
Implications of the Niche Partitioning and Coexistence of Two Resident Parasitoids for Drosophila suzukii Management in Non-Crop Areas

 , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Drosophilid Puparia Sampling
2.3. Drosophilid Puparia Processing and Identification
2.4. Adult Parasitoid and Fly Identification
2.5. Data Analysis
3. Results
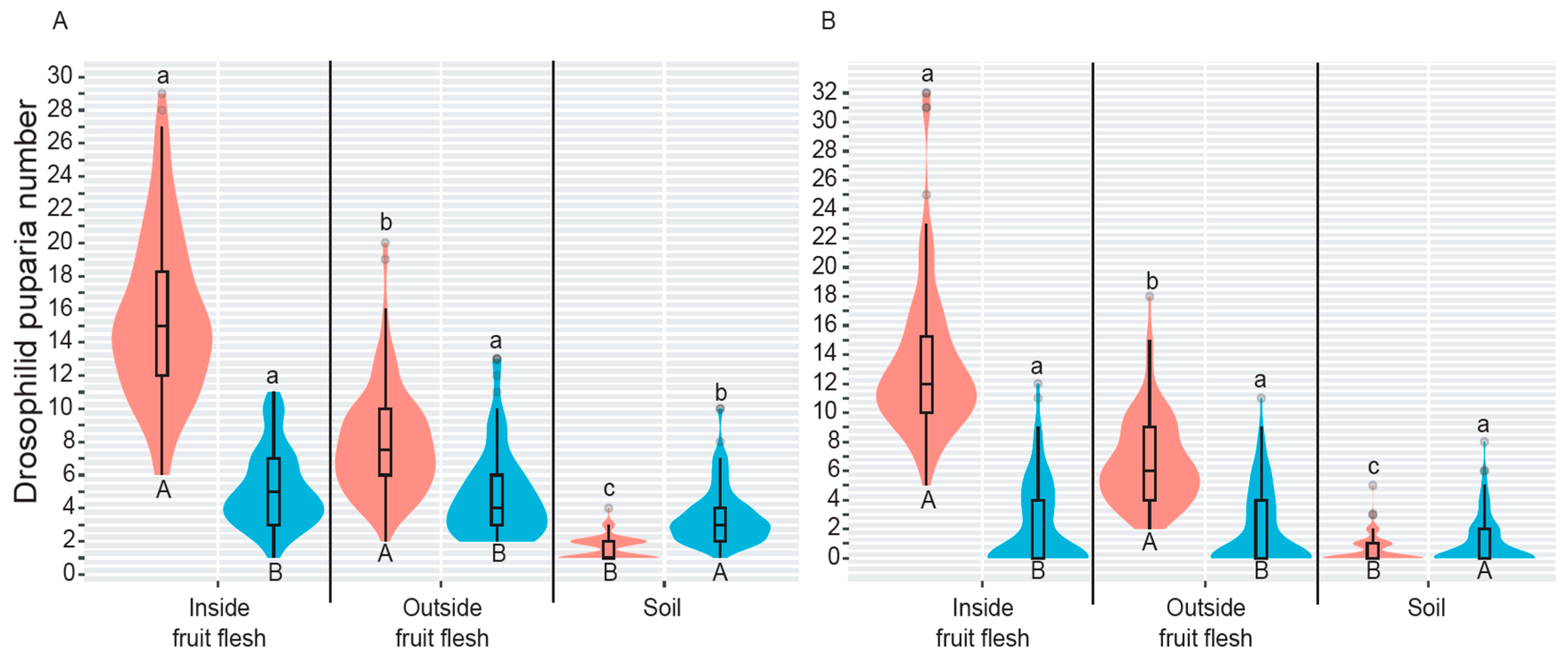
3.1. Drosophilid Fly Abundance and Relationship with Microhabitats Tested
3.2. Pupal Parasitoid Abundance and Relationship with Microhabitats Tested
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reitz, S.R.; Trumble, J.T. Competitive displacement among insects and arachnids. Annu. Rev. Entomol. 2002, 47, 435–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonsall, M.B.; Hassell, M.P.; Asefa, G. Ecological trade-offs, resource partitioning, and coexistence in a host–parasitoid assemblage. Ecology 2002, 83, 925–934. [Google Scholar]
- Amarasekare, P. Competitive coexistence in spatially structured environments: A synthesis. Ecol. Lett. 2003, 6, 1109–1122. [Google Scholar] [CrossRef]
- Wang, X.G.; Messing, R.H. Two different life history strategies determine the competitive outcome between Dirhinus giffardii and Pachycrepoideus vindemmiae, ectoparasitoids of cyclorrhaphous Diptera. Bull. Entomol. Res. 2004, 94, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pekas, A.; Tena, A.; Harvey, J.A.; Garcia-Marí, F.; Frag, F. Host size and spatiotemporal patterns mediate the coexistence of specialist parasitoids. Ecology 2016, 97, 1345–1356. [Google Scholar] [CrossRef] [PubMed]
- Aluja, M.; Ovruski, S.M.; Sivinski, J.; Córdova-García, G.; Schliserman, P.; Campero, S.R.N.; Ordano, M. Inter-specific competition and competition free space in the tephritid parasitoids Utetes anastrephae and Doryctobracon areolatus (Hymenoptera: Braconidae: Opiinae). Ecol. Entomol. 2013, 38, 485–496. [Google Scholar] [CrossRef]
- Tscharntke, T. Coexistence, tritrophic interactions and density dependence in a species-rich parasitoid community. J. Anim. Ecol. 1992, 61, 59–67. [Google Scholar] [CrossRef]
- Pedersen, B.S.; Mills, N.J. Single vs. multiple introduction in biological control: The roles of parasitoid efficiency, antagonism and niche overlap. J. Appl. Ecol. 2004, 41, 973–984. [Google Scholar] [CrossRef]
- Duan, J.J.; Van Driesche, R.G.; Schmude, J.M.; Quinn, N.F.; Petrice, T.R.; Rutledge, C.E.; Poland, T.M.; Bauer, L.S.; Elkinton, J.S. Niche partitioning and coexistence of parasitoids of the same feeding guild introduced for biological control of an invasive forest pest. Biol. Control 2021, 160, 104698. [Google Scholar] [CrossRef]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted-wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Garcia, F.R.M.; Lasa, R.; Funes, C.F.; Buzzetti, K. Drosophila suzukii Management in Latin America: Current Status and Perspectives. J. Econ. Entomol. 2022, 115, 1008–1023. [Google Scholar] [CrossRef]
- Escobar, L.I.; Ovruski, S.M.; Kirschbaum, D.S. Foreign invasive pests Drosophila suzukii (Matsumura) and Zaprionus indianus Gupta (Diptera: Drosophilidae) threaten fruit production in northwestern Argentina. Dros. Info. Serv. 2018, 101, 9–14. [Google Scholar]
- De Ros, G.; Grassi, A.; Pantezzi, T. Recent Trends in the Economic Impact of Drosophila suzukii. In Drosophila Suzukii Management; Garcia, F.R.M., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 11–28. [Google Scholar]
- Atallah, J.; Teixeira, L.; Salazar, R.; Zaragoza, G.; Kopp, A. The making of a pest: The evolution of a fruit-penetrating ovipositor in Drosophila suzukii and related species. Proc. Royal Soc. B 2014, 281, 20132840. [Google Scholar] [CrossRef] [Green Version]
- Kirschbaum, D.S.; Funes, C.F.; Buonocore-Biancheri, M.J.; Suárez, L.; Ovruski, S.M. The biology and ecology of Drosophila suzukii (Diptera: Drosophilidae) (Chapter 4). In Drosophila Suzukii Management; Garcia, F.R.M., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 41–91. [Google Scholar]
- Biancheri, M.J.B.; Suárez, L.; Kirschbaum, D.S.; Garcia, F.R.M.; Funes, C.F.; Ovruski, S.M. Natural parasitism influences biological control strategies against both global invasive pests Ceratitis capitata (Diptera: Tephritidae) and Drosophila suzukii (Diptera: Drosophilidae), and the Neotropical-native pest Anastrepha fraterculus (Diptera: Tephritidae). Environ. Entomol. 2022, 51, 1120–1135. [Google Scholar]
- Sassù, F.; Nikolouli, K.; Stauffer, C.; Bourtzis, K.; Cáceres, C. Insect technique and incompatible insect technique for the integrated Drosophila suzukii management (Chapter 9). In Drosophila Suzukii Management; Garcia, F.R.M., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 169–194. [Google Scholar]
- Lee, J.C.; Dreves, A.J.; Cave, A.M.; Kawai, S.; Isaacs, R.; Miller, J.C.; Van Timmeren, S.; Bruck, D.J. Infestation of wild and ornamental noncrop fruits by Drosophila suzukii (Diptera: Drosophilidae). Ann. Entomol. Soc. Am. 2015, 108, 117–129. [Google Scholar] [CrossRef]
- Kenis, M.; Tonina, L.; Eschen, R.; van der Sluis, B.; Sancassani, M.; Mori, N.; Haye, T.; Helsen, H. Non-crop plants used as hosts by Drosophila suzukii in Europe. J. Pest Sci. 2016, 89, 735–748. [Google Scholar] [CrossRef] [Green Version]
- Rossi Stacconi, M.V.; Grassi, A.; Ioriatti, C.; Anfora, G. Augmentative releases of Trichopria drosophilae for the suppression of early season Drosophila suzukii populations. BioControl 2019, 64, 9–19. [Google Scholar] [CrossRef]
- Garcia, F.R.M. Basis for area-wide management of Drosophila suzukii in Latin America (Chapter 5). In Drosophila Suzukii Management; Garcia, F.R.M., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 93–110. [Google Scholar]
- Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Lee, J.C. Biological control of spotted-wing drosophila: An update on promising agents (Chapter 8). In Drosophila Suzukii Management; Garcia, F.R.M., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 143–168. [Google Scholar]
- Abram, P.K.; Wang, X.G.; Hueppelsheuser, T.; Franklin, M.T.; Daane, K.M.; Lee, J.C.; Lue, C.H.; Girod, P.; Carrillo, J.; Wong, W.H.L.; et al. A coordinated sampling and identification methodology for larval parasitoids of spotted-wing drosophila. J. Econ. Entomol. 2022, 115, 922–942. [Google Scholar] [CrossRef]
- Garcia, F.R.M.; Corseuil, E. Native hymenopteran parasitoids associated with fruit flies (Diptera: Tephritoidea) in Santa Catarina State, Brazil. Fla. Entomol. 2004, 87, 517–521. [Google Scholar] [CrossRef]
- da Costa Oliveira, D.; Stupp, P.; Martins, L.N.; Wollmann, J.; Geisler, F.C.S.; Cardoso, T.D.N.; Bernardi, D.; Garcia, F.R.M. Interspecific competition in Trichopria anastrephae parasitism (Hymenoptera: Diapriidae) and Pachycrepoideus vindemmiae (Hymenoptera: Pteromalidae) parasitism on pupae of Drosophila suzukii (Diptera: Drosophilidae). Phytoparasitica 2021, 49, 207–215. [Google Scholar] [CrossRef]
- Wang, X.G.; Messing, R.H. The ectoparasitic pupal parasitoid, Pachycrepoideus vindemmiae (Hymenoptera: Pteromalidae), attacks other primary tephritid fruit fly parasitoids: Host expansion and non-target impact. Biol. Control 2004, 31, 227–236. [Google Scholar] [CrossRef]
- Garcia, F.R.M.; Ovruski, S.M.; Suárez, L.; Cancino, J.; Liburd, O.E. Biological control of tephritid fruit fies in the Americas and Hawaii: A review of the use of parasitoids and predators. Insects 2020, 11, 662. [Google Scholar] [CrossRef] [PubMed]
- Funes, C.F.; Garrido, S.A.; Aquino, D.A.; Escobar, L.I.; Gómez Segade, C.; Cichón, L.; Ovruski, S.M.; Kirschbaum, D.S. New records of Pachycrepoideus vindemmiae (Hymenoptera: Pteromalidae) associated with Drosophila suzukii (Diptera: Drosophilidae) in cherry and berry crops from Argentina. Rev. Soc. Entomol. Arg. 2020, 79, 39–43. [Google Scholar] [CrossRef]
- Ovruski, S.M.; Aluja, M.; Sivinski, J.; Wharton, R. Hymenopteran parasitoids on fruit infesting Tephritidae (Diptera) in Latin America and the Southern United States: Diversity, distribution, taxonomic status and their use in fruit fly biological control. Integr. Pest Manag. Rev. 2000, 5, 81–107. [Google Scholar] [CrossRef]
- Ehler, L.E. Parasitoid communities, parasitoid guilds, and biological control. In Parasitoid Community Ecology; Hawkins, B.A., Sheehan, W., Eds.; Oxford University Press: Oxford, UK, 1994; pp. 418–436. [Google Scholar]
- Wang, X.G.; Kaçar, G.; Biondi, A.; Daane, K.M. Foraging efficiency and outcomes of interactions of two pupal parasitoids attacking the invasive spotted wing drosophila. Biol. Control 2016, 96, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Hogg, B.N.; Lee, L.C.; Rogers, M.A.; Worth, L.; Nieto, D.J.; Stahl, J.M.; Daane, K.M. Releases of the parasitoid Pachycrepoideus vindemmiae for augmentative biological control of spotted wing drosophila, Drosophila suzukii. Biol. Control 2022, 168, 104865. [Google Scholar] [CrossRef]
- Krüger, A.P.; Scheunemann, T.; Vieira, J.G.A.; Morais, M.C.; Bernardi, D.; Nava, D.E.; Garcia, F.R.M. Effects of extrinsic, intraspecific competition and host deprivation on the biology of Trichopria anastrephae (Hymenoptera: Diapriidae) reared on Drosophila suzukii (Diptera: Drosophilidae). Neotrop. Entomol. 2019, 48, 957–965. [Google Scholar] [CrossRef]
- Vieira, J.G.A.; Krüger, A.P.; Scheuneumann, T.; Morais, M.C.; Speriogin, H.S.; Garcia, F.R.M.; Nava, D.E.; Bernardi, D. Some aspects of the biology of Trichopria anastrephae (Hymenoptera: Diapriidae), a resident parasitoid attacking Drosophila suzukii (Diptera: Drosophilidae) in Brazil. J. Econ. Entomol. 2019, 113, 81–87. [Google Scholar] [CrossRef]
- Wollmann, J.; Schlesener, D.C.H.; Ferreira, M.S.; Garcia, M.S.; Costa, V.A.; Garcia, F.R.M. Parasitoids of drosophilidae with potential for parasitism on Drosophila suzukii in Brazil. Dros. Info. Serv. 2016, 99, 38–42. [Google Scholar]
- Trivellone, V.; Meier, M.; Cara, C.; Pollini Paltrinieri, L.; Gugerli, F.; Moretti, M.; Wolf, S.; Collatz, J. Multiscale determinants drive parasitization of Drosophilidae by Hymenopteran parasitoids in agricultural landscapes. Insects 2020, 11, 334. [Google Scholar] [CrossRef]
- Brown, A.D. Las selvas pedemontanas de las Yungas: Manejo sustentable y conservación de la biodiversidad de un ecosistema prioritario del noroeste argentino. In Selva Pedemontana de las Yungas: Historia Natural, Ecología y Manejo de un Ecosistema en Peligro; Brown, A.D., Blendinger, P.G., Lomáscolo, T., García Bes, P., Eds.; Ediciones del Subtrópico: Yerba Buena, Argentina, 2009; pp. 13–36. [Google Scholar]
- Brown, A.D.; Grau, H.R.; Malizia, L.R.; Grau, A. Argentina. In Bosques Nublados del Geotrópico; Kappelle, M., Brown, A.D., Eds.; Instituto Nacional de Biodiversidad: San José, Costa Rica, 2001; pp. 623–659. [Google Scholar]
- Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef]
- Ranz, J.M.; Maurin, D.; Chan, Y.S.; von Grotthuss, M.; Hillier, L.D.W.; Roote, J.; Ashburner, M.; Bergman, C.M. Principles of Genome Evolution in the Drosophila melanogaster Species Group. PLoS Biol. 2007, 5, e152. [Google Scholar] [CrossRef]
- Gibson, G.A.P. Illustrated Key to the Native and Introduced Chalcidoid Parasitoids of Filth Flies in America North of Mexico (Hymenoptera: Chalcidoidea). Available online: http://res2.agr.ca/ecorc/apss/chalkey/keyintro.htm (accessed on 20 October 2000).
- Risbec, J. La Faune Entomologique des Cultures au Sénégal et au Soudan: II. Contribution à L’étude des Proctotrupidae (Serphiidae). [s.l.], (Travaux du Laboratoire d’Entomologie du Secteur Soudanais de Recherche Agronomique–Station de Bambey); Gouvernement Général de l’Afrique Occidentale Française, 1950; 638p. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R. Foundat Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 10 October 2022).
- Wobbrock, J.O.; Findlater, L.; Gergle, D.; Higgins, J.J. The Aligned Rank Transform for nonparametric factorial analyses using only Anova Procedures. In Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, Vancouver, BC, Canada, 7–12 May 2011. [Google Scholar] [CrossRef]
- Elkin, L.A.; Kay, M.; Higgins, J.J.; Wobbrock, J.O. An Aligned Rank Transform procedure for multifactor contrast tests. In Proceedings of the 34th Annual ACM Symposium on User Interface Software and Technology, 10–14 October 2021. [Google Scholar] [CrossRef]
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R. Package Version 1.3-5. 2021. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 10 October 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Woltz, J.M.; Lee, J.C. Pupation behavior and larval and pupal biocontrol of Drosophila suzukii in the field. Biol. Control 2017, 110, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C.; Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Isaacs, R.; Sial, A.A.; Walton, V.M. Biological control of spotted-wing drosophila (Diptera: Drosophilidae): Current and pending tactics. J. Integr. Pest Manag. 2019, 10, 13. [Google Scholar] [CrossRef]
- Kienzle, R.; Groß, L.B.; Caughman, S.; Rohlfs, M. Resource use by individual Drosophila suzukii reveals a flexible preference for oviposition into healthy fruits. Sci. Rep. 2020, 10, 3132. [Google Scholar] [CrossRef] [Green Version]
- Mazzetto, F.; Lessio, F.; Giacosa, S.; Rolle, L.; Alma, A. Relationships between Drosophila suzukii and grapevine in Northwestern Italy: Seasonal presence and cultivar susceptibility. Bull. Insectol. 2020, 73, 29–38. [Google Scholar]
- Scheidler, N.H.; Liu, C.; Hamby, K.A.; Zalom, F.G.; Syed, Z. Volatile codes: Correlation of olfactory signals and reception in Drosophila-yeast chemical communication. Sci. Rep. 2015, 5, 14059. [Google Scholar] [CrossRef] [Green Version]
- Schlesener, D.C.H.; Wollmann, J.J.; de Bastos Pazini, J.; Padilha, A.C.; Grützmacher, A.D.; Garcia, F.R.M. Insecticide Toxicity to Drosophila suzukii (Diptera: Drosophilidae) parasitoids: Trichopria anastrephae (Hymenoptera: Diapriidae) and Pachycrepoideus vindemmiae (Hymenoptera: Pteromalidae). J. Econ. Entomol. 2019, 112, 1197–1206. [Google Scholar] [CrossRef]
- Stacconi, M.V.R.; Panel, A.; Baser, N.; Ioriatti, C.; Pantezzi, T.; Anfora, G. Comparative life history traits of indigenous Italian parasitoids of Drosophila suzukii and their effectiveness at different temperatures. Biol. Control 2017, 112, 20–27. [Google Scholar] [CrossRef]
- Schliserman, P.; Aluja, M.; Rull, J.; Ovruski, S.M. Habitat degradation and introduction of exotic plants favor persistence of invasive species and population growth of native polyphagous fruit fly pests in a Northwestern Argentinean mosaic. Biol. Invasions 2014, 16, 2599–2613. [Google Scholar] [CrossRef]
- Schliserman, P.; Aluja, M.; Rull, J.; Ovruski, S.M. Temporal diversity and abundance patterns of parasitoids of fruit-infesting Tephritidae (Diptera) in the Argentinean Yungas: Implications for biological control. Environ. Entomol. 2016, 45, 1184–1198. [Google Scholar] [CrossRef] [PubMed]
- Daane, K.M.; Wang, X.G.; Biondi, A.; Miller, B.; Miller, J.C.; Riedl, H.; Shearer, P.W.; Guerrieri, E.; Giorgini, M.; Buffington, M. First exploration of parasitoids of Drosophila suzukii in South Korea as potential classical biological agents. J. Pest Sci. 2016, 89, 823–835. [Google Scholar] [CrossRef]
- Wang, X.G.; Kaçar, G.; Biondi, A.; Daane, K.M. Life-history and host preference of the pupal parasitoid Trichopria drosophilae of spotted wing drosophila. BioControl 2016, 61, 387–397. [Google Scholar] [CrossRef]
- Wolf, S.; Barmettler, E.; Eisenring, M.; Romeis, J.; Collatz, J. Host searching and host preference of resident pupal parasitoids of Drosophila suzukii in the invaded regions. Pest Manag. Sci. 2021, 77, 243–252. [Google Scholar] [CrossRef]
- Mazzetto, F.; Marchetti, E.; Amiresmaeili, N.; Sacco, D.; Francati, S.; Jucker, C.; Dindo, M.L.; Lupi, D.; Tavell, L. Drosophila parasitoids in northern Italy and their potential to attack the exotic pest Drosophila suzukii. J. Pest Sci. 2016, 89, 837–850. [Google Scholar] [CrossRef]
- Kaçar, G.; Wang, X.G.; Biondi, A.; Daane, K.M. Linear functional response by two pupal Drosophila parasitoids foraging within single or multiple patch environments. PLoS ONE 2017, 12, e0183525. [Google Scholar] [CrossRef] [Green Version]
- Rossi-Stacconi, M.V.; Amiresmaeili, N.; Biondi, A.; Carli, C.; Caruso, S.; Dindo, M.L.; Francati, S.; Gottardello, A.; Grassi, A.; Lupi, D.; et al. Host location and dispersal ability of the cosmopolitan parasitoid Trichopria drosophilae released to control the invasive spotted wing Drosophila. Biol. Control 2018, 117, 188–196. [Google Scholar] [CrossRef]
- Pfab, F.; Stacconi, M.V.R.; Anfora, G.; Grassi, A.; Walton, V.M.; Pugliese, A. Optimized timing of parasitoid release: A mathematical model for biological control of Drosophila suzukii. Theor. Ecol. 2018, 11, 450–489. [Google Scholar] [CrossRef]
- Wang, X.G.; Serrato, M.A.; Son, Y.; Walton, V.M.; Hogg, B.N.; Daane, K.M. Thermal performance of two indigenous pupal parasitoids attacking the invasive Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2018, 47, 764–772. [Google Scholar] [CrossRef] [Green Version]
- Yi, C.; Cai, P.; Lin, J.; Liu, X.; Ao, G.; Zhang, Q.; Xia, H.; Yang, J.; Ji, Q. Life History and Host Preference of Trichopria drosophilae from Southern China, One of the Effective Pupal Parasitoids on the Drosophila Species. Insects 2020, 11, 103. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Cabrera, J.; Moreno-Carrillo, G.; Sanchez-Gonzalez, J.A. Single and combined release of Trichopria drosophilae (Hymenoptera: Diapriidae) to control Drosophila suzukii (Diptera: Drosophilidae). Neotrop. Entomol. 2019, 48, 949–956. [Google Scholar] [CrossRef]
- Harvey, J.A.; Tanaka, T.; Poelman, E.H. Intrinsic inter- and intraspecific competition in parasitoid wasps. Annu. Rev. Entomol. 2013, 58, 333–351. [Google Scholar] [CrossRef] [Green Version]
- Ovruski, S.M.; Schliserman, P. Biological control of tephritid fruit flies in Argentina: Historical review, current status, and future trends for developing a parasitoid mass-release program. Insects 2012, 3, 870–888. [Google Scholar] [CrossRef] [Green Version]
- Kruitwagen, A.; Beukeboom, L.W.; Wertheim, B. Optimization of native biocontrol agents, with parasitoids of the invasive pest D. suzukii as an example. Evol. Appl. 2018, 11, 1473–1497. [Google Scholar] [CrossRef]
- Jarrett, B.J.M.; Linder, S.; Fanning, P.D.; Isaacs, R.; Szucs, M. Experimental adaptation of native parasitoids to the invasive insect pest, Drosophila suzukii. Biol. Control 2022, 167, 104843. [Google Scholar] [CrossRef]
- Vargas, G.; Rivera-Pedroza, L.-F.; Cracía, L.F.; Mundstock Jahnke, S. Conservation Biological Control as an Important Tool in the Neotropical Region. Neotrop. Entomol. 2022. [Google Scholar] [CrossRef]
- Wolf, S.; Boycheva-Woltering, S.; Romeis, J.; Collatz, J. Trichopria drosophilae parasitizes Drosophila suzukii in seven common non-crop fruits. J. Pest Sci. 2020, 93, 627–638. [Google Scholar] [CrossRef]
- Funes, C.F.; Kirschbaum, D.S.; Escobar, L.I.; Heredia, A.M. La Mosca de las alas Manchadas, Drosophila suzukii (Matsumura), Nueva Plaga de las Frutas Finas en Argentina; PDF, Digital Book; Ediciones INTA: Famaillá, Argentina, 2018; 28p. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microhabitat | Host Fruit | Statistical Analysis Outcome | ||
|---|---|---|---|---|
| df | Χ2 | p | ||
| Inside fruit flesh | Peach | 1 | 166.94 | <0.0001 * |
| Guava | 1 | 173.95 | <0.0001 * | |
| Outside fruit flesh | Peach | 1 | 66.78 | <0.0001 * |
| Guava | 1 | 108.91 | <0.0001 * | |
| Soil | Peach | 1 | 122.78 | <0.0001 * |
| Guava | 1 | 0.02 | =0.0270 * | |
| Host Fruit | Parasitoid Species | Source of Variation | Statistical Analysis Outcome | |||
|---|---|---|---|---|---|---|
| df | Residuals df | F | p | |||
| Peach | T. anastrephae | THU | 2 | 714 | 566.19 | <0.0001 * |
| PDS | 1 | 714 | 389.58 | <0.0001 * | ||
| THU × PDS | 2 | 714 | 204.23 | <0.0001 * | ||
| Guava | T. anastrephae | THU | 2 | 714 | 599.24 | <0.0001 * |
| PDS | 1 | 714 | 733.73 | <0.0001 * | ||
| THU × PDS | 2 | 714 | 298.26 | <0.0001 * | ||
| Peach | P. vindemiae | THU | 2 | 714 | 85.728 | <0.0001 * |
| PDS | 1 | 714 | 84.386 | <0.0001 * | ||
| THU × PDS | 2 | 714 | 113.08 | <0.0001 * | ||
| Guava | P. vindemiae | THU | 2 | 714 | 52.673 | <0.0001 * |
| PDS | 1 | 714 | 102.91 | <0.0001 * | ||
| THU × PDS | 2 | 714 | 42.630 | <0.0001 * | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buonocore Biancheri, M.J.; Núñez-Campero, S.R.; Suárez, L.; Ponssa, M.D.; Kirschbaum, D.S.; Garcia, F.R.M.; Ovruski, S.M. Implications of the Niche Partitioning and Coexistence of Two Resident Parasitoids for Drosophila suzukii Management in Non-Crop Areas. Insects 2023, 14, 222. https://doi.org/10.3390/insects14030222
Buonocore Biancheri MJ, Núñez-Campero SR, Suárez L, Ponssa MD, Kirschbaum DS, Garcia FRM, Ovruski SM. Implications of the Niche Partitioning and Coexistence of Two Resident Parasitoids for Drosophila suzukii Management in Non-Crop Areas. Insects. 2023; 14(3):222. https://doi.org/10.3390/insects14030222
Chicago/Turabian StyleBuonocore Biancheri, María Josefina, Segundo Ricardo Núñez-Campero, Lorena Suárez, Marcos Darío Ponssa, Daniel Santiago Kirschbaum, Flávio Roberto Mello Garcia, and Sergio Marcelo Ovruski. 2023. "Implications of the Niche Partitioning and Coexistence of Two Resident Parasitoids for Drosophila suzukii Management in Non-Crop Areas" Insects 14, no. 3: 222. https://doi.org/10.3390/insects14030222
APA StyleBuonocore Biancheri, M. J., Núñez-Campero, S. R., Suárez, L., Ponssa, M. D., Kirschbaum, D. S., Garcia, F. R. M., & Ovruski, S. M. (2023). Implications of the Niche Partitioning and Coexistence of Two Resident Parasitoids for Drosophila suzukii Management in Non-Crop Areas. Insects, 14(3), 222. https://doi.org/10.3390/insects14030222