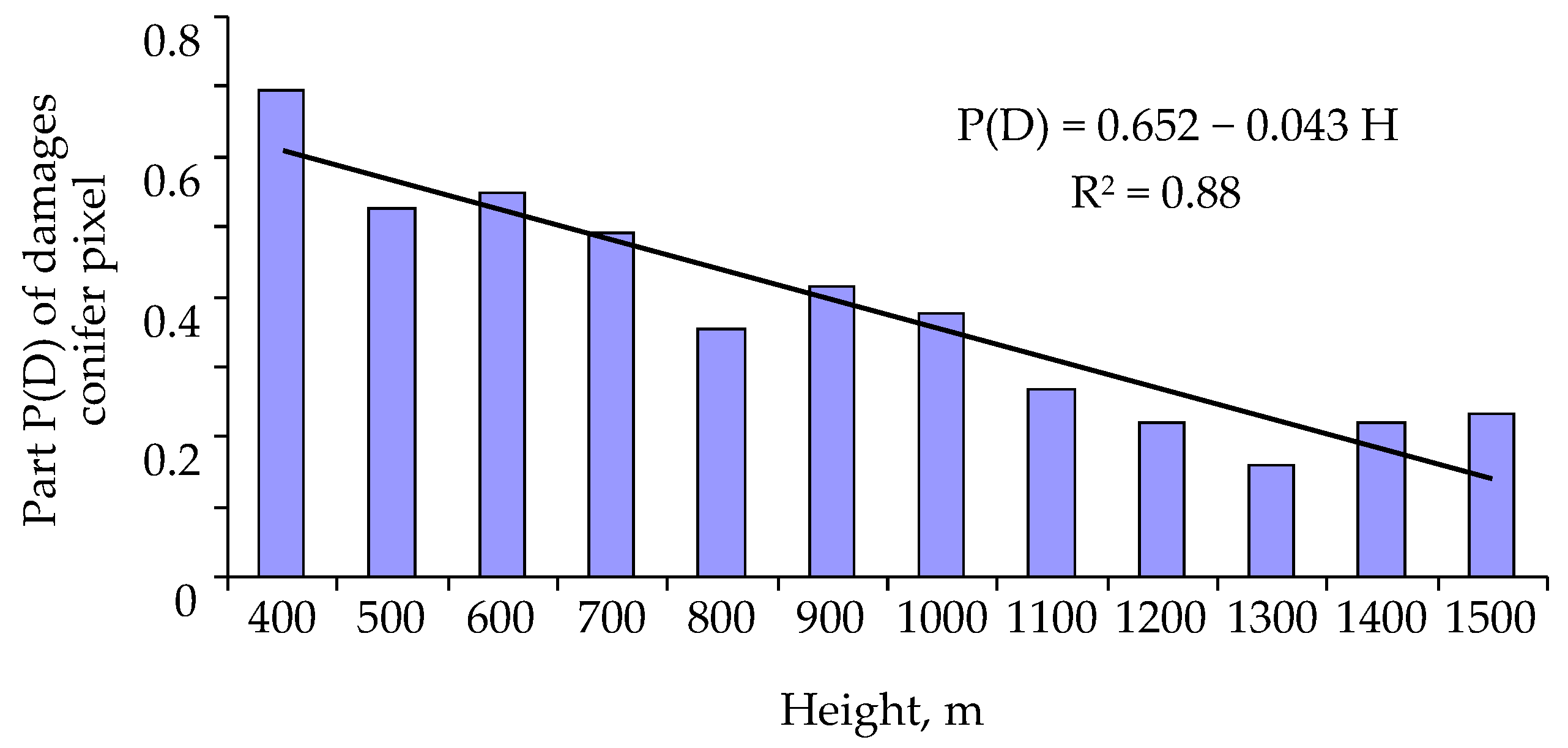
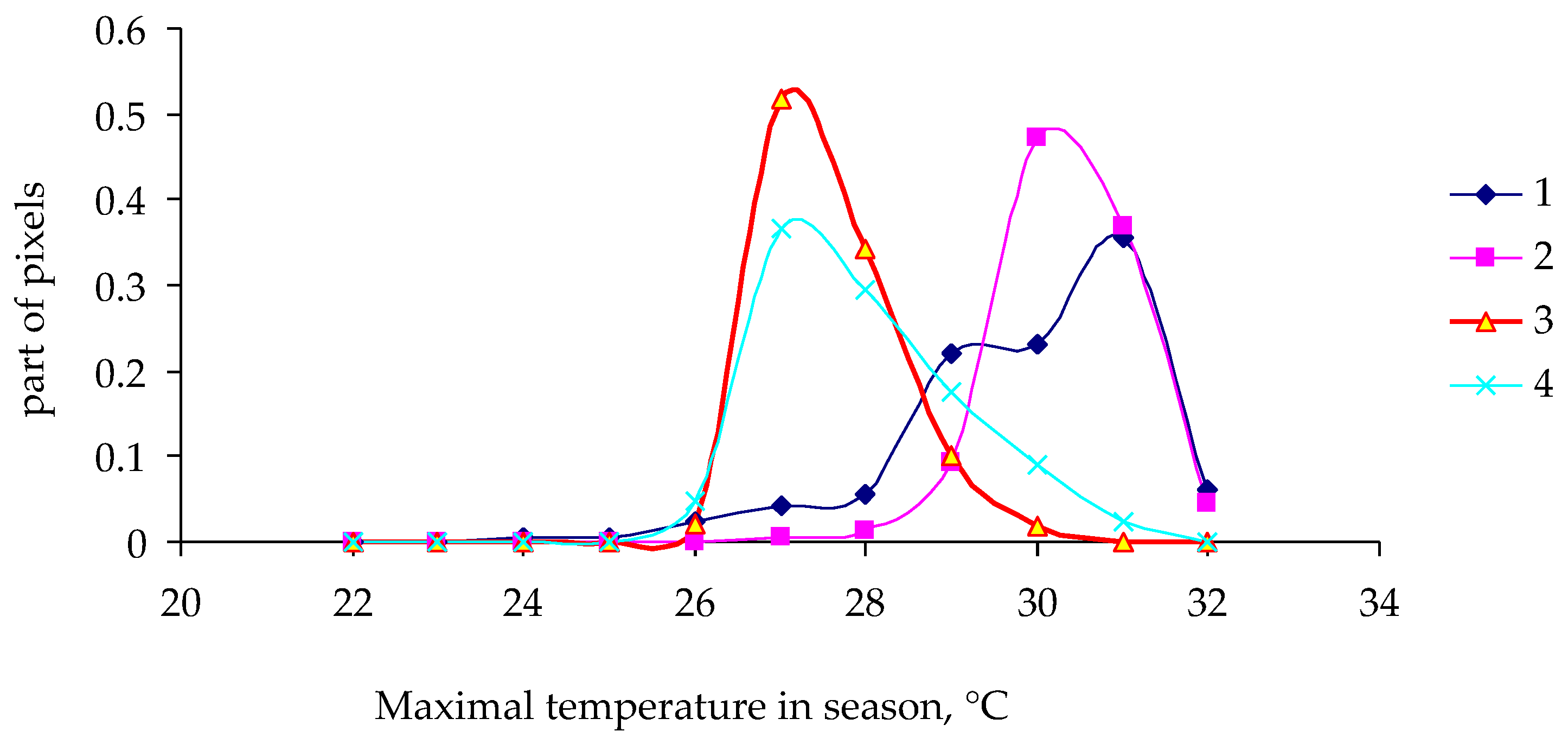
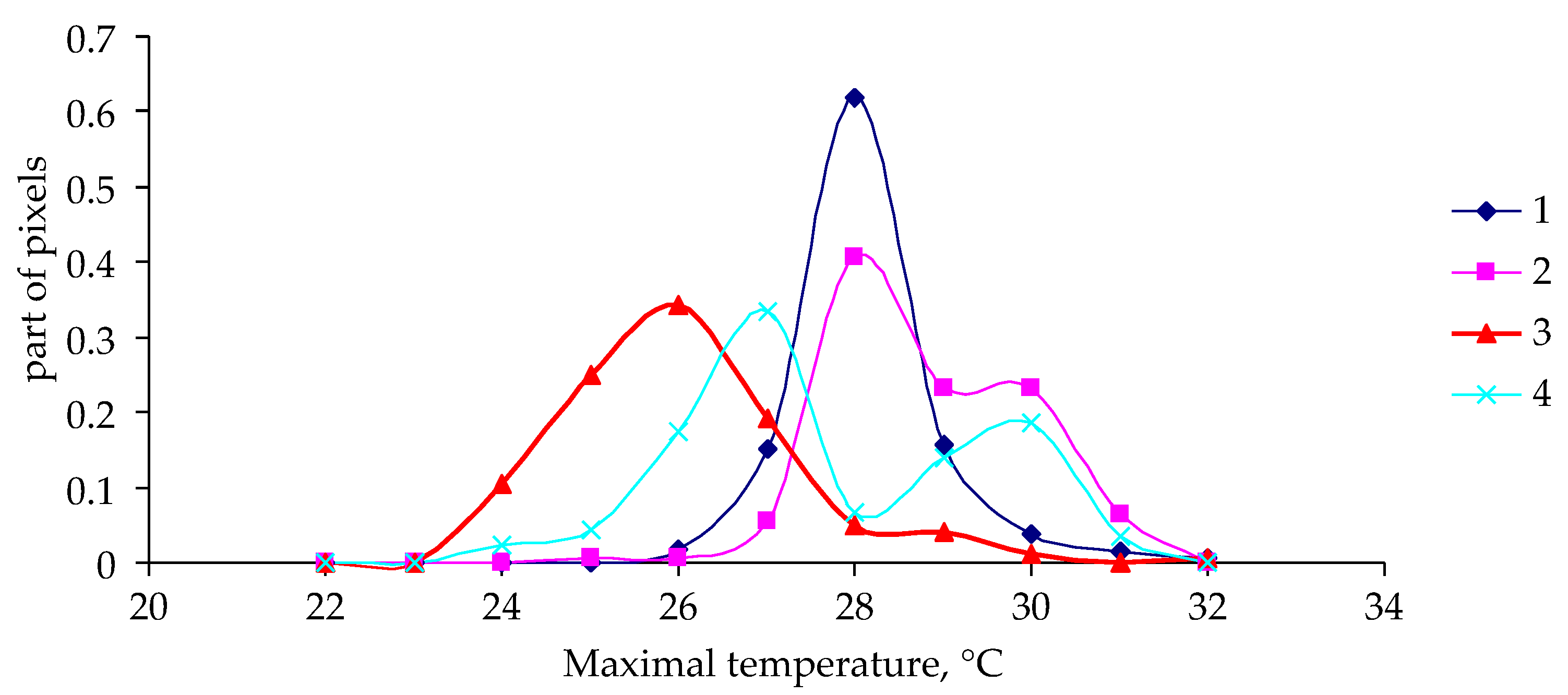
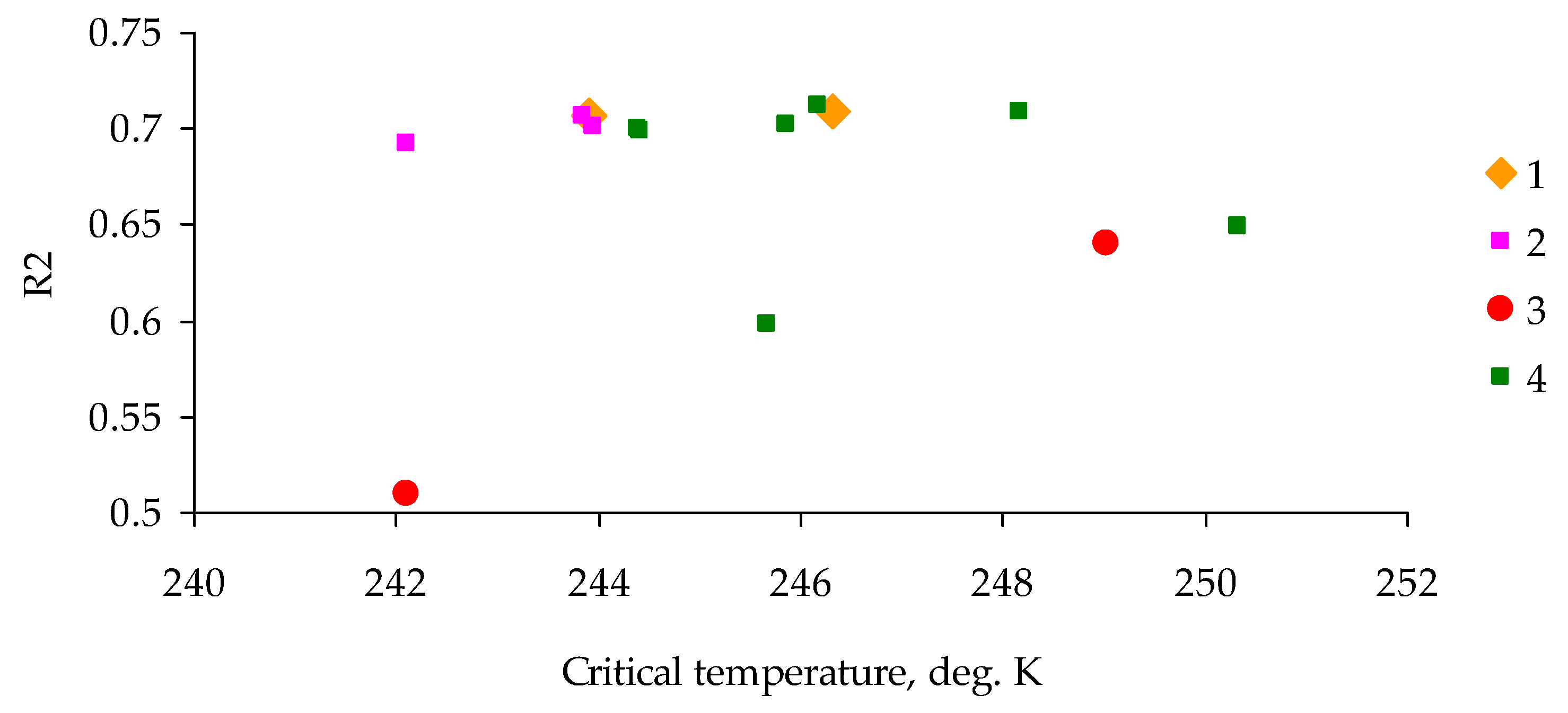
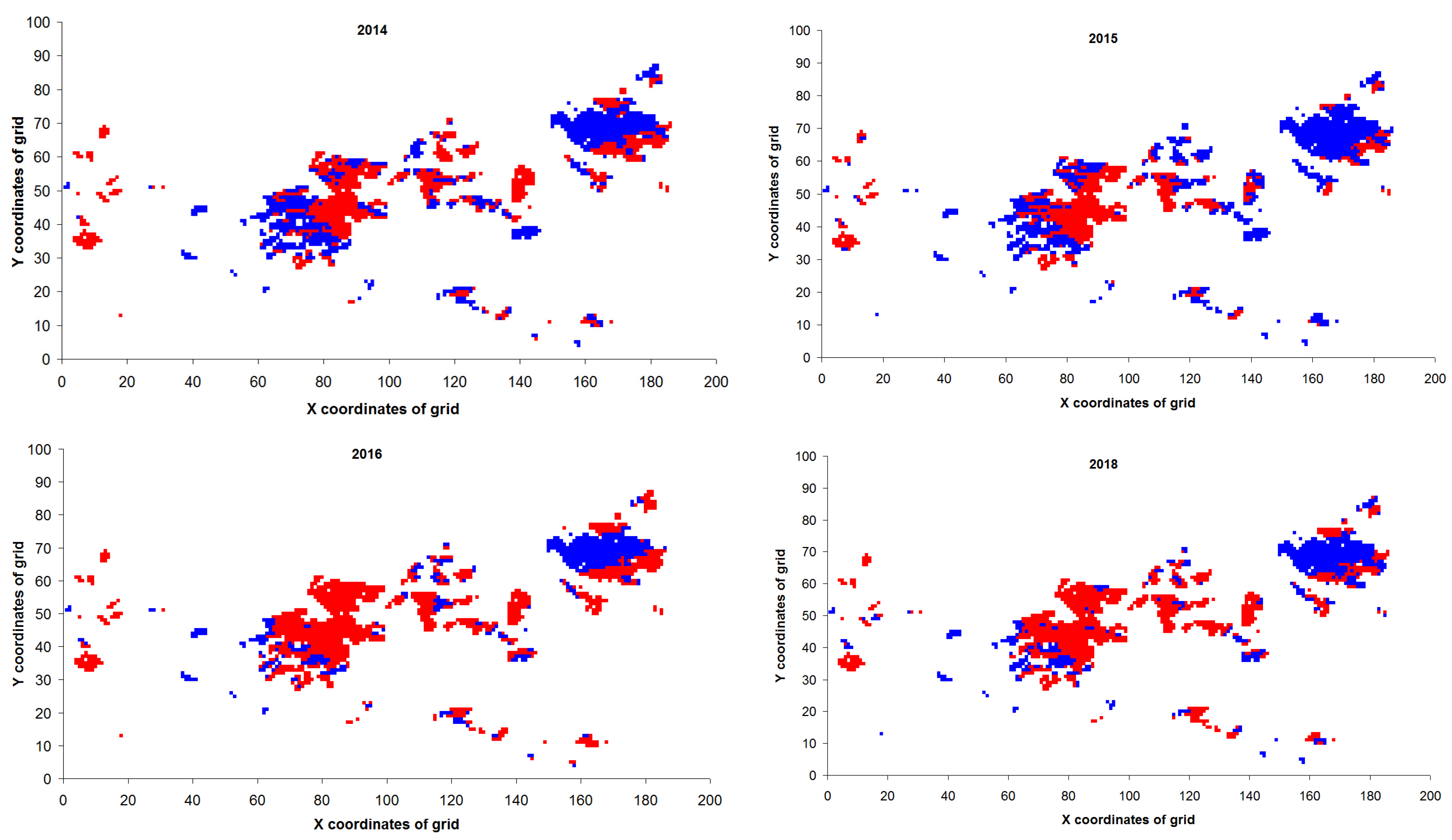
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Correction Statement
References
- Sproull, G.J.; Adamus, M.; Szewczyk, J.; Kersten, G.; Szwagrzyk, J. Fine-scale spruce mortality dynamics driven by bark beetle disturbance in Babia Góra National Park. Poland. Eur. J. For. Res. 2016, 135, 507–517. [Google Scholar] [CrossRef]
- Overbeck, M.; Schmidt, M. Modelling infestation risk of Norway spruce by Ips typographus (L.) in the Lower Saxon Harz Mountains (Germany). For. Ecol. Manag. 2012, 266, 115–125. [Google Scholar] [CrossRef]
- Stadelmann, G.; Bugmann, H.; Wermelinger, B.; Meier, F.; Bigler, C. A predictive framework to assess spatio-temporal variability of infestations by the European spruce bark beetle. Ecography 2013, 36, 1208–1217. [Google Scholar] [CrossRef]
- Kärvemo, S.; Van Boeckel, T.P.; Gilbert, M.; Grégoire, J.C.; Schroeder, M. Large-scale risk mapping of an eruptive bark beetle—Importance of forest susceptibility and beetle pressure. For. Ecol. Manag. 2014, 318, 158–166. [Google Scholar] [CrossRef]
- Soukhovolsky, V.; Kovalev, A.; Tarasova, O.; Modlinger, R.; Krenova, Z.; Mezei, P.; Jaroslav, S.; Rožnovský, J.; Korolyova, N.; Majdák, A.; et al. Wind Damage and Temperature Effect on Tree Mortality Caused by Ips Typographus L.: Phase Transition Model. Forests 2022, 13, 180. [Google Scholar] [CrossRef]
- Senf, C.; Campbell, E.; Wulder, M.; Pflugmacher, D.; Hostert, P. A multi-scale analysis of western spruce budworm outbreak dynamics. Landsc. Ecol. 2017, 32, 501–514. [Google Scholar] [CrossRef]
- Seidl, R.; Müller, J.; Hothorn, T.; Bässler, C.; Heurich, M.; Kautz, M. Small beetle, large-scale drivers: How regional and landscape factors affect outbreaks of the european spruce bark beetle. J. Appl. Ecol. 2016, 53, 530–540. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. Bioscience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Fernández, A.; Fort, H. Catastrophic phase transitions and early warnings in a spatial ecological model. J. Stat. Mech. Theor. Exp. 2009, 2009, P09014. [Google Scholar] [CrossRef]
- Lewis, M.A.; Nelson, W.; Xu, C. A structured threshold model for mountain pine beetle outbreak. Bull. Math. Biol. 2010, 72, 565–589. [Google Scholar] [CrossRef]
- Fahse, L.; Heurich, M. Simulation and analysis of outbreaks of bark beetle infestations and their management at the stand level. Ecol. Model. 2011, 222, 1833–1846. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R.; Hostert, P. Remote sensing of forest insect disturbances: Current state and future directions. Int. J. Appl. Earth Obs. Geoinf. 2017, 60, 49–60. [Google Scholar] [CrossRef]
- Wulder, M.A.; Dymond, C.C.; White, J.C.; Leckie, D.G.; Carroll, A.L. Surveying mountain pine beetle damage of forests: A review of remote sensing opportunities. For. Ecol. Manag. 2006, 221, 27–41. [Google Scholar] [CrossRef]
- Long, J.A.; Lawrence, R.L. Mapping Percent Tree Mortality Due to Mountain Pine Beetle Damage. For. Sci. 2016, 62, 392–402. [Google Scholar] [CrossRef]
- Meddens, A.J.H.; Hicke, J.A.; Vierling, L.A.; Hudak, A.T. Evaluating methods to detect bark beetle-caused tree mortality using single-date and multi-date Landsat imagery. Remote Sens. Environ. 2013, 132, 49–58. [Google Scholar] [CrossRef]
- Meigs, G.W.; Kennedy, R.E.; Cohen, W.B. A Landsat time series approach to characterize bark beetle and defoliator impacts on tree mortality and surface fuels in conifer forests. Remote Sens. Environ. 2011, 115, 3707–3718. [Google Scholar] [CrossRef]
- Skakun, R.S.; Wulder, M.A.; Franklin, S.E. Sensitivity of the thematic mapper enhanced wetness difference index to detect mountain pine beetle red-attack damage. Remote Sens. Environ. 2003, 86, 433–443. [Google Scholar] [CrossRef]
- Anees, A.; Aryal, J. Near-Real Time Detection of Beetle Infestation in Pine Forests Using MODIS Data. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 3713–3723. [Google Scholar] [CrossRef]
- Spruce, J.P.; Hicke, J.A.; Hargrove, W.W.; Grulke, N.E.; Meddens, A.J.H. Use of MODIS NDVI Products to Map Tree Mortality Levels in Forests Affected by Mountain Pine Beetle Outbreaks. Forests 2019, 10, 811. [Google Scholar] [CrossRef]
- Latifi, H.; Dahms, T.; Beudert, B.; Heurich, M.; Kübert, C.; Dech, S. Synthetic RapidEye data used for the detection of area-based spruce tree mortality induced by bark beetles. GIScience Remote Sens. 2018, 55, 839–859. [Google Scholar] [CrossRef]
- Nansen, C.; Elliott, N. Remote sensing and reflectance profiling in entomology. Annu. Rev. Entomol. 2016, 61, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Riley, J.R. Remote sensing in entomology. Annu. Rev. Entomol. 1989, 34, 247–271. [Google Scholar] [CrossRef]
- Latchininsky, A.V. Locusts and remote sensing: A review. J. Appl. Remote Sens. 2013, 7, 075099. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, Y.; Pu, R.; Gonzalez-Moreno, P.; Yuan, L.; Wu, K.; Huang, W. Monitoring plant diseases and pests through remote sensing technology: A review. Comput. Electron. Agric. 2019, 165, 104943. [Google Scholar] [CrossRef]
- Iost Filho, F.H.; Heldens, W.B.; Kong, Z.; de Lange, E.S. Drones: Innovative technology for use in precision pest management. J. Econ. Entomol. 2020, 113, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A. Principles of Thermal Ecology: Temperature, Energy, and Life; Oxford University Press: Oxford, UK, 2017; p. 480. [Google Scholar]
- Potter, K.A.; Woods, H.A.; Pincebourde, S. Microclimatic challenges in global change biology. Glob. Change Biol. 2013, 19, 2932–2939. [Google Scholar] [CrossRef]
- Bramer, I.; Anderson, B.J.; Bennie, J.; Bladon, A.J.; De Frenne, P.; Hemming, D.; Hill, R.A.; Kearney, M.R.; Körner, C.; Korstjens, A.H.; et al. Advances in monitoring and modelling climate at ecologically relevant scales. In Advances in Ecological Research; Woodward, G., Jackson, M., Eds.; Elsevier: London, UK, 2018; pp. 101–161. [Google Scholar]
- Suggitt, A.J.; Platts, P.J.; Barata, I.M.; Bennie, J.J.; Burgess, M.D.; Bystriakova, N.; Duffield, S.; Ewing, S.R.; Gillingham, P.K.; Harper, A.B.; et al. Conducting robust ecological analyses with climate data. Oikos 2017, 126, 1533–1541. [Google Scholar] [CrossRef]
- Spruce, J.P.; Sader, S.; Ryan, R.E.; Smoot, J.; Kuper, P.; Ross, K.; Prados, D.; Russell, J.; Gasser, G.; McKellip, R.; et al. Assessment of MODIS NDVI time series data products for detecting forest defoliation by gypsy moth outbreaks. Remote Sens. Environ. 2011, 115, 427–437. [Google Scholar] [CrossRef]
- Olsson, P.O.; Lindström, J.; Eklundh, L. Near real-time monitoring of insect induced defoliation in subalpine birch forests with MODIS derived NDVI. Remote Sens. Environ. 2016, 181, 42–53. [Google Scholar] [CrossRef]
- Eklundh, L.; Johansson, T.; Solberg, S. Mapping insect defoliation in scots pine with MODIS time-series data. Remote Sens. Environ. 2009, 113, 1566–1573. [Google Scholar] [CrossRef]
- Hart, S.J.; Veblen, T.T. Detection of spruce beetle-induced tree mortality using high- and medium-resolution remotely sensed imagery. Remote Sens. Environ. 2015, 168, 134–145. [Google Scholar] [CrossRef]
- Bryk, M.; Kołodziej, B.; Pliszka, R. Changes of Norway spruce health in the Białowieża forest (CE Europe) in 2013–2019 during a bark beetle infestation, studied with Landsat imagery. Forests 2021, 12, 34. [Google Scholar] [CrossRef]
- de Beurs, K.M.; Townsend, P.A. Estimating the effect of gypsy moth defoliation using MODIS. Remote Sens. Environ. 2008, 112, 3983–3990. [Google Scholar] [CrossRef]
- Rullan-Silva, C.D.; Olthoff, A.E.; de la Mata, J.A.D.; Pajares-Alonso, J.A. Remote monitoring of forest insect defoliation. A review. For. Syst. 2013, 22, 377–391. [Google Scholar] [CrossRef]
- Hais, M.; Wild, J.; Berec, L.; Brůna, J.; Kennedy, R.; Braaten, J.; Brož, Z. Landsat Imagery Spectral Trajectories—Important Variables for Spatially Predicting the Risks of Bark Beetle Disturbance. Remote Sens. 2016, 8, 687. [Google Scholar] [CrossRef]
- Stych, P.; Lastovicka, J.; Hladky, R.; Paluba, D. Evaluation of the Influence of Disturbances on Forest Vegetation Using the Time Series of Landsat Data: A Comparison Study of the Low Tatras and Sumava National Parks. ISPRS Int. J. Geo-Inf. 2019, 8, 71. [Google Scholar] [CrossRef]
- Havašová, M.; Bucha, T.; Ferenčík, J.; Jakuš, R. Applicability of a vegetation indices-based method to map bark beetle outbreaks in the High Tatra Mountains. Ann. For. Res. 2015, 58, 295–310. [Google Scholar] [CrossRef]
- Latifi, H.; Fassnacht, F.E.; Schumann, B.; Dech, S. Object-based extraction of bark beetle (Ips typographus L.) infestations using multi-date LANDSAT and SPOT satellite imagery. Prog. Phys. Geogr. Earth Environ. 2014, 38, 755–785. [Google Scholar] [CrossRef]
- Abdullah, H.; Skidmore, A.K.; Darvishzadeh, R.; Heurich, M. Sentinel-2 accurately maps green-attack stage of European spruce bark beetle (Ips typographus, L.) compared with Landsat-8. Remote Sens. Ecol. Conserv. 2019, 5, 87–106. [Google Scholar] [CrossRef]
- Zimmermann, S.; Hoffmann, K. Evaluating the capabilities of Sentinel-2 data for large-area detection of bark beetle infestation in the Central German Uplands. J. Appl. Remote Sens. 2020, 14, 024515. [Google Scholar] [CrossRef]
- Zhan, Z.; Yu, L.; Li, Z.; Ren, L.; Gao, B.; Wang, L.; Luo, Y. Combining GF-2 and Sentinel-2 Images to Detect Tree Mortality Caused by Red Turpentine Beetle during the Early Outbreak Stage in North China. Forests 2020, 11, 172. [Google Scholar] [CrossRef]
- Immitzer, M.; Atzberger, C. Early Detection of Bark Beetle Infestation in Norway Spruce (Picea abies, L.) using WorldView-2 Data Photogramm. Fernerkund. Geoinformation 2014, 5, 351–367. [Google Scholar]
- Tane, Z.; Roberts, D.; Koltunov, A.; Sweeney, S.; Ramirez, C. A framework for detecting conifer mortality across an ecoregion using high spatial resolution spaceborne imaging spectroscopy. Remote Sens. Environ. 2018, 209, 195–210. [Google Scholar] [CrossRef]
- Abdullah, H.; Darvishzadeh, R.; Skidmore, A.K.; Groen, T.A.; Heurich, M. European spruce bark beetle (Ips typographus, L.) green attack affects foliar reflectance and biochemical properties. Int. J. Appl. Earth Obs. Geoinf. 2018, 64, 199–209. [Google Scholar] [CrossRef]
- Foster, A.C.; Walter, J.A.; Shugart, H.H.; Sibold, J.; Negron, J. Spectral evidence of early-stage spruce beetle infestation in Engelmann spruce. For. Ecol. Manag. 2017, 384, 347–357. [Google Scholar] [CrossRef]
- Lausch, A.; Heurich, M.; Gordalla, D.; Dobner, H.J.; Gwillym-Margianto, S.; Salbach, C. Forecasting potential bark beetle outbreaks based on spruce forest vitality using hyperspectral remote-sensing techniques at different scales. For. Ecol. Manag. 2013, 308, 76–89. [Google Scholar] [CrossRef]
- Bárta, V.; Lukeš, P.; Homolová, L. Early detection of bark beetle infestation in Norway spruce forests of Central Europe using Sentinel-2. Int. J. Appl. Earth Obs. Geoinf. 2021, 100, 102335. [Google Scholar] [CrossRef]
- Dash, J.P.; Watt, M.S.; Pearse, G.D.; Heaphy, M.; Dungey, H.S. Assessing very high-resolution UAV imagery for monitoring forest health during a simulated disease outbreak. ISPRS J. Photogramm. Remote Sens. 2017, 131, 1–14. [Google Scholar] [CrossRef]
- Näsi, R.; Honkavaara, E.; Blomqvist, M.; Lyytikäinen-Saarenmaa, P.; Hakala, T.; Viljanen, N.; Kantola, T.; Holopainen, M. Remote sensing of bark beetle damage in urban forests at individual tree level using a novel hyperspectral camera from UAV and aircraft. Urban For. Urban Green. 2018, 30, 72–83. [Google Scholar] [CrossRef]
- Klouček, T.; Komárek, J.; Surový, P.; Hrach, K.; Janata, P.; Vašíček, B. The Use of UAV Mounted Sensors for Precise Detection of Bark Beetle Infestation. Remote Sens. 2019, 11, 1561. [Google Scholar] [CrossRef]
- Prozorov, S.S. The silk moth in fir forests of Siberia. Proc. SibLTI. Krasn. 1952, 93–132. (In Russian) [Google Scholar]
- Boldaruyev, V.O. Population Dynamics of the Siberian Silk Moth and Its Parasites; Buryat Publishers: Ulan-Ude, Russia, 1969; p. 162. (In Russian) [Google Scholar]
- Epova, V.I.; Pleshanov, A.S. Zones of Severity of Phyllophagous Insects in Asian Russia; Nauka: Novosibirsk, Russia, 1995; p. 147. (In Russian) [Google Scholar]
- Kolomiyets, N.G. Parasites and Predators of the Siberian Silk Moth; Nauka: Novosibirsk, Russia, 1962; p. 172. (In Russian) [Google Scholar]
- Kondakov, Y.P. Siberian silk moth outbreaks. In Population Ecology of Forest Animals in Siberia; Nauka: Novosibirsk, Russia, 1974; pp. 206–265. (In Russian) [Google Scholar]
- Rozhkov, A.S. Siberian Silk Moth; Nauka: Moscow, Russia, 1963; p. 175. (In Russian) [Google Scholar]
- Rozhkov, A.S. Outbreak of the Siberian Silk Moth and Insect Control Measures; Nauka: Moscow, Russia, 1965; p. 178. (In Russian) [Google Scholar]
- Soukhovolsky, V.G.; Tarasova, O.V.; Kovalev, A.V. A modeling of critical events in forest insects populations. Rus. J. Gen. Biol. 2020, 81, 374–386. (In Russian) [Google Scholar]
- Yurchenko, G.I.; Turova, G.I. Siberian and White-Striped Silkworms in the Far East; DalNIILCH Puplishers: Habarovsk, Russia, 2007; p. 98. (In Russian) [Google Scholar]
- Flø, D.; Rafoss, T.; Wendell, M.; Sundheim, L. The Siberian moth (Dendrolimus sibiricus), a pest risk assessment for Norway. Forest Ecosyst. 2020, 7, 48. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Ranson, K.J.; Kozuhovskaya, A.G.; Kondakov, Y.P.; Pestunov, I.A. NOAA/AVHRR satellite detection of Siberian silkmoth outbreaks in eastern Siberia. Int. J. Remote Sens. 2004, 25, 5543–5555. [Google Scholar] [CrossRef]
- Kirichenko, N.I.; Baranchikov, Y.N.; Vidal, S. Performance of the potentially invasive Siberian moth Dendrolimus superans sibiricus on coniferous species in Europe. Agric. For. Entomol. 2009, 11, 247–254. [Google Scholar] [CrossRef]
- Möykkynen, T.; Pukkala, T. Modelling of the spread of a potential invasive pest, the Siberian moth (Dendrolimus sibiricus) in Europe. For. Ecosyst. 2014, 1, 10. [Google Scholar] [CrossRef]
- Pavlov, I.N.; Litovka, Y.A.; Golubev, D.V.; Astapenko, S.A.; Chromogin, P.V. New outbreak of Dendrolimus sibiricus tschetv. in Siberia (2012–2017): Monitoring, modeling and biological control. Contemp. Probl. Ecol. 2018, 11, 406–419. [Google Scholar] [CrossRef]
- Sultson, S.; Goroshko, A.; Verkhovets, S.; Mikhaylov, P.; Ivanov, V.; Demidko, D.; Kulakov, S. Orographic Factors as a Predictor of the Spread of the Siberian Silk Moth Outbreak in the Mountainous Southern Taiga Forests of Siberia. Land 2021, 10, 115. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Im, S.T.; Soldatov, V.V. Siberian silkmoth outbreaks surpassed geoclimatic barrier in Siberian Mountains. J. Mt. Sci. 2020, 17, 1891–1900. [Google Scholar] [CrossRef]
- NASA EOS (Earth Observing System), Appeears. Available online: https://appeears.earthdatacloud.nasa.gov/ (accessed on 14 July 2023).
- Sentinel EO-Browser. Available online: https://apps.sentinel-hub.com/eo-browser/ (accessed on 15 July 2023).
- Polikarpov, N.P.; Chebakova, N.M.; Nazimova, D.I. Climate and Mountain Forests of Southern Siberia; Nauka: Novosibirsk, Russia, 1986; p. 224. [Google Scholar]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with Erts. NASA Spec. Publ. 1974, 351, 538. [Google Scholar]
- McFeeters, S.K. The use of the Normalized Difference Water Index (NDWI) in the delineation of open water features. Int. J. Remote Sens. 1996, 17, 1425–1432. [Google Scholar] [CrossRef]
- Hunt, E.R.; Rock, B.N. Detection of changes in leaf water content using near- and middle-infrared reflectances. Remote Sens. Environ. 1989, 30, 43–54. [Google Scholar]
- Jiang, Z.; Huete, A.R.; Didan, K.; Miura, T. Development of a two-band enhanced vegetation index without a blue band. Remote Sens. Environ. 2008, 112, 3833–3845. [Google Scholar] [CrossRef]
- Xiaoyang, Z.; Jinfeng, L. The Derivation of a Reflectance Model for The Estimation of Leaf Area Index Using Perpendicular Vegetation Index. Remote Sens. Technol. Appl. 1995, 10, 13–18. [Google Scholar]
- Rondeaux, G.; Steven, M.; Baret, F. Optimization of soil-adjusted vegetation indices. Remote Sens. Environ. 1996, 55, 95–107. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Clevers, J.G.P.W. Application of a weighted infrared-red vegetation index for estimating leaf Area Index by Correcting for Soil Moisture. Remote Sens. Environ. 1989, 29, 25–37. [Google Scholar] [CrossRef]
- McDade, I.C.; Llewellyn, E.J.; Greer, R.G.H.; Murtagh, D.P. ETON 3: Altitude profiles of the nightglow continuum at green and near infrared wavelengths. Planet. Space Sci. 1986, 34, 801–810. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
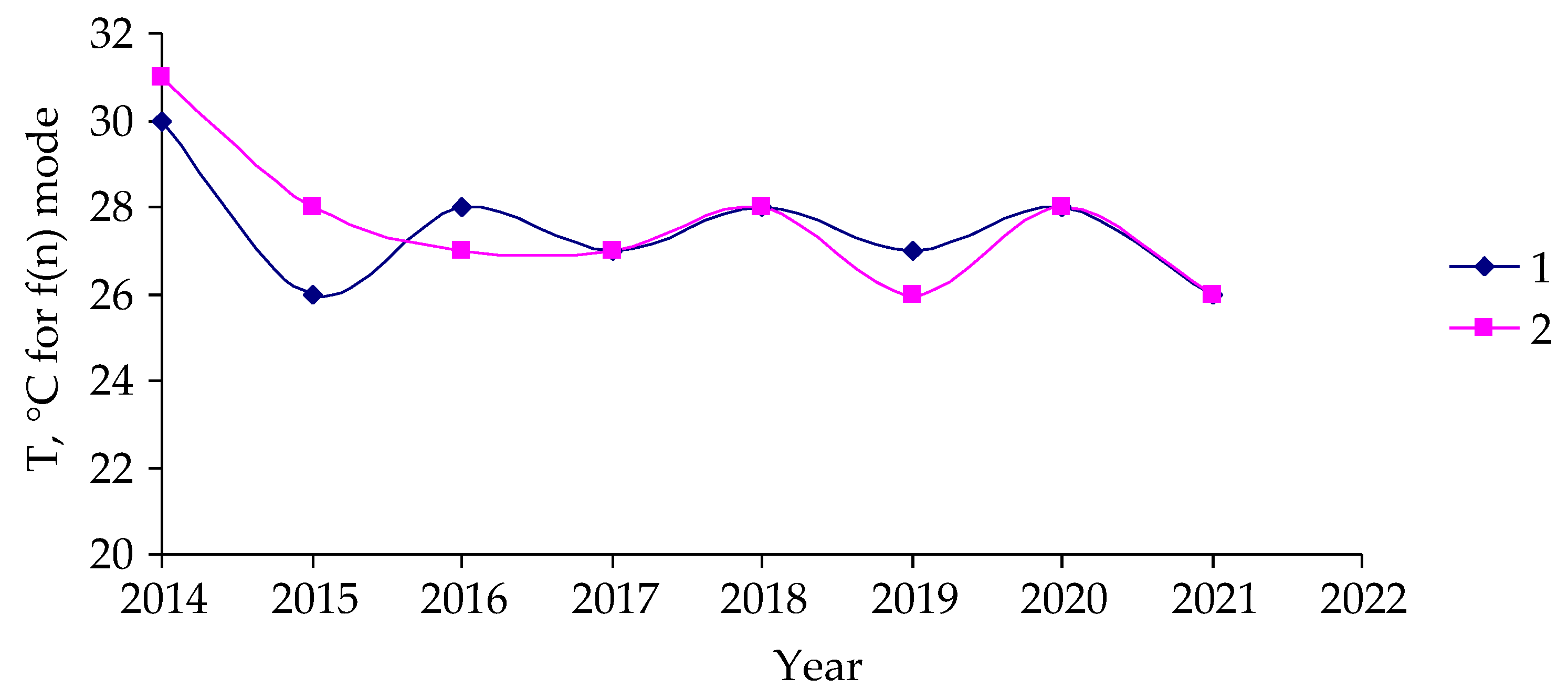{kind=link}
{kind=link}
{kind=link}
| Year | Proportion of Pixels with U = 0 | |
|---|---|---|
| Control Stands | 2019 Outbreak Zone | |
| 2014 | 0.52 | 0.51 |
| 2015 | 0.31 | 0.42 |
| 2016 | 0.58 | 0.70 |
| 2017 | 0.45 | 0.62 |
| 2018 | 0.40 | 0.64 |
| 2019 | 0.48 | 0.67 |
| 2020 | 0.24 | 0.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soukhovolsky, V.; Kovalev, A.; Goroshko, A.A.; Ivanova, Y.; Tarasova, O. Monitoring and Prediction of Siberian Silk Moth Dendrolimus sibiricus Tschetv. (Lepidoptera: Lasiocampidae) Outbreaks Using Remote Sensing Techniques. Insects 2023, 14, 955. https://doi.org/10.3390/insects14120955
Soukhovolsky V, Kovalev A, Goroshko AA, Ivanova Y, Tarasova O. Monitoring and Prediction of Siberian Silk Moth Dendrolimus sibiricus Tschetv. (Lepidoptera: Lasiocampidae) Outbreaks Using Remote Sensing Techniques. Insects. 2023; 14(12):955. https://doi.org/10.3390/insects14120955
Chicago/Turabian StyleSoukhovolsky, Vladislav, Anton Kovalev, Andrey A. Goroshko, Yulia Ivanova, and Olga Tarasova. 2023. "Monitoring and Prediction of Siberian Silk Moth Dendrolimus sibiricus Tschetv. (Lepidoptera: Lasiocampidae) Outbreaks Using Remote Sensing Techniques" Insects 14, no. 12: 955. https://doi.org/10.3390/insects14120955
APA StyleSoukhovolsky, V., Kovalev, A., Goroshko, A. A., Ivanova, Y., & Tarasova, O. (2023). Monitoring and Prediction of Siberian Silk Moth Dendrolimus sibiricus Tschetv. (Lepidoptera: Lasiocampidae) Outbreaks Using Remote Sensing Techniques. Insects, 14(12), 955. https://doi.org/10.3390/insects14120955