

Host Preferences Shown by Drosophilids (Diptera) in a Commercial Fruit and Vegetable Distribution Center Follow the Wild Neotropical Pattern


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collections and Taxonomic Determination
2.2. Data Analyses
3. Results
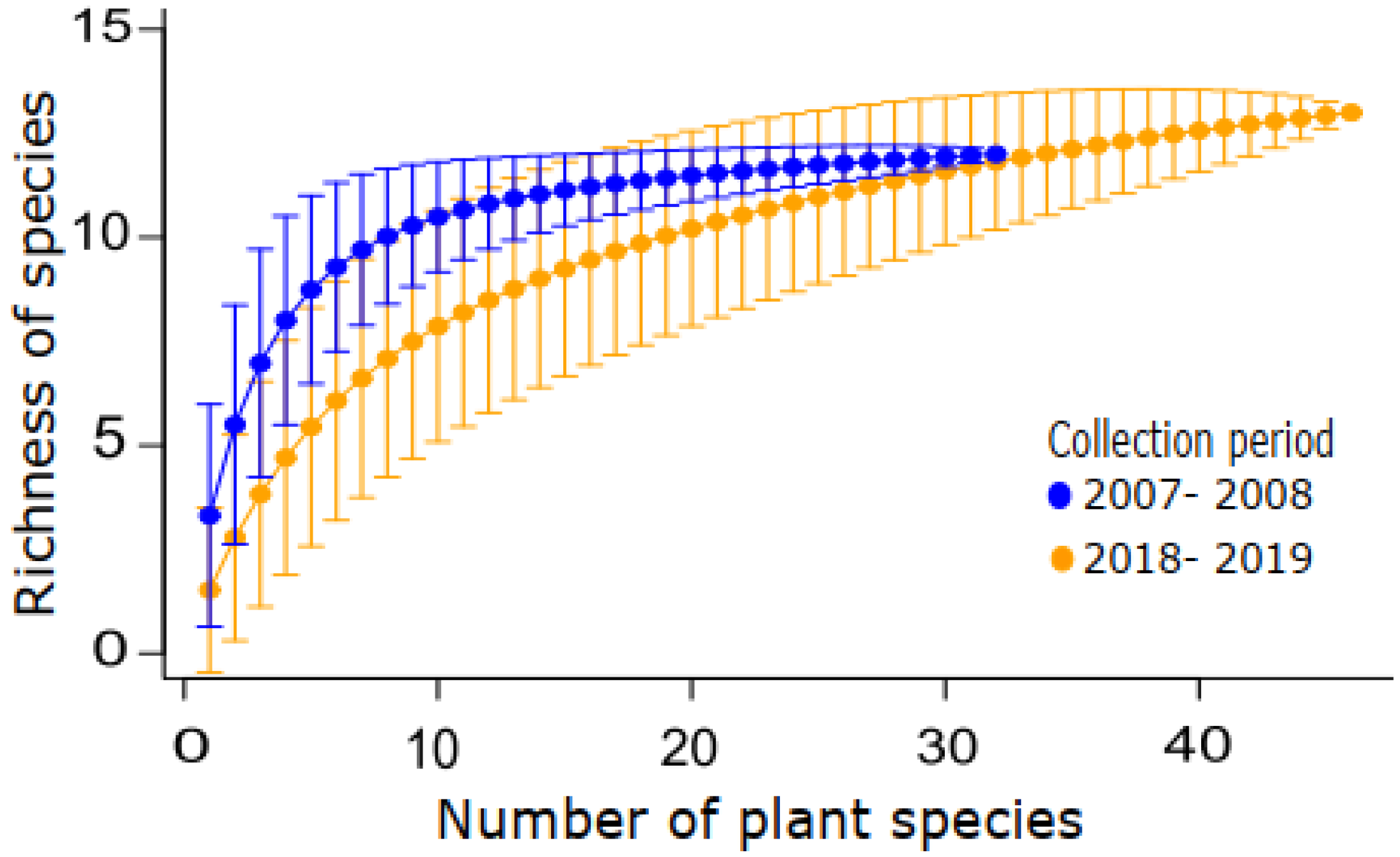
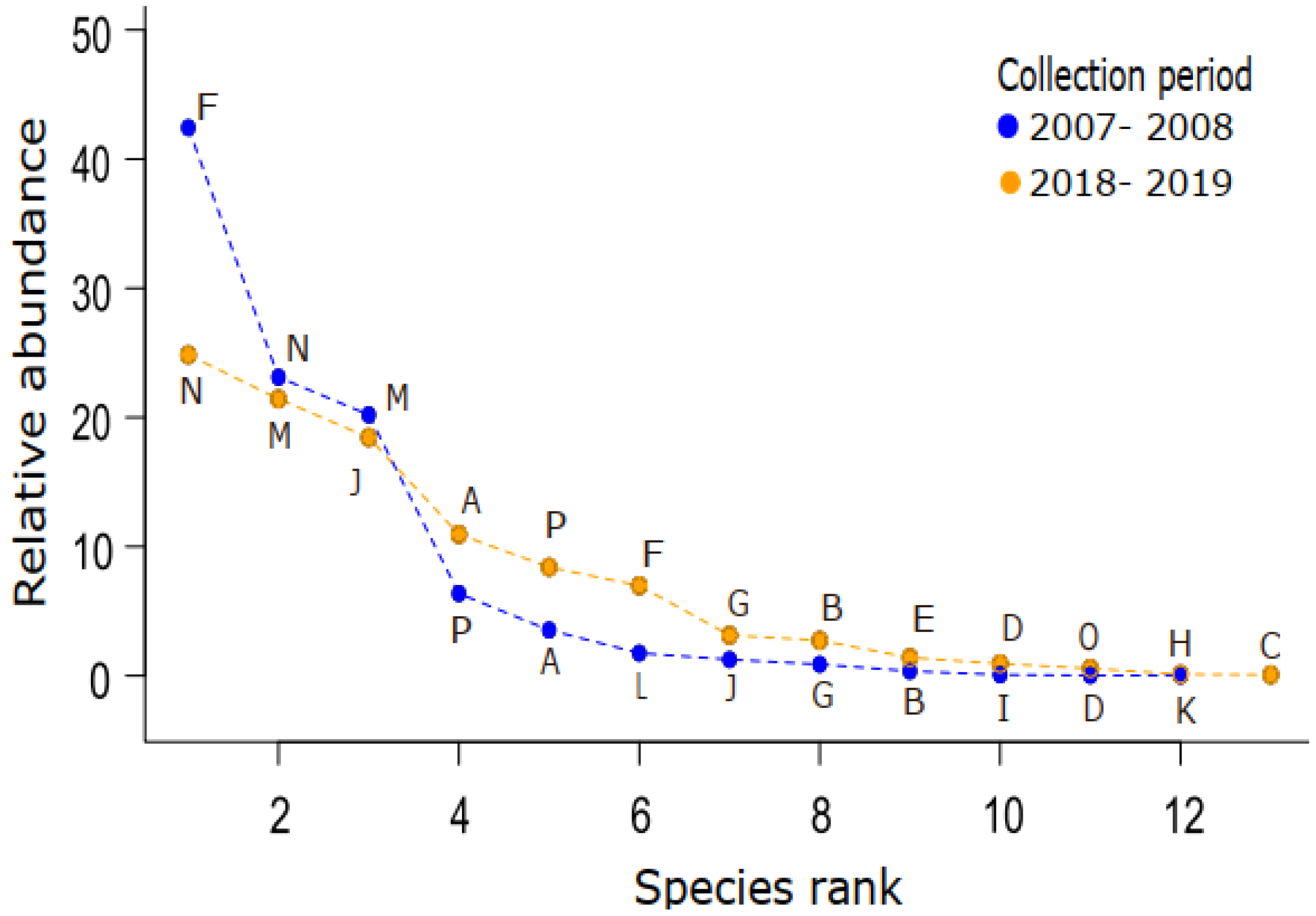
3.1. Temporal Changes
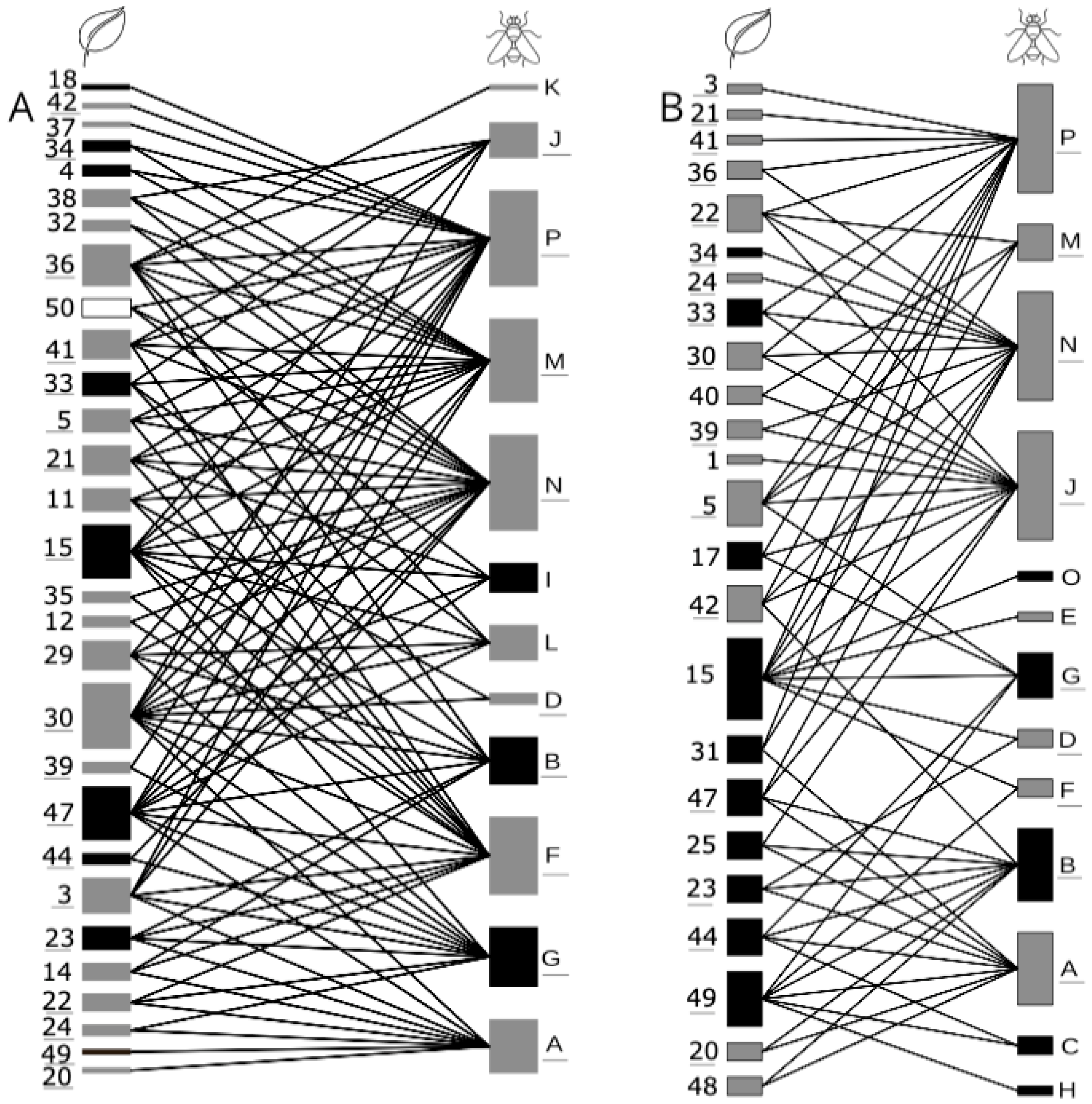
3.2. Relationships between Plant and Drosophilid Species
3.3. Neotropical and Exotic Resources Explored by Neotropical and Exotic Drosophilids
4. Discussion
4.1. The Fly Community Remained Relatively Stable after Ten Years
4.2. Drosophilid Species Are Not Randomly Distributed among Plant Species
4.3. Exotic Drosophilids Use More Resources Than Neotropical Drosophilids
4.4. Future Research
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- TaxoDros: The Database on Taxonomy of Drosophilidae, v.1.04. Database. Available online: https://www.taxodros.uzh.ch/search/class.php (accessed on 18 January 2023).
- Carson, H.L. The Ecology of Drosophila Breeding Sites; Harold L-Lyon Arboretum Lecture; University of Hawaii Foundation Lyon Arboretum Fund: New York, NY, USA, 1971; p. 28. [Google Scholar]
- David, J.R.; Tsacas, L. Cosmopolitan, subcosmopolitan and widespread species: Different strategies within the Drosophilid family (Diptera). C. R. Soc. Biogeogr. 1981, 57, 11–26. [Google Scholar]
- Drosophilidae in Catálogo Taxonômico da Fauna do Brasil. PNUD. Available online: http://fauna.jbrj.gov.br/fauna/faunadobrasil/183186 (accessed on 19 January 2023).
- Val, F.C.; Sene, F.M. Nova Introdução de Drosophila No Brasil. Papéis Avulsos Zool. 1980, 33, 293–298. [Google Scholar]
- Vilela, C.R. Is Zaprionus indianus Gupta, 1970 (Diptera, Drosophilidae) Currently Colonising the Neotropical Region? Drosoph. Inf. Serv. 1999, 82, 37–38. [Google Scholar]
- Vilela, C.R.; Goñi, B. Is Drosophila nasuta Lamb (Diptera, Drosophilidae) Currently Reaching the Status of a Cosmopolitan Species? Rev. Bras. Entomol. 2015, 59, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Deprá, M.; Poppe, J.L.; Schmitz, H.J.; De Toni, D.C.; Valente, V.L.S. The First Records of the Invasive Pest Drosophila suzukii in the South American Continent. J. Pest Sci. 2014, 87, 379–383. [Google Scholar] [CrossRef]
- Cavalcanti, F.A.G.S.; Ribeiro, L.B.; Marins, G.; Tonelli, G.S.S.S.; Báo, S.N.; Yassin, A.; Tidon, R. Geographic Expansion of an Invasive Fly: First Record of Zaprionus tuberculatus (Diptera: Drosophilidae) in the Americas. Ann. Entomol. Soc. Am. 2022, 115, 267–274. [Google Scholar] [CrossRef]
- Garcia, F.R.M.; Lasa, R.; Funes, C.F.; Buzzetti, K. Drosophila suzukii Management in Latin America: Current Status and Perspectives. J. Econ. Entomol. 2022, 115, 1008–1023. [Google Scholar] [CrossRef]
- Lee, J.C.; Rodriguez-Saona, C.R.; Zalom, F.G. Introductory Remarks: Spotlight on Spotted-Wing Drosophila. J. Econ. Entomol. 2022, 115, 919–921. [Google Scholar] [CrossRef]
- Roque, F.; Matavelli, C.; Lopes, P.H.S.; Machida, W.S.; Von Zuben, C.J.; Tidon, R. Brazilian Fig Plantations Are Dominated by Widely Distributed Drosophilid Species (Diptera: Drosophilidae). Ann. Entomol. Soc. Am. 2017, 110, 521–527. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health (PLH); Bragard, C.; Baptista, P.; Chatzivassiliou, E.; Di Serio, F.; Gonthier, P.; Jaques Miret, J.A.; Justesen, A.F.; Magnusson, C.S.; Milonas, P.; et al. Pest Categorisation of Zaprionus indianus. EFSA J. 2022, 20, e07144. [Google Scholar] [CrossRef]
- Mollot, G.; Pantel, J.H.; Romanuk, T.N. Chapter Two—The Effects of Invasive Species on the Decline in Species Richness: A Global Meta-Analysis. In Advances in Ecological Research; Bohan, D.A., Dumbrell, A.J., Massol, F., Eds.; Networks of Invasion: A Synthesis of Concepts; Academic Press: Cambridge, MA, USA, 2017; Volume 56, pp. 61–83. [Google Scholar]
- da Silva, C.R.B.; Beaman, J.E.; Dorey, J.B.; Barker, S.J.; Congedi, N.C.; Elmer, M.C.; Galvin, S.; Tuiwawa, M.; Stevens, M.I.; Alton, L.A.; et al. Climate Change and Invasive Species: A Physiological Performance Comparison of Invasive and Endemic Bees in Fiji. J. Exp. Biol. 2021, 224, jeb230326. [Google Scholar] [CrossRef]
- Valadão, H.; Proença, C.E.B.; Kuhlmann, M.P.; Harris, S.A.; Tidon, R. Fruit-Breeding Drosophilids (Diptera) in the Neotropics: Playing the Field and Specialising in Generalism? Ecol. Entomol. 2019, 44, 721–737. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, L.B.; Tidon, R. Colonizing Potential of Drosophilidae (Insecta, Diptera) in Environments with Different Grades of Urbanization. Biodivers. Conserv. 2005, 14, 1809–1821. [Google Scholar] [CrossRef]
- Gottschalk, M.S.; De Toni, D.C.; Valente, V.L.S.; Hofmann, P.R.P. Changes in Brazilian Drosophilidae (Diptera) Assemblages across an Urbanisation Gradient. Neotrop. Entomol. 2007, 36, 848–862. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.C.L.; Valiati, V.H.; Gottschalk, M.S.; Rohde, C.; Valente, V.L.d.S. Two Decades of Colonization of the Urban Environment of Porto Alegre, Southern Brazil, by Drosophila paulistorum (Diptera, Drosophilidae). Iheringia Série Zool. 2008, 98, 329–338. [Google Scholar] [CrossRef]
- Bombin, A.; Reed, L.K. The Changing Biodiversity of Alabama Drosophila: Important Impacts of Seasonal Variation, Urbanization, and Invasive Species. Ecol. Evol. 2016, 6, 7057–7069. [Google Scholar] [CrossRef]
- Grimaldi, D.; Ginsberg, P.S.; Thayer, L.; McEvey, S.; Hauser, M.; Turelli, M.; Brown, B. Strange Little Flies in the Big City: Exotic Flower-Breeding Drosophilidae (Diptera) in Urban Los Angeles. PLoS ONE 2015, 10, e0122575. [Google Scholar] [CrossRef] [Green Version]
- Eiten, G. The Cerrado Vegetation of Brazil. Bot. Rev. 1972, 38, 201–341. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity Hotspots for Conservation Priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Oliveira-Filho, A.T.; Ratter, J.A. 6. Vegetation Physiognomies and Woody Flora of the Cerrado Biome. In The Cerrados of Brazil: Ecology and Natural History of a Neotropical Savanna; Oliveira, P.S., Marquis, R.J., Eds.; Columbia University Press: New York, NY, USA, 2002; pp. 91–120. [Google Scholar]
- Ribeiro, L.B.; Cavalcanti, F.A.G.S.; Viana, J.P.C.; Amaral, J.M.; Costa, S.C.; Tidon, R. Updated List of Drosophilid Species Recorded in the Brazilian Savanna. Dros. Inf. Serv. 2022, 105, 85–89. [Google Scholar]
- Tidon, R.; Leite, D.F.; Leão, B.F.D. Impact of the Colonisation of Zaprionus (Diptera, Drosophilidae) in Different Ecosystems of the Neotropical Region: 2 Years after the Invasion. Biol. Conserv. 2003, 112, 299–305. [Google Scholar] [CrossRef]
- Tidon, R. Relationships between Drosophilids (Diptera, Drosophilidae) and the Environment in Two Contrasting Tropical Vegetations. Biol. J. Linn. Soc. 2006, 87, 233–247. [Google Scholar] [CrossRef] [Green Version]
- Roque, F.; Hay, J.D.V.; Tidon, R. Breeding sites of drosophilids (Diptera) in the Brazilian Savanna. I. Fallen fruits of Emmotum nitens (Icacinaceae), Hancornia speciosa (Apocynaceae) and Anacardium humile (Anacardiaceae). Rev. Bras. Entomol. 2009, 53, 308–313. [Google Scholar] [CrossRef]
- Valadão, H.; Hay, J.D.V.; Tidon, R. Temporal Dynamics and Resource Availability for Drosophilid Fruit Flies (Insecta, Diptera) in a Gallery Forest in the Brazilian Savanna. Int. J. Ecol. 2010, 2010, e152437. [Google Scholar] [CrossRef] [Green Version]
- Roque, F.; da Mata, R.A.; Tidon, R. Temporal and Vertical Drosophilid (Insecta; Diptera) Assemblage Fluctuations in a Neotropical Gallery Forest. Biodivers. Conserv. 2013, 22, 657–672. [Google Scholar] [CrossRef]
- Mata, R.A.d.; Valadão, H.; Tidon, R. Spatial and Temporal Dynamics of Drosophilid Larval Assemblages Associated to Fruits. Rev. Bras. Entomol. 2015, 59, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Döge, J.d.S.; de Oliveira, H.V.; Tidon, R. Rapid Response to Abiotic and Biotic Factors Controls Population Growth of Two Invasive Drosophilids (Diptera) in the Brazilian Savanna. Biol. Invasions 2015, 17, 2461–2474. [Google Scholar] [CrossRef]
- Schneider, D.I.; Sujii, E.R.; Laumann, R.A.; Tidon, R. Parasitoids of Drosophilids in the Brazilian Savanna: Spatial–Temporal Distribution and Host Associations with Native and Exotic Species. Neotrop. Entomol. 2022, 51, 514–525. [Google Scholar] [CrossRef]
- Freire-Maia, N.; Pavan, C. Introdução ao Estudo da Drosophila. Rev. Cultus 1949, 1, 3–61. [Google Scholar]
- Yuzuki, K.; Tidon, R. Identification Key for Drosophilid Species (Diptera, Drosophilidae) Exotic to the Neotropical Region and Occurring in Brazil. Rev. Bras. Entomol. 2020, 64, e2019100. [Google Scholar] [CrossRef] [Green Version]
- Val, F.C. Male genitalia of some Neotropical Drosophila: Notes and illustrations. Papéis Avulsos Zool. 1982, 34, 309–347. [Google Scholar]
- Vilela, C.R. A revision of the Drosophila repleta Species Group (Diptera, Drosophilidae). Rev. Bras. Entomol. 1983, 27, 1–114. [Google Scholar]
- The World Flora Online. Available online: http://www.worldfloraonline.org/ (accessed on 19 January 2023).
- Gotelli, N.J.; Colwell, R.K. Quantifying Biodiversity: Procedures and Pitfalls in the Measurement and Comparison of Species Richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package 2022 (Version 2.6-4). Available online: https://CRAN.R-project.org/package=vegan (accessed on 18 January 2023).
- Magurran, A.E. The commonness, and rarity, of species. In Measuring Biological Diversity, 1st ed.; Wiley-Blackwell: Malden, MA, USA, 2003; pp. 18–71. ISBN 978-0-632-05633-0. [Google Scholar]
- Dormann, C.F.; Gruber, B.; Fründ, J. The R Journal: Introducing the Bipartite Package: Analysing Ecological Networks. R News 2008, 8, 8–11. [Google Scholar]
- Magnacca, K.N.; Foote, D.; O’Grady, P.M. A review of the endemic Hawaiian Drosophilidae and their host plants. Zootaxa 2008, 1728, 1–58. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Hoboken, NJ, USA, 1999; p. 663. ISBN 978-0-13-081542-2. [Google Scholar]
- Argemi, M.; Monclús, M.; Mestres, F.; Serra, L. Comparative Analysis of a Community of Drosophilids (Drosophilidae; Diptera) Sampled in Two Periods Widely Separated in Time. J. Zool. Syst. Evol. Res. 1999, 37, 203–210. [Google Scholar] [CrossRef]
- Parsons, P.A. The Evolutionary Biology of Colonizing Species; Cambridge University Press: Cambridge, UK, 1983; p. 262. ISBN 0-521-25247-4. [Google Scholar]
- Kishimoto-Yamada, K.; Itioka, T. How Much Have We Learned about Seasonality in Tropical Insect Abundance since Wolda (1988)? Entomol. Sci. 2015, 18, 407–419. [Google Scholar] [CrossRef]
- Braga, L.; Diniz, I.R. Trophic Interactions of Caterpillars in the Seasonal Environment of the Brazilian Cerrado and Their Importance in the Face of Climate Change. In Caterpillars in the Middle: Tritrophic Interactions in a Changing World; Marquis, R.J., Koptur, S., Eds.; Fascinating Life Sciences; Springer International Publishing: Cham, Switzerland, 2022; pp. 485–508. ISBN 978-3-030-86688-4. [Google Scholar]
- Mavasa, R.; Yekwayo, I.; Mwabvu, T.; Tsvuura, Z. Preliminary Patterns of Seasonal Changes in Species Composition of Surface-Active Arthropods in a South African Savannah. Austral Ecol. 2022, 47, 1222–1231. [Google Scholar] [CrossRef]
- Breitmeyer, C.M.; Markow, T.A. Resource Availability and Population Size in Cactophilic Drosophila. Funct. Ecol. 1998, 12, 14–21. [Google Scholar] [CrossRef]
- Heed, W.B. A New Cactus-Feeding but Soil-Breeding Species of Drosophila (Diptera: Drosophilidae). Proc. Entomol. Soc. Wash. 1977, 79, 649–654. [Google Scholar]
- Markow, T.A.; O’Grady, P. Reproductive Ecology of Drosophila. Funct. Ecol. 2008, 22, 747–759. [Google Scholar] [CrossRef]
- Yang, C.; Belawat, P.; Hafen, E.; Jan, L.Y.; Jan, Y.-N. Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes. Science 2008, 319, 1679–1683. [Google Scholar] [CrossRef] [Green Version]
- Chandler, J.A.; Lang, J.M.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial Communities of Diverse Drosophila Species: Ecological Context of a Host–Microbe Model System. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.A.; Eisen, J.A.; Kopp, A. Yeast Communities of Diverse Drosophila Species: Comparison of Two Symbiont Groups in the Same Hosts. Appl. Environ. Microbiol. 2012, 78, 7327–7336. [Google Scholar] [CrossRef] [Green Version]
- Levins, R.; MacArthur, R. An Hypothesis to Explain the Incidence of Monophagy. Ecology 1969, 50, 910–911. [Google Scholar] [CrossRef]
- Etges, W.J. Evolutionary Genomics of Host Plant Adaptation: Insights from Drosophila. Curr. Opin. Insect Sci. 2019, 36, 96–102. [Google Scholar] [CrossRef]
- Auer, T.O.; Shahandeh, M.P.; Benton, R. Drosophila sechellia: A Genetic Model for Behavioral Evolution and Neuroecology. Annu. Rev. Genet. 2021, 55, 527–554. [Google Scholar] [CrossRef]
- Lauer Garcia, A.C.; Pessoa Da Silva, F.; Campos Bezerra Neves, C.H.; Montes, M.A. Current and Future Potential Global Distribution of the Invading Species Drosophila nasuta (Diptera: Drosophilidae). Biol. J. Linn. Soc. 2022, 135, 208–221. [Google Scholar] [CrossRef]
- Sene, F.M.; Val, F.C.; Vilela, C.R.; Pereira, M.A.Q.R. Preliminary Data on the Geographical Distribution of Drosophila Species within Morphoclimatic Domains of Brazil. Papéis Avulsos Zool. 1980, 33, 315–326. [Google Scholar]
- Bakula, M. The Persistence of a Microbial Flora during Postembryogenesis of Drosophila melanogaster. J. Invertebr. Pathol. 1969, 14, 365–374. [Google Scholar] [CrossRef]
- Gilbert, D.G. Dispersal of Yeasts, and Bacteria by Drosophila in a Temperate Forest. Oecologia 1980, 46, 135–137. [Google Scholar] [CrossRef]
- Atkinson, W.D. A Field Investigation of Larval Competition in Domestic Drosophila. J. Anim. Ecol. 1979, 48, 91–102. [Google Scholar] [CrossRef]
- Sexton, J.P.; Montiel, J.; Shay, J.E.; Stephens, M.R.; Slatyer, R.A. Evolution of Ecological Niche Breadth. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 183–206. [Google Scholar] [CrossRef] [Green Version]
- McKinney, M.L.; Lockwood, J.L. Biotic Homogenization: A Few Winners Replacing Many Losers in the next Mass Extinction. Trends Ecol. Evol. 1999, 14, 450–453. [Google Scholar] [CrossRef]
- Olden, J.D.; Comte, L.; Giam, X. The Homogocene: A Research Prospectus for the Study of Biotic Homogenisation. NeoBiota 2018, 36, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide Decline of the Entomofauna: A Review of Its Drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Wagner, D.L. Insect Declines in the Anthropocene. Annu. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef] [Green Version]
- Didham, R.K.; Basset, Y.; Collins, C.M.; Leather, S.R.; Littlewood, N.A.; Menz, M.H.M.; Müller, J.; Packer, L.; Saunders, M.E.; Schönrogge, K.; et al. Interpreting Insect Declines: Seven Challenges and a Way Forward. Insect Conserv. Divers. 2020, 13, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Mohr, S.E. First in Fly: Drosophila Research and Biologica Discovery; Harvard University Press: Cambridge, MA, USA, 2018; p. 257. ISBN 978-0-674-97101-1. [Google Scholar]
- da Mata, R.A.; McGeoch, M.; Tidon, R. Drosophilids (Insecta, Diptera) as Tools for Conservation Biology. Nat. Conserv. 2010, 8, 60–65. [Google Scholar] [CrossRef]
- Gibert, P.; Hill, M.; Pascual, M.; Plantamp, C.; Terblanche, J.S.; Yassin, A.; Sgrò, C.M. Drosophila as Models to Understand the Adaptive Process during Invasion. Biol. Invasions 2016, 18, 1089–1103. [Google Scholar] [CrossRef]
- Slove, J.; Janz, N. The Relationship between Diet Breadth and Geographic Range Size in the Butterfly Subfamily Nymphalinae—A Study of Global Scale. PLoS ONE 2011, 6, e16057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahner, J.P.; Bonilla, M.M.; Badik, K.J.; Shapiro, A.M.; Forister, M.L. Use of Exotic Hosts by Lepidoptera: Widespread Species Colonize more Novel Hosts. Evolution 2011, 65, 2719–2724. [Google Scholar] [CrossRef] [PubMed]
- Latombe, G.; Pyšek, P.; Jeschke, J.M.; Blackburn, T.M.; Bacher, S.; Capinha, C.; Costello, M.J.; Fernández, M.; Gregory, R.D.; Hobern, D.; et al. A Vision for Global Monitoring of Biological Invasions. Biol. Conserv. 2017, 213, 295–308. [Google Scholar] [CrossRef]
- Simberloff, D.; Von Holle, B. Positive Interactions of Nonindigenous Species. Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- David, J.R.; McEvey, S.F.; Solignac, M.; Tsacas, L. Drosophila Communities on Mauritius and the Ecological Niche of Drosophila mauritiana (Diptera, Drosophilidae). Rev. Zool. Afr. 1989, 103, 107–116. [Google Scholar]
- Montes, M.A.; Neves, C.H.C.B.; Ferreira, A.F.; dos Santos, M.d.F.S.; Quintas, J.I.F.P.; Manetta, G.D.Â.; de Oliveira, P.V.; Garcia, A.C.L. Invasion and Spreading of Drosophila nasuta (Diptera, Drosophilidae) in the Caatinga Biome, Brazil. Neotrop. Entomol. 2021, 50, 571–578. [Google Scholar] [CrossRef]
- Medeiros, H.F.; Monteiro, M.P.; Caçador, A.W.B.; Pereira, C.M.; de Lurdes Bezerra Praxedes, C.; Martins, M.B.; Montes, M.A.; Garcia, A.C.L. First Records of the Invading Species Drosophila nasuta (Diptera: Drosophilidae) in the Amazon. Neotrop. Entomol. 2022, 51, 493–497. [Google Scholar] [CrossRef]
- Paula, M.A.; Lopes, P.H.S.; Tidon, R. First Record of Drosophila suzukii in the Brazilian Savanna. Dros. Inf. Serv. 2014, 97, 113–115. [Google Scholar]
- Wollmann, J.; Schlesener, D.C.H.; Mendes, S.R.; Krüger, A.P.; Martins, L.N.; Bernardi, D.; Garcia, M.S.; Garcia, F.R.M. Infestation Index of Drosophila suzukii (Diptera: Drosophilidae) in Small Fruit in Southern Brazil. Arq. Inst. Biológico 2020, 87, e0432018. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Family | Taxa | Popular Name | Code | Type | Mass (g) | Empty Mass (%) | Collection Period |
|---|---|---|---|---|---|---|---|
| Actinidiaceae | Actinidia chinensis Planch. E | Kiwi; Kiwi | 1 | FF | 592.4 | 86.26 | 1′, 2 |
| Amaranthaceae | Beta vulgaris L. E | Beterraba; Beetroot | 2 | RT | 482.9 | 100 | 2′ |
| Amaryllidaceae | Allium cepa L. E | Cebola; Onion | 3 | SB | 3037.1 | 57.81 | 1, 2 |
| Anacardiaceae | Anacardium occidentale L. N | Caju; Cashew fruit | 4 | FF | 524.0 | 25.38 | 1 |
| Mangifera indica L. E | Manga; Mango | 5 | FF | 8595.8 | 69.42 | 1, 2 | |
| Spondias mombin L. N | Cajá,Cajazinho; Java plum | 6 | FF | 107.2 | 100 | 2′ | |
| Spondias purpurea L. N | Ciriguela, Seriguela; Gambia plum, Purple mombin | 7 | FF | 8.9 | 100 | 2′ | |
| Annonaceae | Annona squamosa L. N | Pinha, Fruta do Conde; Custard apple | 8 | FF | 398.6 | 100 | 2′ |
| Apiaceae | Arracacia xanthorrhiza Bancr. N | Batata baroa, Mandioquinha; Arracache, Peruvian parsnip | 9 | RT | 79.7 | 100 | 2′ |
| Daucus carota L. E | Cenoura; Carrot | 10 | RT | 501.8 | 100 | 2′ | |
| Araceae | Colocasia esculenta (L.) Schott E | Inhame; Cocoyam, Taro | 11 | ST | 711 | 73.42 | 1 |
| Asteraceae | Lactuca sativa L. E | Alface; Lettuce | 12 | VL | 290 | 0 | 1 |
| Brassicaceae | Brassica oleracea L. var. acephala DC. E | Couve; Collard greens, Kale | 13 | VL | 59.9 | 100 | 2′ |
| Brassica oleracea L. var. capitata L. E | Repolho; Cabbage | 14 | VL | 585 | 0 | 1 | |
| Bromeliaceae | Ananas comosus (L.) Merr. N | Abacaxi; Pineapple | 15 | FF | 27,195.5 | 0 | 1, 2 |
| Cactaceae | Selenicereus undatus (Haw.) D.R.Hunt N | Pitaya; Dragon fruit | 16 | FF | 205.9 | 100 | 2′ |
| Caricaceae | Carica papaya L. N | Mamão; Papaya | 17 | FF | 6256.2 | 87.24 | 1′, 2 |
| Caryocaraceae | Caryocar brasiliense Cambess. N | Pequi; no English name | 18 | FF | 171 | 17.54 | 1 |
| Convolvulaceae | Ipomoea batatas (L.) Lam. N | Batata doce; Sweet potato | 19 | RT | 125.2 | 100 | 2′ |
| Cucurbitaceae | Cucumis anguria L. E | Maxixe; West indian gherkin | 20 | FF | 577.7 | 78.38 | 1, 2 |
| Citrullus lanatus (Thunb.) Matsum. and NakaiE | Melancia; Watermelon | 21 | FF | 6071.9 | 2.86 | 1, 2 | |
| Cucumis melo L. E | Melão; Melon | 22 | FF | 962.9 | 0 | 1, 2 | |
| Curcubita moschata Duchesne N | Abóbora; Pumpkin, Winter squash | 23 | FF | 1497.3 | 11.98 | 1, 2 | |
| Cucumis sativus L. E | Pepino; Cucumber | 24 | FF | 844.7 | 58.01 | 1, 2 | |
| Sicyos edulis Jacq. N | Chuchu; Chayote, Corstophine | 25 | FF | 558.9 | 31.10 | 2 | |
| Ebenaceae | Diospyros kaki L.f. E | Caqui; Persimmon | 26 | FF | 282.8 | 100 | 2′ |
| Lauraceae | Persea americana Mill. N | Abacate; Avocado | 27 | FF | 1201.4 | 100 | 2′ |
| Malvaceae | Hibiscus esculentus L. N | Quiabo; Okra, Gumbo, Lady’s fingers | 28 | DF | 62.9 | 100 | 2′ |
| Moraceae | Artocarpus heterophyllus Lam. E | Jaca; Jackfruit | 29 | FF | 1490 | 0 | 1 |
| Musaceae | Musa x paradisiaca L. E | Banana; Banana | 30 | FF | 5860.3 | 34.55 | 1, 2 |
| Myrtaceae | Psidium guajava L. N | Goiaba; Guava | 31 | FF | 1035.7 | 54.28 | 1′, 2 |
| Oxalidaceae | Averrhoa carambola L. E | Carambola; Star fruit | 32 | FF | 105.3 | 13.58 | 1, 2′ |
| Passifloraceae | Passiflora edulis Sims N | Maracujá; Passion fruit | 33 | FF | 1800.9 | 43.17 | 1, 2 |
| Rosaceae | Fragaria vesca L. N | Morango; Strawberry | 34 | FF | 209.8 | 64.59 | 1, 2 |
| Malus domestica (Suckow) Borkh. E | Maçã; Apple | 35 | FF | 5128 | 94.,44 | 1, 2′ | |
| Prunus domestica L. E | Ameixa; Plum | 36 | FF | 1393 | 63.03 | 1, 2 | |
| Prunus persica (L.) Batsch E | Pêssego; Peach | 37 | FF | 709 | 56.56 | 1, 2′ | |
| Prunus persica var. nucipersica (L.)C.K. Schneid. E | Nectarina; Nectarine | 38 | FF | 433.8 | 76.26 | 1, 2′ | |
| Pyrus communis L. E | Pera; Pear | 39 | FF | 1568.4 | 73.64 | 1, 2 | |
| Rutaceae | Citrus x aurantiifolia (Christm.) Swingle E | Limão; Lime | 40 | FF | 571 | 80,14 | 2 |
| Citrus x reticulata Blanco E | Mexirica, Bergamota; Tangerine | 41 | FF | 2391 | 38,62 | 1, 2 | |
| Citrus sinensis (L.) Osbeck E | Laranja; Orange | 42 | FF | 2681.2 | 47.,78 | 1, 2 | |
| Sapindaceae | Litchi chinensis Sonn. E | Lichia; Lychee | 43 | FF | 19.9 | 100 | 2′ |
| Solanaceae | Capsicum annuum L. N | Pimentão; Bell pepper | 44 | FF | 2346.5 | 71.02 | 1, 2 |
| Capsicum chinense L. N | Pimenta; Chili pepper | 45 | FF | 25.5 | 100 | 2′ | |
| Solanum aethiopicum L. E | Jiló; Bitterberry | 46 | FF | 239.6 | 100 | 2′ | |
| Solanum lycopersicum Lam. N | Tomate; Tomato | 47 | FF | 5592 | 39.33 | 1, 2 | |
| Solanum melongena L. E | Beringela; Eggplant, Aubergine | 48 | FF | 360.5 | 39.92 | 2 | |
| Solanum tuberosum L. N | Batata; Potato | 49 | ST | 3420.4 | 79.69 | 1, 2 | |
| Vitaceae | Vitis vinifera L. × Vitis labrusca EN | Uva; Grape | 50 | FF | 108.4 | 40.96 | 1, 2′ |
| Genus | Subgenus | Group | Species | Code | Plant Fam/Spp | Abundance | |
|---|---|---|---|---|---|---|---|
| 2007–2008 | 2018–2019 | ||||||
| Drosophila | Dorsilopha | busckii | D. busckii Coquillett E | A | 6/13 | 1198 | 1647 |
| Drosophila | cardini | D. cardini Sturtevant N | B | 9/14 | 121 | 412 | |
| D. cardinoides Dobzhansky and Pavan N | C | 1/2 | 0 | 11 | |||
| immigrans | D. immigrans Sturtevant E | D | 4/4 | 8 | 140 | ||
| D. nasuta Lamb E | E | 1/1 | 0 | 211 | |||
| repleta | D. hydei Sturtevant E | F | 11/14 | 14,361 | 1049 | ||
| D. mercatorum Patterson and Wheeler N | G | 9/13 | 299 | 472 | |||
| D. repleta Wollaston N | H | 1/1 | 0 | 17 | |||
| willistoni | D. nebulosa Sturtevant N | I | 4/4 | 25 | 0 | ||
| Sophophora | melanogaster | D. ananassae Doleschall E | J | 10/14 | 424 | 2775 | |
| D. kikkawai Burla E | K | 1/1 | 2 | 0 | |||
| D. malerkotliana Parshad and Paika E | L | 6/6 | 592 | 0 | |||
| D. melanogaster Meigen E | M | 11/15 | 6838 | 3225 | |||
| D. simulans Sturtevant E | N | 15/22 | 7824 | 3739 | |||
| saltans | D. sturtevanti Duda N | O | 1/1 | 0 | 85 | ||
| Zaprionus | Z. indianus Gupta E | P | 14/20 | 2154 | 1265 | ||
| Plant Hosts | ||||
|---|---|---|---|---|
| Drosophilids | Neotropical (21 Taxa) | Exotic (29 Taxa) | ||
| Neotropical (6 species) | 19/126 = 15.08% | (p = 0.067) | 15/174 = 8.93% | (p < 0.001) |
| Exotic (10 species) | 40/210 = 19.05% | (p = 0.064) | 68/280 = 24.28% | (p = 0.012) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, L.B.; Proença, C.E.B.; Tidon, R. Host Preferences Shown by Drosophilids (Diptera) in a Commercial Fruit and Vegetable Distribution Center Follow the Wild Neotropical Pattern. Insects 2023, 14, 375. https://doi.org/10.3390/insects14040375
Ribeiro LB, Proença CEB, Tidon R. Host Preferences Shown by Drosophilids (Diptera) in a Commercial Fruit and Vegetable Distribution Center Follow the Wild Neotropical Pattern. Insects. 2023; 14(4):375. https://doi.org/10.3390/insects14040375
Chicago/Turabian StyleRibeiro, Laís Barbosa, Carolyn Elinore Barnes Proença, and Rosana Tidon. 2023. "Host Preferences Shown by Drosophilids (Diptera) in a Commercial Fruit and Vegetable Distribution Center Follow the Wild Neotropical Pattern" Insects 14, no. 4: 375. https://doi.org/10.3390/insects14040375
APA StyleRibeiro, L. B., Proença, C. E. B., & Tidon, R. (2023). Host Preferences Shown by Drosophilids (Diptera) in a Commercial Fruit and Vegetable Distribution Center Follow the Wild Neotropical Pattern. Insects, 14(4), 375. https://doi.org/10.3390/insects14040375