

Stage Transitions in Lucilia sericata and Phormia regina (Diptera: Calliphoridae) and Implications for Forensic Science


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Flies
2.2. Incubators
2.3. Experimental Design
2.4. Analysis
- A Gaussian equation (a standard normal curve):
- 2.
- A modified Gaussian equation (a form of Gaussian curve with a plateau at 100%):
- 3.
- A cumulative Gaussian equation (a form of the Gaussian curve used for adults to model a sigmoidal increase to a plateau):
- 4.
- A reversed cumulative Gaussian equation (a form of the cumulative Gaussian equation used for eggs, to model a sigmoidal decrease from a plateau):
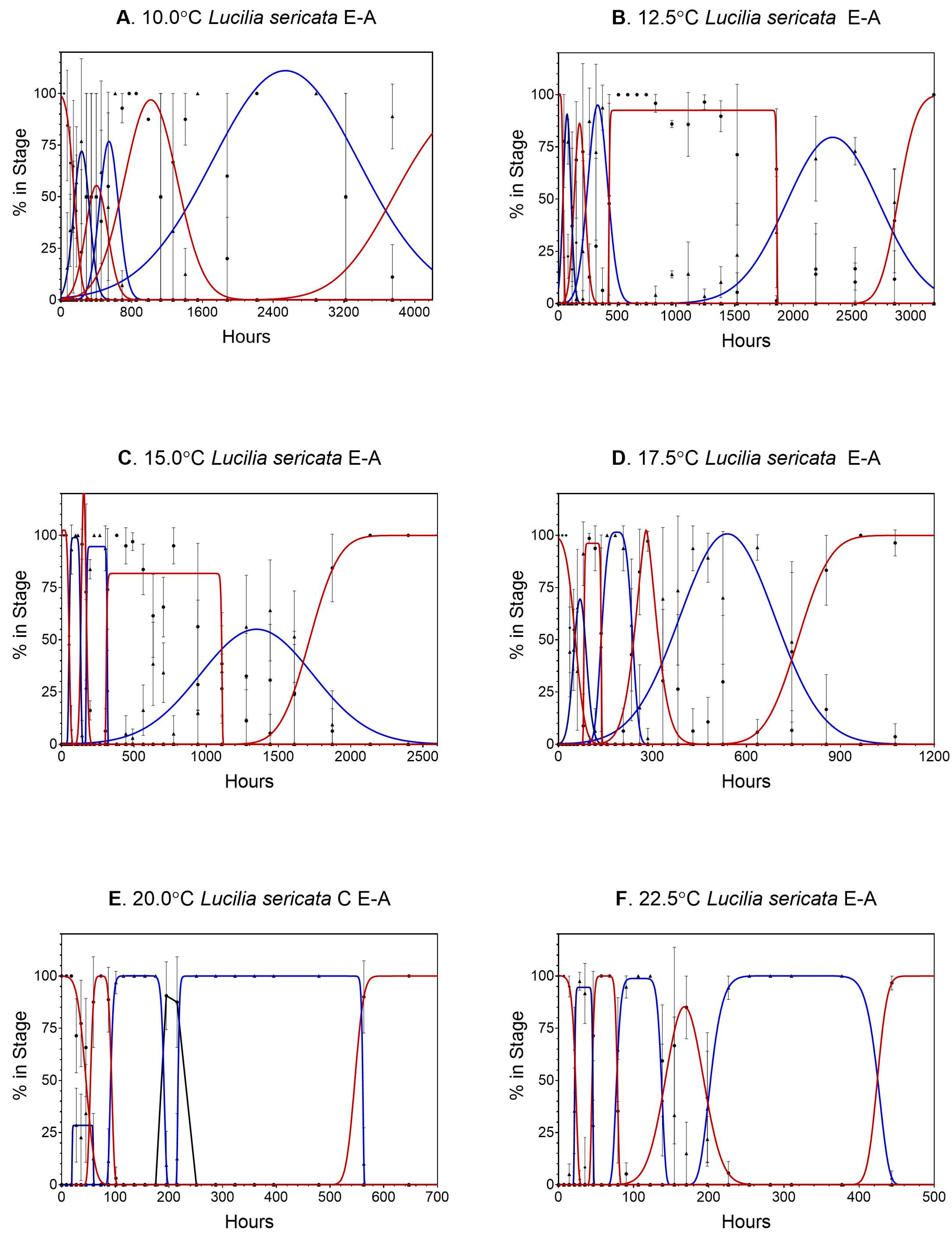
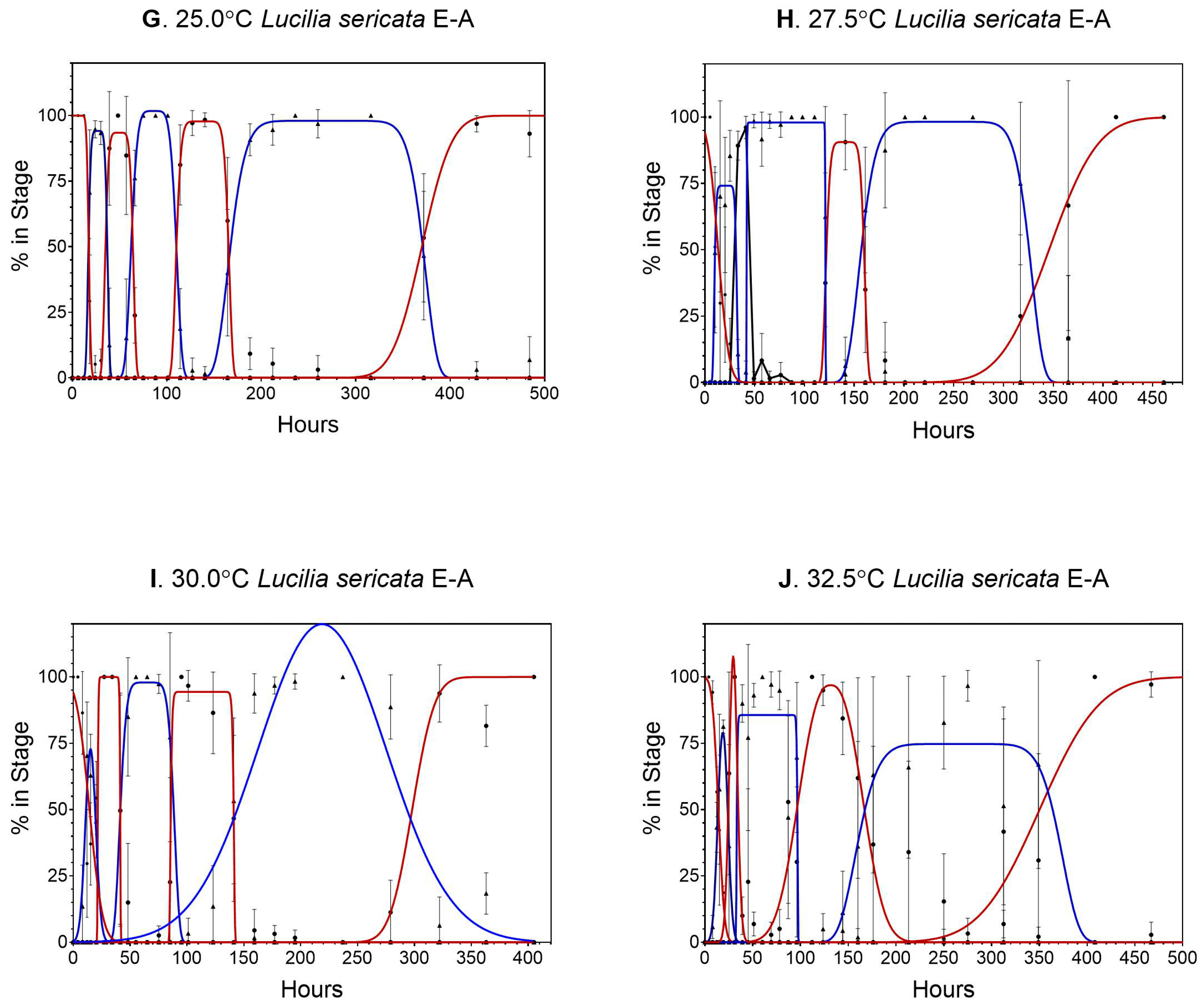
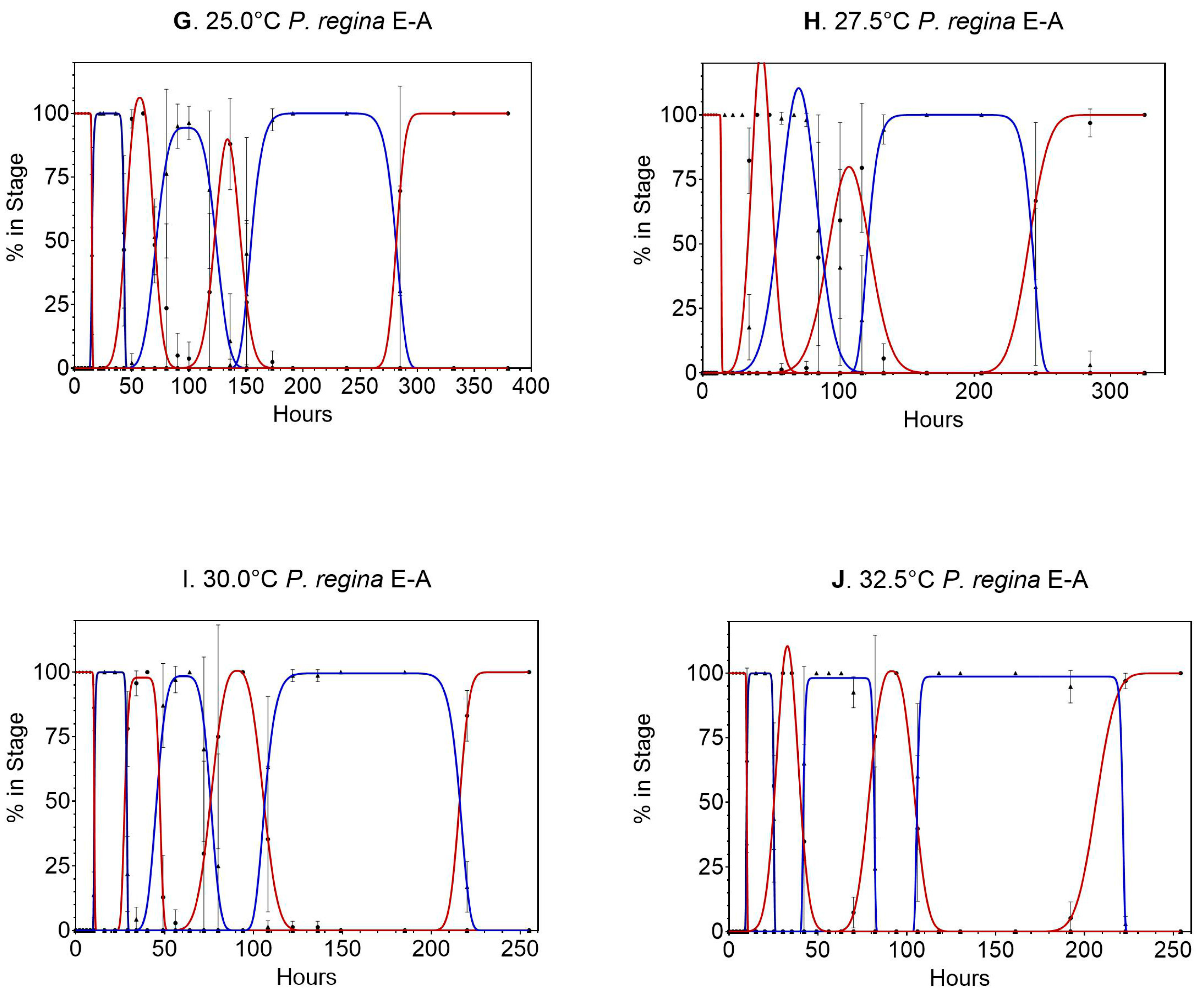
3. Results
3.1. Lucilia sericata
3.2. Phormia regina
4. Discussion
4.1. Lucilia sericata
4.2. Phormia regina
4.3. Forensic Implications of Findings
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Higley, L.G.; Haskell, N.H. Insect development and forensic entomology. In Forensic Entomology: The Utility of Arthropods in Legal Investigations; Byrd, J.H., Castner, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 389–405. [Google Scholar]
- Greenberg, B. Flies as forensic indicators. J. Med. Entomol. 1991, 28, 565–577. [Google Scholar] [CrossRef]
- Rivers, D.B.; Dahlem, G.A. The Science of Forensic Entomology; Wiley: Oxford, UK, 2014; pp. 131–149. [Google Scholar]
- Hall, D.G. The Blowflies of North America; Thomas Say Foundation: Annapolis, MD, USA, 1948. [Google Scholar]
- Wells, J.D.; LaMotte, L.R. Estimating postmortem interval. In Forensic Entomology: The Utility of Arthropods in Legal Investigations; Byrd, J.H., Castner, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 367–388. [Google Scholar]
- Wells, J.D.; Kurahashi, H. Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae) development: Rate, variation and the implications for forensic entomology. Japan. J. Sanit. Zool. 1994, 45, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Kamal, A.S. Comparative study of thirteen species of sarcosaprophagous Calliphoridae and Sarcophagidae (Diptera) I. Bionomics. Ann. Entomol. Soc. Am. 1958, 51, 261–271. [Google Scholar] [CrossRef]
- Richards, C.S.; Villet, M.H. Factors affecting accuracy and precision of thermal summation models of insect development used to estimate post-mortem intervals. Int. J. Leg. Med. 2008, 122, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Ash, N.; Greenberg, B. Developmental temperature responses of the sibling species Phaenicia sericata and Phaenicia pallescens. Ann. Entomol. Soc. Am. 1975, 68, 197–200. [Google Scholar] [CrossRef]
- Anderson, G.S. Minimum and maximum developmental rates of some forensically important Calliphoridae (Diptera). J. Forensic Sci. 2000, 45, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Byrd, J.H.; Allen, J.C. The development of the black blow fly, Phormia regina (Meigen). Forensic Sci. Inter. 2001, 120, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Byrd, J.H.; Castner, J.L. Insects of forensic importance. In Forensic Entomology: The Utility of Arthropods in Legal Investigations, 2nd ed.; Byrd, J.H., Castner, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 39–126. [Google Scholar]
- Nabity, P.D.; Higley, L.G.; Heng-Moss, T.M. Effects of temperature on development of Phormia regina (Diptera: Calliphoridae) and use of development data in determining time intervals in forensic entomology. J. Med. Entomol. 2006, 43, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Roe, A.; Higley, L.G. Development modeling of Lucilia sericata (Diptera: Calliphoridae). PeerJ 2015, 3, e803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Authement, M.L.; Higley, L.G.; Hoback, W.W. Anoxia tolerance in four forensically important calliphorid species. Forensic Sci. 2023, 3, 1. [Google Scholar] [CrossRef]
- Nabity, P.; Higley, L.; Heng-Moss, T. Light-induced variability in development of forensically important blow fly Phormia regina (Diptera: Calliphoridae). J. Med. Entomol. 2007, 44, 351–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, L.; Ratcliffe, G.G. Development rates of some pre-adult stages in blowflies with reference to low temperatures. Med. Vet. Entomol. 1994, 8, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Grassberger, M.; Reiter, C. Effect of temperature on Lucilia sericata (Diptera: Calliphoridae) development with special reference to the isomegalen- and isomorphen-diagram. Forensic Sci. Int. 2001, 120, 32–36. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature °C | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Life Stage | 7.5 | 10.0 | 12.5 | 15.0 | 17.5 | 20.0 | 22.5 | 25.0 | 27.5 | 30.0 | 32.5 |
| Egg–1st | 35 | 35 | 35 | 17 | 12 | 9 | 7 | 6 | 5 | 4 | 4 |
| 1st–2nd | 56 | 56 | 56 | 28 | 19 | 14 | 11 | 9 | 8 | 7 | 6 |
| 2nd–3f | 79 | 79 | 79 | 39 | 26 | 20 | 16 | 13 | 11 | 10 | 9 |
| 3f–3m | 143 | 143 | 143 | 71 | 48 | 36 | 29 | 24 | 20 | 18 | 16 |
| 3m–Pupal | 335 | 335 | 335 | 167 | 112 | 84 | 67 | 56 | 48 | 42 | 37 |
| Pupal–Adult | 527 | 527 | 527 | 263 | 176 | 132 | 105 | 88 | 75 | 66 | 59 |
| Temperature (°C) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Life Stage | 7.5 | 10.0 | 12.5 | 15.0 | 17.5 | 20.0 | 22.5 | 25.0 | 27.5 | 30.0 | 32.5 |
| Egg–1st | 16 | 16 | 16 | 8 | 5 | 4 | 3 | 3 | 2 | 2 | 2 |
| 1st–2nd | 44 | 44 | 44 | 22 | 15 | 11 | 9 | 7 | 6 | 6 | 5 |
| 2nd–3f | 63 | 63 | 63 | 31 | 21 | 16 | 13 | 10 | 9 | 8 | 7 |
| 3f–3m | 111 | 111 | 111 | 55 | 37 | 28 | 22 | 18 | 16 | 14 | 12 |
| 3m–Pupal | 281 | 281 | 281 | 141 | 94 | 70 | 56 | 47 | 40 | 35 | 31 |
| Pupal–Adult | 441 | 441 | 441 | 221 | 147 | 110 | 88 | 74 | 63 | 55 | 49 |
| Temperature °C | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Life Stage | 7.5 | 10.0 | 12.5 | 15.0 | 17.5 | 20.0 | 22.5 | 25.0 | 27.5 | 30.0 | 32.5 |
| Egg–1st | N/A | 186 | 41 | 52 | 47 | 28 | 21 | 17 | 18 | 12 | 14 |
| 1st–2nd | N/A | 407 | 136 | 131 | 79 | 59 | 46 | 36 | 31 | 24 | 24 |
| 2nd–3f | N/A | 463 | 231 | 179 | 137 | 92 | 78 | 63 | 49 | 42 | 36 |
| 3f–3m | N/A | 683 | 431 | 314 | 237 | 193 | 141 | 111 | 124 | 85 | 95 |
| 3m–Pupal | N/A | 2179 | 1935 | 1206 | 326 | 217 | 202 | 168 | 158 | 139 | 180 |
| Pupal–Adult | N/A | 4011 | 2896 | 1710 | 760 | 554 | 424 | 371 | 344 | 297 | 353 |
| Temperature (°C) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Life Stage | 7.5 | 10.0 | 12.5 | 15.0 | 17.5 | 20.0 | 22.5 | 25.0 | 27.5 | 30.0 | 32.5 |
| Egg–1st | N/A | 173 | 70 | 61 | 40 | 24 | 16 | 15 | 14 | 10 | 10 |
| 1st–2nd | N/A | N/A | 354 | 154 | 127 | 75 | 50 | 43 | 34 | 27 | 25 |
| 2nd–3f | N/A | N/A | 564 | 231 | 218 | 123 | 92 | 71 | 57 | 47 | 42 |
| 3f–3m | N/A | N/A | 612 | 521 | 277 | 202 | 134 | 123 | 95 | 76 | 78 |
| 3m–Pupal | N/A | N/A | 843 | 469 | 319 | 245 | 178 | 155 | 122 | 107 | 106 |
| Pupal–Adult | N/A | N/A | 1450 | 966 | 622 | 447 | 356 | 283 | 239 | 218 | 206 |
| Temp | Egg | L1 | L2 | L3f | L3m | P | |
|---|---|---|---|---|---|---|---|
| R and H | 27.5 | 5.2% | 3.7% | 5.4% | 22.0% | 9.7% | 54.1% |
| Kamal | 26.7 | 5.2% | 5.7% | 3.4% | 11.5% | 25.9% | 48.3% |
| Difference | 0.0% | −2.1% | 1.9% | 10.5% | −16.2% | 5.8% | |
| Transition Stages | Mean of Transition Time in Hours | |||||
|---|---|---|---|---|---|---|
| A & G 1975 | R & H | A & G 1975 | R & H | A & G 1975 | R & H | |
| 19.0 C | 20.0 C | 27.0 C | 27.5 C | 35.0 C | 32.5 C | |
| E-L1 | 29.4 | 28.1 | 14.4 | 17.9 | 10.2 | 13.8 |
| L3m-P | 691.2 | 216.8 | 194.4 | 157.6 | 333.6 | 180.0 |
| P-A | 1312.8 | 553.7 | 384.0 | 343.5 | 295.2 | 352.5 |
| Transition Stages | Difference in Transition Means (A&G (1975)—Presented Data) as Proportion of A&G Standard Deviations | ||
|---|---|---|---|
| 19 and 20 C | 27 and 27.5 C | 35 and 32.5 C | |
| E-L1 | 0.6 | 2.7 | 1.9 |
| L3m-P | 3.9 | 0.5 | 0.9 |
| P-A | 2.5 | 0.3 | 1.0 |
| E-L1 | E-L2 | E-L3f | E-L3m | E-P | E-A | ||
|---|---|---|---|---|---|---|---|
| R and H | mean | 13.8 | 33.9 | 56.6 | 95.3 | 122.1 | 239.2 |
| Kamal | mode | 16 | 34 | 45 | 81 | 165 | 309 |
| Difference | −2.2 | −0.1 | 11.6 | 14.3 | −42.9 | −69.8 | |
| Temp | Egg | L1 | L2 | L3f | L3m | P | |
|---|---|---|---|---|---|---|---|
| R and H | 27.5 | 6.1% | 8.9% | 10.1% | 17.2% | 11.9% | 52.0% |
| Kamal | 26.7 | 5.2% | 5.8% | 3.6% | 11.7% | 27.2% | 46.6% |
| Difference | 0.9% | −3.1% | 6.5% | 5.5% | −15.3% | 5.4% | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roe, A.L.; Higley, L.G. Stage Transitions in Lucilia sericata and Phormia regina (Diptera: Calliphoridae) and Implications for Forensic Science. Insects 2023, 14, 315. https://doi.org/10.3390/insects14040315
Roe AL, Higley LG. Stage Transitions in Lucilia sericata and Phormia regina (Diptera: Calliphoridae) and Implications for Forensic Science. Insects. 2023; 14(4):315. https://doi.org/10.3390/insects14040315
Chicago/Turabian StyleRoe, Amanda L., and Leon G. Higley. 2023. "Stage Transitions in Lucilia sericata and Phormia regina (Diptera: Calliphoridae) and Implications for Forensic Science" Insects 14, no. 4: 315. https://doi.org/10.3390/insects14040315
APA StyleRoe, A. L., & Higley, L. G. (2023). Stage Transitions in Lucilia sericata and Phormia regina (Diptera: Calliphoridae) and Implications for Forensic Science. Insects, 14(4), 315. https://doi.org/10.3390/insects14040315