
Seasonal Dynamics of Tick Species in the Ecotone of Parks and Recreational Areas in Middlesex County (New Jersey, USA)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
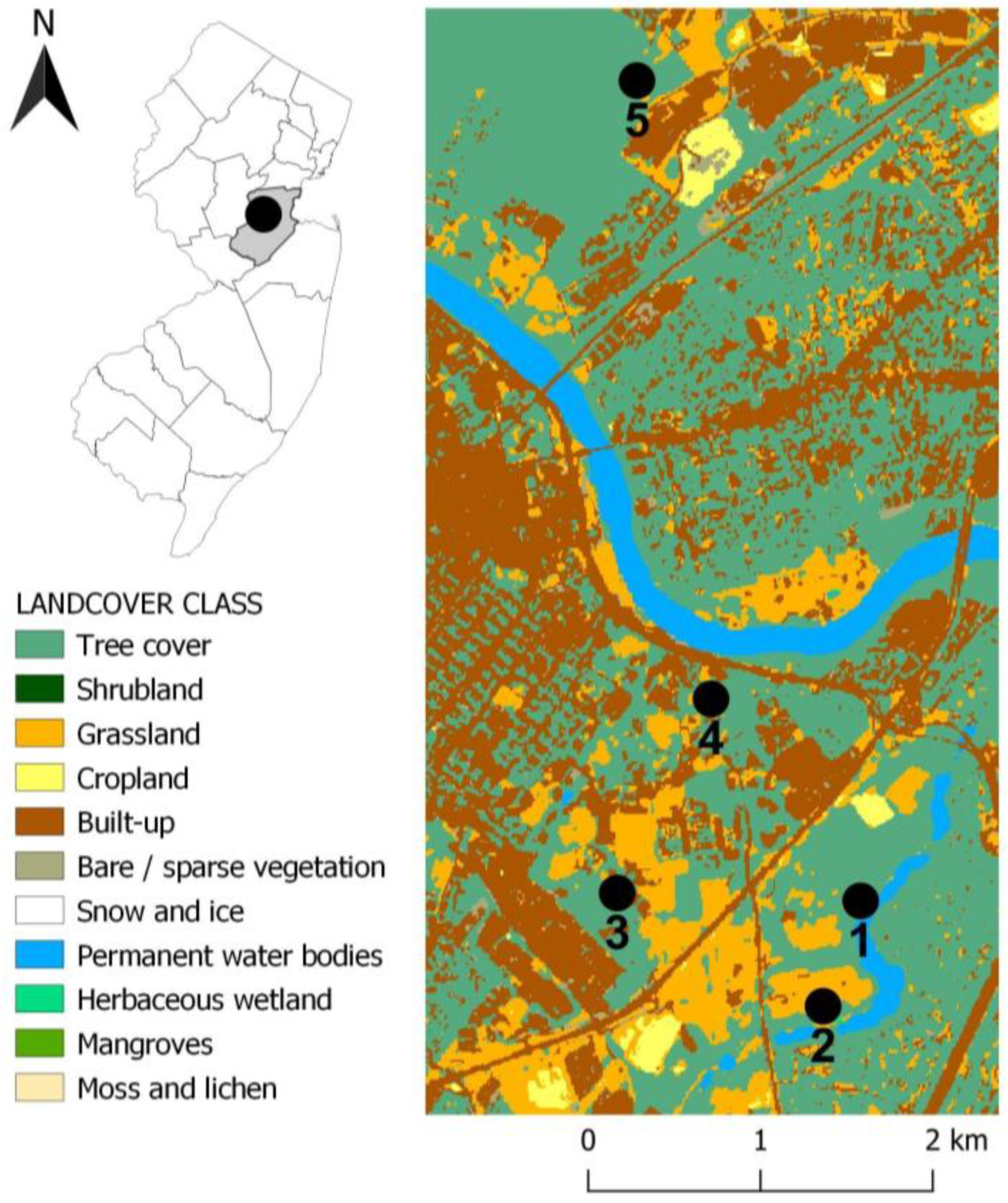
2.1. Study Area
2.2. Tick Surveillance
2.3. Data Analyses
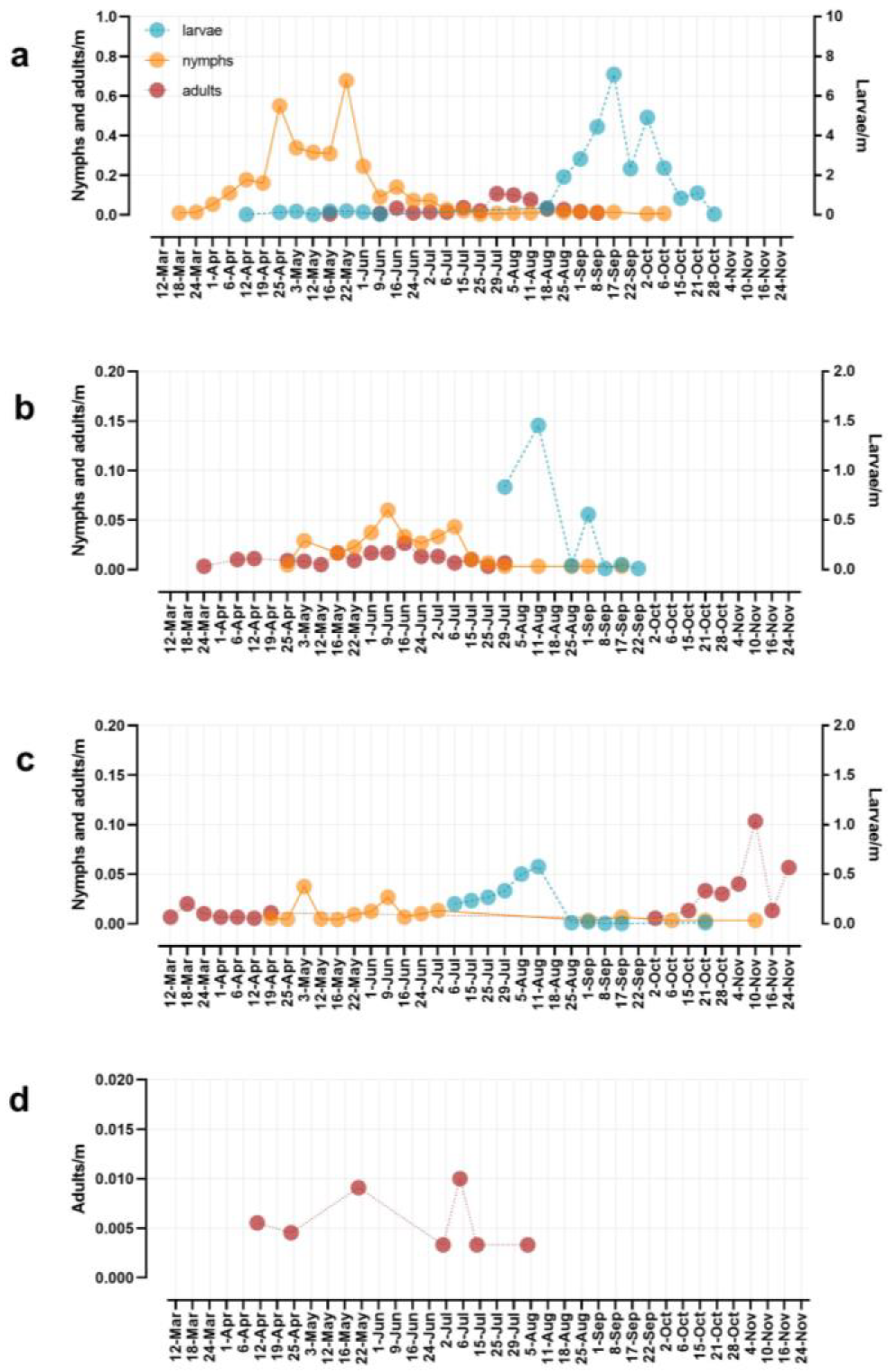
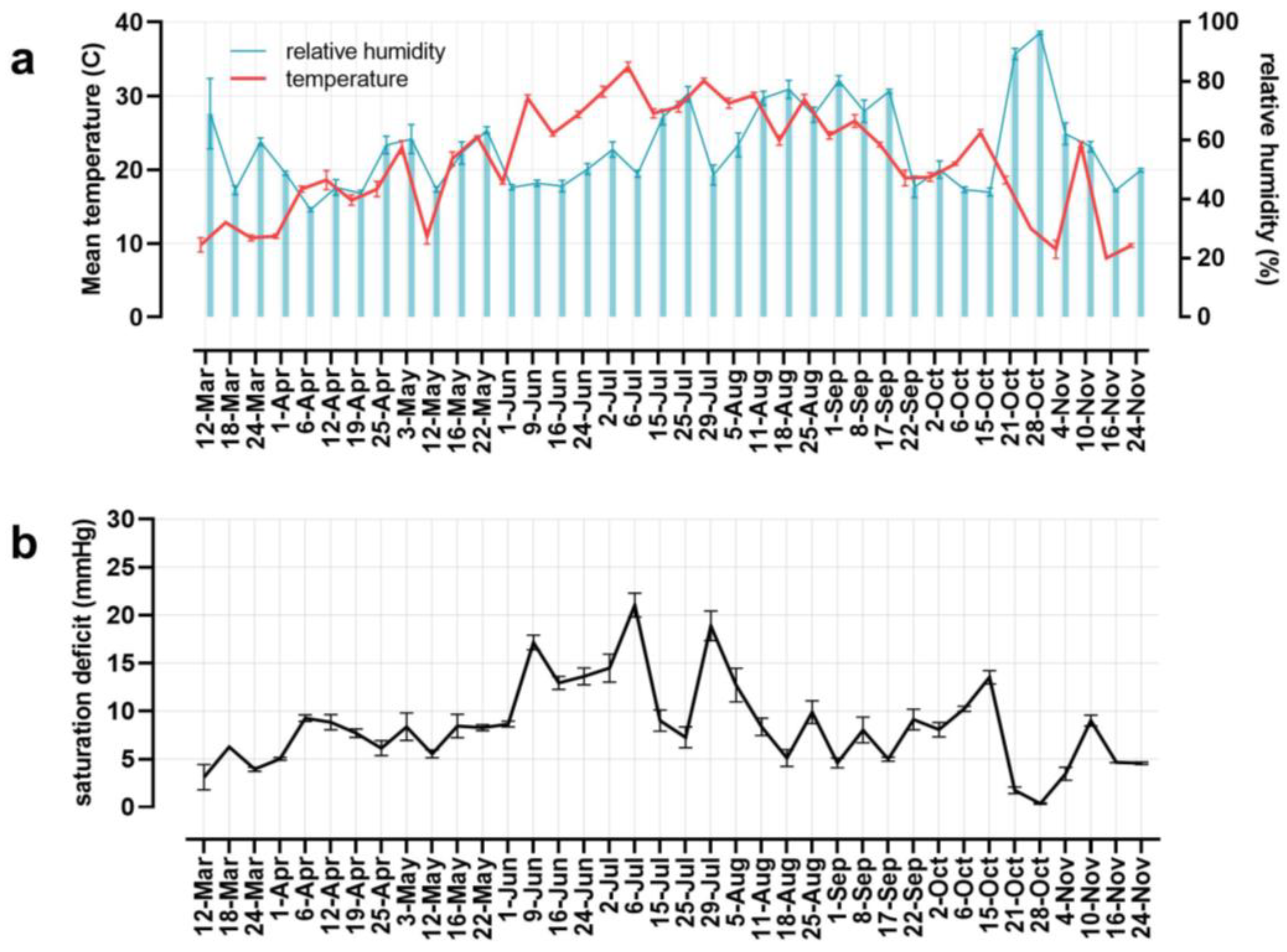
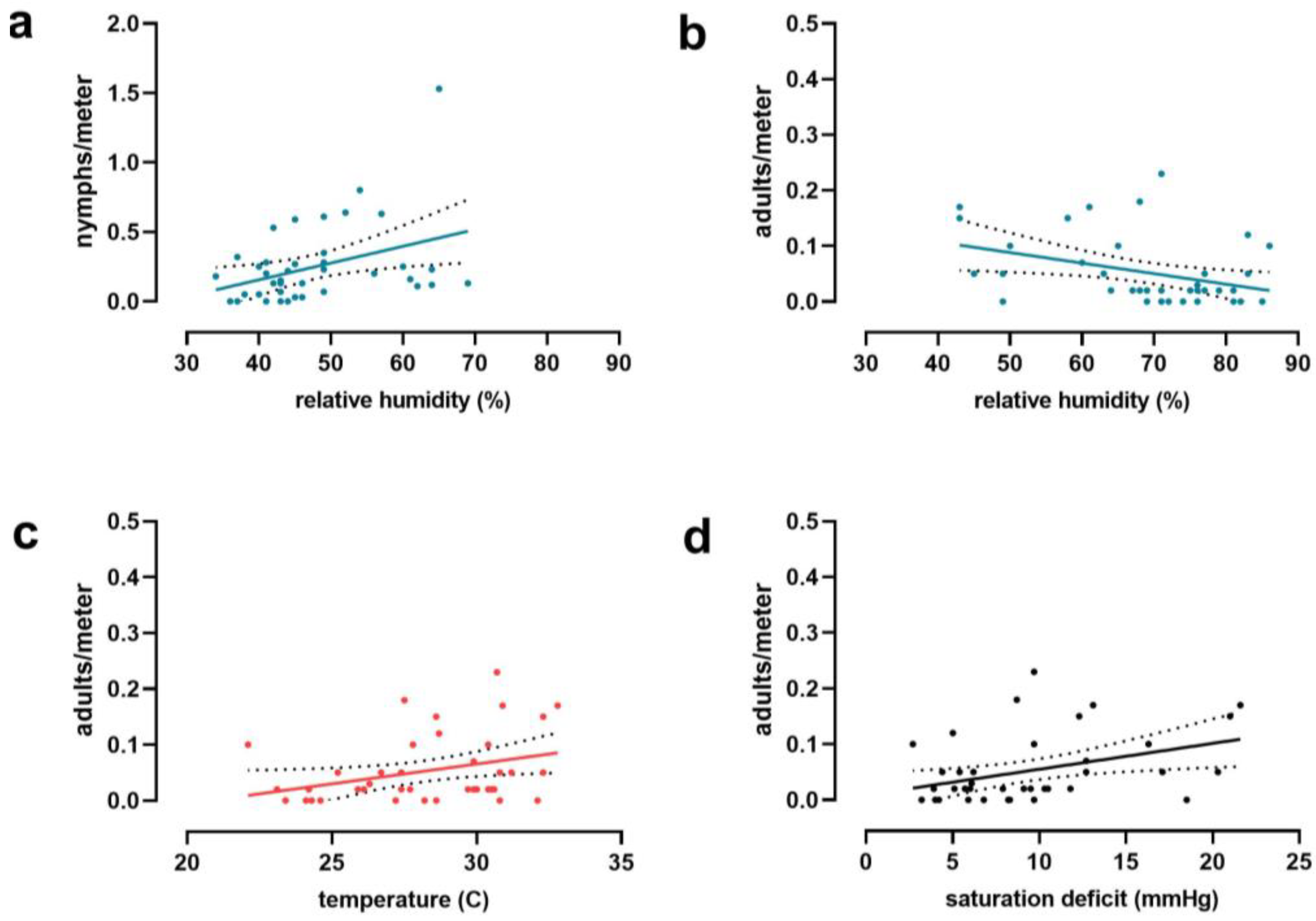
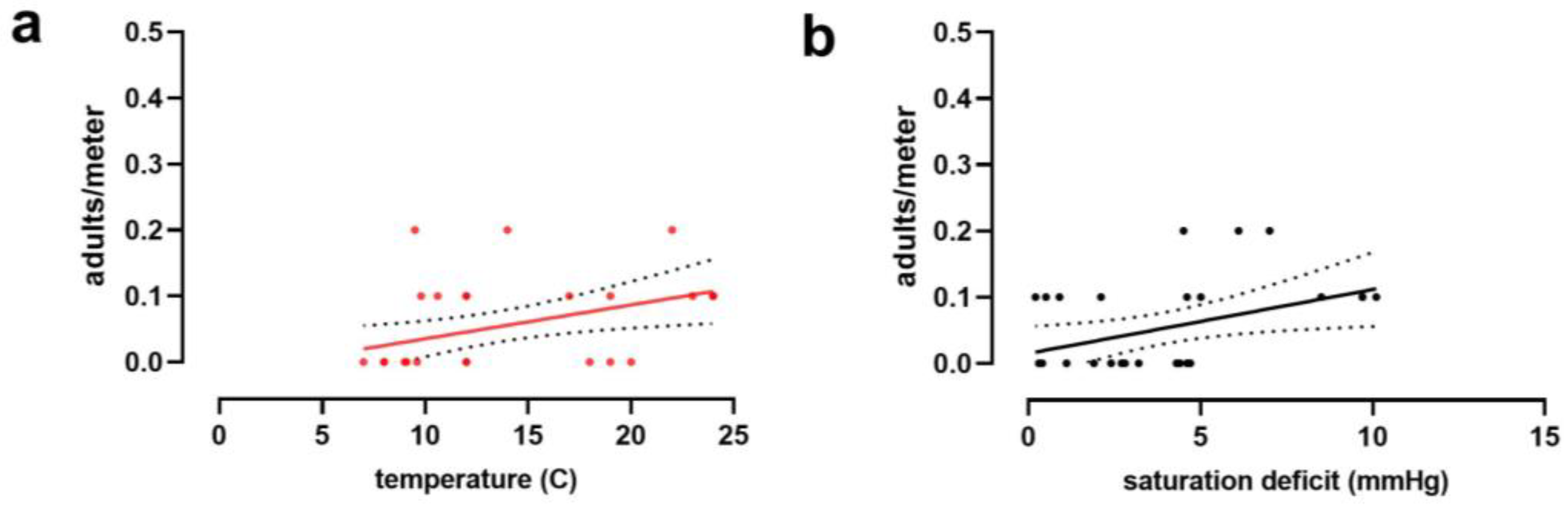
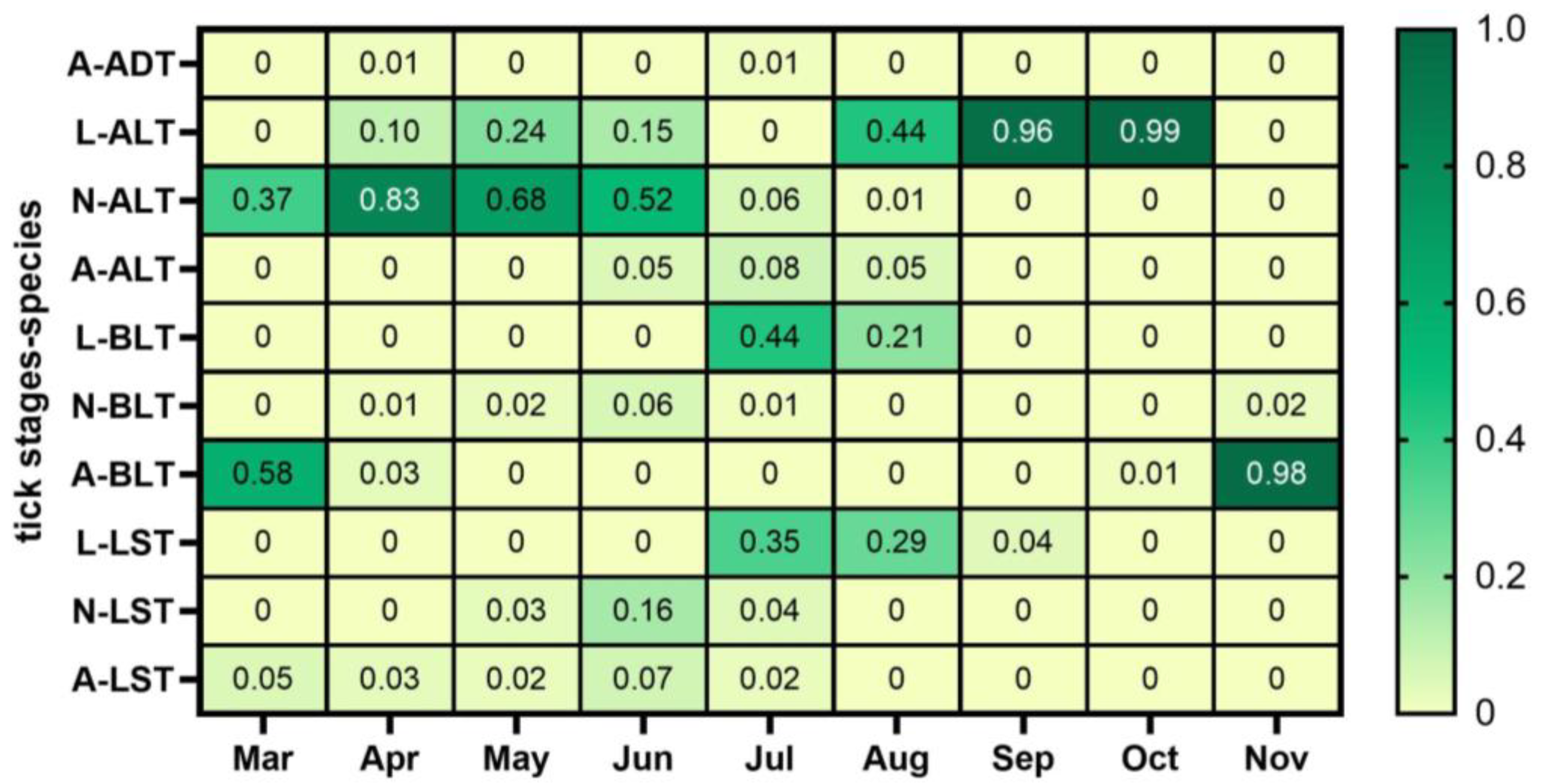
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beard, C.B.; Eisen, L.; Eisen, R.J. The Rise of Ticks and Tickborne Diseases in the United States—Introduction. J. Med. Entomol. 2021, 58, 1487–1489. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, R.; Lindsey, N.P.; Fischer, M.; Gregory, C.J.; Hinckley, A.F.; Mead, P.S.; Paz-Bailey, G.; Waterman, S.H.; Drexler, N.A.; Kersh, G.J.; et al. Vital Signs: Trends in Reported Vectorborne Disease Cases—United States and Territories, 2004–2016. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 496–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochlin, I.; Egizi, A.; Lindström, A. The Original Scientific Description of the Lone Star Tick (Amblyomma americanum, Acari: Ixodidae) and Implications for the Species’ Past and Future Geographic Distributions. J. Med. Entomol. 2022, 59, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Rochlin, I.; Toledo, A. Emerging Tick-Borne Pathogens of Public Health Importance: A Mini-Review. J. Med. Microbiol. 2020, 69, 781–791. [Google Scholar] [CrossRef]
- Eisen, R.J.; Paddock, C.D. Tick and Tickborne Pathogen Surveillance as a Public Health Tool in the United States. J. Med. Entomol. 2021, 58, 1490–1502. [Google Scholar] [CrossRef]
- Ogden, N.H.; Ben Beard, C.; Ginsberg, H.S.; Tsao, J.I. Possible Effects of Climate Change on Ixodid Ticks and the Pathogens They Transmit: Predictions and Observations. J. Med. Entomol. 2021, 58, 1536–1545. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, H.S.; Ewing, C.P. Comparison of Flagging, Walking, Trapping, and Collecting from Hosts as Sampling Methods for Northern Deer Ticks, Ixodes dammini, and Lone- Star Ticks, Amblyomma americanum (Acari: Ixodidae). Exp. Appl. Acarol. 1989, 7, 313–322. [Google Scholar] [CrossRef]
- Lord, C.C. Seasonal Activity of Nymphal Ixodes scapularis (Acari: Ixodidae) in Different Habitats in New Jersey. J. Med. Entomol. 1995, 32, 66–70. [Google Scholar] [CrossRef]
- Schulze, T.L.; Jordan, R.A. Influence of Meso- and Microscale Habitat Structure on Focal Distribution of Sympatric Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 2005, 42, 285–294. [Google Scholar] [CrossRef]
- Schulze, T.L.; Jordan, R.A.; White, J.C.; Roegner, V.E.; Healy, S.P. Geographical Distribution and Prevalence of Selected Borrelia, Ehrlichia, and Rickettsia Infections in Amblyomma americanum (Acari: Ixodidae) in New Jersey. J. Am. Mosq. Control Assoc. 2011, 27, 236–244. [Google Scholar] [CrossRef]
- Stafford, K.C. Reduced Abundance of Ixodes scapularis (Acari: Ixodidae) with Exclusion of Deer by Electric Fencing. J. Med. Entomol. 1993, 30, 986–996. [Google Scholar] [CrossRef] [PubMed]
- Dronova, I. Environmental Heterogeneity as a Bridge between Ecosystem Service and Visual Quality Objectives in Management, Planning and Design. Landsc. Urban Plan. 2017, 163, 90–106. [Google Scholar] [CrossRef]
- Dröse, W.; Podgaiski, L.R.; Dias, C.F.; Mendonça, M.d.S., Jr. Local and Regional Drivers of Ant Communities in Forest-Grassland Ecotones in South Brazil: A Taxonomic and Phylogenetic Approach. PLoS ONE 2019, 14, e0215310. [Google Scholar] [CrossRef]
- Bond, W.J.; Parr, C.L. Beyond the Forest Edge: Ecology, Diversity and Conservation of the Grassy Biomes. Biol. Conserv. 2010, 143, 2395–2404. [Google Scholar] [CrossRef]
- Despommier, D.; Ellis, B.R.; Wilcox, B.A. The Role of Ecotones in Emerging Infectious Diseases. EcoHealth 2006, 3, 281–289. [Google Scholar] [CrossRef]
- Eisen, L.; Stafford, K.C. Barriers to Effective Tick Management and Tick-Bite Prevention in the United States (Acari: Ixodidae). J. Med. Entomol. 2021, 58, 1588–1600. [Google Scholar] [CrossRef]
- McKay, R.; Talbot, B.; Slatculescu, A.; Stone, A.; Kulkarni, M.A. Woodchip Borders at the Forest Ecotone as an Environmental Control Measure to Reduce Questing Tick Density along Recreational Trails in Ottawa, Canada. Ticks Tick-Borne Dis. 2020, 11, 101361. [Google Scholar] [CrossRef] [PubMed]
- CDC Tickborne Disease Surveillance Data Summary|CDC. Available online: https://www.cdc.gov/ticks/data-summary/index.html (accessed on 13 November 2021).
- Zanaga, D.; Van De Kerchove, R.; De Keersmaecker, W.; Souverijns, N.; Brockmann, C.; Quast, R.; Wevers, J.; Grosu, A.; Paccini, A.; Vergnaud, S.; et al. WorldCover 10 m 2020 v100; European Space Agency (ESA): Paris, France, 2021. [Google Scholar] [CrossRef]
- Breden, T.F.; Alger, Y.; Walz, K.S.; Windisch, A.G. Classification of Vegetation Communities of New Jersey: Second Iteration; Association for Biodiversity Information and New Jersey Natural Heritage Program, Office of Natural Lands Management, Division of Parks and Forestry, NJ Department of Environmental Protection: Trenton, NJ, USA, 2001; p. 230. [Google Scholar]
- ONJSC Monthly Climate Tables. Office of the New Jersey State Climatologist. Rutgers University. Available online: https://climate.rutgers.edu/stateclim_v1/nclimdiv/index.php?stn=NJ023&elem=pcpn (accessed on 26 February 2023).
- Egizi, A.M.; Occi, J.L.; Price, D.C.; Fonseca, D.M. Leveraging the Expertise of the New Jersey Mosquito Control Community to Jump Start Standardized Tick Surveillance. Insects 2019, 10, 219. [Google Scholar] [CrossRef] [Green Version]
- Bickerton, M.; McSorley, K.; Toledo, A. A Life Stage-Targeted Acaricide Application Approach for the Control of Haemaphysalis longicornis. Ticks Tick-Borne Dis. 2021, 12, 101581. [Google Scholar] [CrossRef] [PubMed]
- Durden, L.A.; Keirans, J.E. Nymphs of the Genus Ixodes (Acari: Ixodidae) of the United States: Taxonomy, Identification Key, Distribution, Hosts, and Medical/Veterinary Importance; Entomological Society of America: Annapolis, MD, USA, 1996; ISBN 978-0-938522-57-7. [Google Scholar]
- Egizi, A.M.; Robbins, R.G.; Beati, L.; Nava, S.; Evans, C.R.; Occi, J.L.; Fonseca, D.M. A Pictorial Key to Differentiate the Recently Detected Exotic Haemaphysalis longicornis Neumann, 1901 (Acari, Ixodidae) from Native Congeners in North America. ZooKeys 2019, 818, 117–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keirans, J.E.; Durden, L.A. Illustrated Key to Nymphs of the Tick Genus Amblyomma (Acari: Ixodidae) Found in the United States. J. Med. Entomol. 1998, 35, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E.; Storey, K. Impact of Microclimate on Immature Tick-Rodent Host Interactions (Acari: Ixodidae): Implications for Parasite Transmission. J. Med. Entomol. 1999, 36, 741–748. [Google Scholar] [CrossRef]
- GraphPad Software GraphPad Prism for Windows 2019.
- Rainey, T.; Occi, J.L.; Robbins, R.G.; Egizi, A. Discovery of Haemaphysalis longicornis (Ixodida: Ixodidae) Parasitizing a Sheep in New Jersey, United States. J. Med. Entomol. 2018, 55, 757–759. [Google Scholar] [CrossRef] [PubMed]
- Bickerton, M.; Rochlin, I.; González, J.; McSorley, K.; Toledo, A. Field Applications of Granular and Liquid Pyrethroids, Carbaryl, and IGRs to Control the Asian Longhorned Tick (Haemaphysalis longicornis) and Impacts on Nontarget Invertebrates. Ticks Tick-Borne Dis. 2022, 13, 102054. [Google Scholar] [CrossRef]
- Cumbie, A.N.; Whitlow, A.M.; Arneson, A.; Du, Z.; Eastwood, G. The Distribution, Seasonal Abundance, and Environmental Factors Contributing to the Presence of the Asian Longhorned Tick (Haemaphysalis longicornis, Acari: Ixodidae) in Central Appalachian Virginia. J. Med. Entomol. 2022, 59, tjac067. [Google Scholar] [CrossRef] [PubMed]
- Piedmonte, N.P.; Vinci, V.C.; Daniels, T.J.; Backenson, B.P.; Falco, R.C. Seasonal Activity of Haemaphysalis longicornis (Acari: Ixodidae) in Southern New York State. J. Med. Entomol. 2021, 58, 676–681. [Google Scholar] [CrossRef]
- Price, K.J.; Witmier, B.J.; Eckert, R.A.; Boyer, C.N.; Helwig, M.W.; Kyle, A.D. Distribution and Density of Haemaphysalis longicornis (Acari: Ixodidae) on Public Lands in Pennsylvania, United States. J. Med. Entomol. 2021, 58, 1433–1438. [Google Scholar] [CrossRef]
- Tufts, D.M.; VanAcker, M.C.; Fernandez, M.P.; DeNicola, A.; Egizi, A.; Diuk-Wasser, M.A. Distribution, Host-Seeking Phenology, and Host and Habitat Associations of Haemaphysalis longicornis Ticks, Staten Island, New York, USA. Emerg. Infect. Dis. 2019, 25, 792–796. [Google Scholar] [CrossRef] [Green Version]
- Cadenasso, M.L.; Traynor, M.M.; Pickett, S.T. Functional Location of Forest Edges: Gradients of Multiple Physical Factors. Can. J. For. Res. 1997, 27, 774–782. [Google Scholar] [CrossRef]
- Heath, A. Biology, Ecology and Distribution of the Tick, Haemaphysalis longicornis Neumann (Acari: Ixodidae) in New Zealand. N. Z. Vet. J. 2016, 64, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, J.; Cui, X.; Jia, N.; Wei, J.; Xia, L.; Wang, H.; Zhou, Y.; Wang, Q.; Liu, X.; et al. Distribution of Haemaphysalis longicornis and Associated Pathogens: Analysis of Pooled Data from a China Field Survey and Global Published Data. Lancet Planet. Health 2020, 4, e320–e329. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Yu, Z.; Zhou, L.; Yang, X.; Liu, J. Seasonal Abundance and Activity of the Hard Tick Haemaphysalis longicornis (Acari: Ixodidae) in North China. Exp. Appl. Acarol. 2012, 56, 133–141. [Google Scholar] [CrossRef]
- Trout Fryxell, R.T.; Moore, J.E.; Collins, M.D.; Kwon, Y.; Jean-Philippe, S.R.; Schaeffer, S.M.; Odoi, A.; Kennedy, M.; Houston, A.E. Habitat and Vegetation Variables Are Not Enough When Predicting Tick Populations in the Southeastern United States. PLoS ONE 2015, 10, e0144092. [Google Scholar] [CrossRef] [PubMed]
- Tufts, D.M.; Goodman, L.B.; Benedict, M.C.; Davis, A.D.; VanAcker, M.C.; Diuk-Wasser, M. Association of the Invasive Haemaphysalis longicornis Tick with Vertebrate Hosts, Other Native Tick Vectors, and Tick-Borne Pathogens in New York City, USA. Int. J. Parasitol. 2021, 51, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Ronai, I.; Tufts, D.M.; Diuk-Wasser, M.A. Aversion of the Invasive Asian Longhorned Tick to the White-Footed Mouse, the Dominant Reservoir of Tick-Borne Pathogens in the U.S.A. Med. Vet. Entomol. 2020, 34, 369–373. [Google Scholar] [CrossRef]
- Chong, S.T.; Kim, H.C.; Lee, I.-Y.; Kollars, T.M.; Sancho, A.R.; Sames, W.J.; Klein, T.A. Comparison of Dragging and Sweeping Methods for Collecting Ticks and Determining Their Seasonal Distributions for Various Habitats, Gyeonggi Province, Republic of Korea. J. Med. Entomol. 2013, 50, 611–618. [Google Scholar] [CrossRef]
- Tsunoda, T. Interspecific and Intraspecific Associations of Two Species of Hard Ticks, Haemaphysalis longicornis and Haemaphysalis megaspinosa, in Relation to Questing Site. J. Parasitol. 2007, 93, 531–541. [Google Scholar] [CrossRef]
- Whitlow, A.M.; Schürch, R.; Mullins, D.; Eastwood, G. The Influence of Southwestern Virginia Environmental Conditions on the Potential Ability of Haemaphysalis longicornis, Amblyomma americanum, and Amblyomma maculatum to Overwinter in the Region. Insects 2021, 12, 1000. [Google Scholar] [CrossRef]
- Bickerton, M.; Toledo, A. Multiple Pruritic Tick Bites by Asian Longhorned Tick Larvae (Haemaphysalis longicornis). Int. J. Acarol. 2020, 46, 373–376. [Google Scholar] [CrossRef]
- Wormser, G.P.; McKenna, D.; Piedmonte, N.; Vinci, V.; Egizi, A.M.; Backenson, B.; Falco, R.C. First Recognized Human Bite in the United States by the Asian Longhorned Tick, Haemaphysalis longicornis. Clin. Infect. Dis. 2020, 70, 314–316. [Google Scholar] [CrossRef]
- Breuner, N.E.; Ford, S.L.; Hojgaard, A.; Osikowicz, L.M.; Parise, C.M.; Rosales Rizzo, M.F.; Bai, Y.; Levin, M.L.; Eisen, R.J.; Eisen, L. Failure of the Asian Longhorned Tick, Haemaphysalis longicornis, to Serve as an Experimental Vector of the Lyme Disease Spirochete, Borrelia burgdorferi Sensu Stricto. Ticks Tick-Borne Dis. 2020, 11, 101311. [Google Scholar] [CrossRef]
- Levin, M.L.; Stanley, H.M.; Hartzer, K.; Snellgrove, A.N. Incompetence of the Asian Longhorned Tick (Acari: Ixodidae) in Transmitting the Agent of Human Granulocytic Anaplasmosis in the United States. J. Med. Entomol. 2021, 58, 1419–1423. [Google Scholar] [CrossRef]
- Tully, B.G.; Huntley, J.F. A Francisella Tularensis Chitinase Contributes to Bacterial Persistence and Replication in Two Major U.S. Tick Vectors. Pathogens 2020, 9, 1037. [Google Scholar] [CrossRef]
- Stanley, H.M.; Ford, S.L.; Snellgrove, A.N.; Hartzer, K.; Smith, E.B.; Krapiunaya, I.; Levin, M.L. The Ability of the Invasive Asian Longhorned Tick Haemaphysalis longicornis (Acari: Ixodidae) to Acquire and Transmit Rickettsia rickettsii (Rickettsiales: Rickettsiaceae), the Agent of Rocky Mountain Spotted Fever, Under Laboratory Conditions. J. Med. Entomol. 2020, 57, 1635–1639. [Google Scholar] [CrossRef]
- Raney, W.R.; Perry, J.B.; Hermance, M.E. Transovarial Transmission of Heartland Virus by Invasive Asian Longhorned Ticks under Laboratory Conditions. Emerg. Infect. Dis. 2022, 28, 726–729. [Google Scholar] [CrossRef] [PubMed]
- Cumbie, A.N.; Trimble, R.N.; Eastwood, G. Pathogen Spillover to an Invasive Tick Species: First Detection of Bourbon Virus in Haemaphysalis longicornis in the United States. Pathogens 2022, 11, 454. [Google Scholar] [CrossRef]
- Chomel, B. Tick-Borne Infections in Dogs—An Emerging Infectious Threat. Vet. Parasitol. 2011, 179, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Greay, T.L.; Oskam, C.L.; Gofton, A.W.; Rees, R.L.; Ryan, U.M.; Irwin, P.J. A Survey of Ticks (Acari: Ixodidae) of Companion Animals in Australia. Parasites Vectors 2016, 9, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boozer, L.; Macintire, D. Babesia Gibsoni: An Emerging Pathogen in Dogs. In The Compendium on Continuing Education for the Practicing Veterinarian; Veterinary Learning Systems: Yardley, PA, USA, 2005. [Google Scholar]
- Higuchi, S.; Simomura, S.; Yoshida, H.; Hoshi, F.; Kawamura, S.; Yasuda, Y. Development of Babesia gibsoni in the Hemolymph of the Vector Tick, Haemaphysalis longicornis. J. Vet. Med. Sci. 1991, 53, 491–493. [Google Scholar] [CrossRef] [Green Version]
- Evers, H.V.; Kocan, A.A.; Reichard, M.V.; Meinkoth, J.H. Experimental Babesia gibsoni Infection in Coyotes (Canis Latrans). J. Wildl. Dis. 2003, 39, 904–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, J.; Playford, M.; Hickey, K. Theileria orientalis: A Review. N. Z. Vet. J. 2016, 64, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.T.; White, S.; Shaw, D.; Egizi, A.; Lahmers, K.; Ruder, M.G.; Yabsley, M.J. Theileria orientalis Ikeda in Host-Seeking Haemaphysalis longicornis in Virginia, U.S.A. Ticks Tick-Borne Dis. 2020, 11, 101450. [Google Scholar] [CrossRef]
- Oakes, V.J.; Yabsley, M.J.; Schwartz, D.; LeRoith, T.; Bissett, C.; Broaddus, C.; Schlater, J.L.; Todd, S.M.; Boes, K.M.; Brookhart, M.; et al. Theileria Orientalis Ikeda Genotype in Cattle, Virginia, USA. Emerg. Infect. Dis. 2019, 25, 1653–1659. [Google Scholar] [CrossRef] [Green Version]
- Dinkel, K.D.; Herndon, D.R.; Noh, S.M.; Lahmers, K.K.; Todd, S.M.; Ueti, M.W.; Scoles, G.A.; Mason, K.L.; Fry, L.M. A U.S. Isolate of Theileria orientalis, Ikeda Genotype, Is Transmitted to Cattle by the Invasive Asian Longhorned Tick, Haemaphysalis longicornis. Parasites Vectors 2021, 14, 157. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Zhioua, E. Nymphal Survival and Habitat Distribution of Ixodes scapularis and Amblyomma americanum Ticks (Acari: Ixodidae) on Fire Island, New York, USA. Exp. Appl. Acarol. 1996, 20, 533–544. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Zhioua, E.; Mitra, S.; Fischer, J.; Buckley, P.A.; Verret, F.; Underwood, H.B.; Buckley, F.G. Woodland Type and Spatial Distribution of Nymphal Ixodes scapularis (Acari: Ixodidae). Environ. Entomol. 2004, 33, 1266–1273. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.B.; Bjork, J.K.H.; Neitzel, D.F.; Dorr, F.M.; Whitemarsh, T.; Boegler, K.A.; Graham, C.B.; Johnson, T.L.; Maes, S.E.; Eisen, R.J. Evaluating Acarological Risk for Exposure to Ixodes scapularis and Ixodes scapularis-Borne Pathogens in Recreational and Residential Settings in Washington County, Minnesota. Ticks Tick-Borne Dis. 2018, 9, 340–348. [Google Scholar] [CrossRef]
- Stafford, K.C.; Magnarelli, L.A. Spatial and Temporal Patterns of Ixodes scapularis (Acari: Ixodidae) in Southeastern Connecticut. J. Med. Entomol. 1993, 30, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, H.S.; Ewing, C.P. Habitat Distribution of Ixodes dammini (Acari: Ixodidae) an Lyme Disease Spirochetes on Fire Island, New York. J. Med. Entomol. 1989, 26, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Linske, M.A.; Stafford, K.C.; Williams, S.C.; Lubelczyk, C.B.; Welch, M.; Henderson, E.F. Impacts of Deciduous Leaf Litter and Snow Presence on Nymphal Ixodes scapularis (Acari: Ixodidae) Overwintering Survival in Coastal New England, USA. Insects 2019, 10, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loydi, A.; Lohse, K.; Otte, A.; Donath, T.W.; Eckstein, R.L. Distribution and Effects of Tree Leaf Litter on Vegetation Composition and Biomass in a Forest–Grassland Ecotone. J. Plant Ecol. 2014, 7, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.C.; BCE; Marshall, E. Lone Star Ticks (Amblyomma americanum): An Emerging Threat in Delaware. Del. J. Public Health 2021, 7, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Mixson, T.R.; Campbell, S.R.; Gill, J.S.; Ginsberg, H.S.; Reichard, M.V.; Schulze, T.L.; Dasch, G.A. Prevalence of Ehrlichia, Borrelia, and Rickettsial Agents in Amblyomma americanum (Acari: Ixodidae) Collected from Nine States. J. Med. Entomol. 2006, 43, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Jordan, R.A.; Egizi, A. The Growing Importance of Lone Star Ticks in a Lyme Disease Endemic County: Passive Tick Surveillance in Monmouth County, NJ, 2006–2016. PLoS ONE 2019, 14, e0211778. [Google Scholar] [CrossRef] [Green Version]
- Schulze, T.L.; Bowen, G.S.; Lakat, M.F.; Parkin, W.E.; Shisler, J.K. Seasonal Abundance and Hosts of Ixodes dammini (Acari: Ixodidae) and Other Ixodid Ticks from an Endemic Lyme Disease Focus in New Jersey, USA. J. Med. Entomol. 1986, 23, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Goddard, J.; Varela-Stokes, A.S. Role of the Lone Star Tick, Amblyomma americanum (L.), in Human and Animal Diseases. Vet. Parasitol. 2009, 160, 1–12. [Google Scholar] [CrossRef]
- Zellner, B.; Huntley, J.F. Ticks and Tularemia: Do We Know What We Don’t Know? Front. Cell. Infect. Microbiol. 2019, 9, 146. [Google Scholar] [CrossRef]
- Commins, S.P.; James, H.R.; Kelly, E.A.; Pochan, S.L.; Workman, L.J.; Perzanowski, M.S.; Kocan, K.M.; Fahy, J.V.; Nganga, L.W.; Ronmark, E.; et al. The Relevance of Tick Bites to the Production of IgE Antibodies to the Mammalian Oligosaccharide Galactose-α-1,3-Galactose. J. Allergy Clin. Immunol. 2011, 127, 1286–1293.e6. [Google Scholar] [CrossRef] [Green Version]
- Occi, J.L.; Egizi, A.M.; Robbins, R.G.; Fonseca, D.M. Annotated List of the Hard Ticks (Acari: Ixodida: Ixodidae) of New Jersey. J. Med. Entomol. 2019, 56, 589–598. [Google Scholar] [CrossRef]
- Salman, M.; Estrada-Peña, A. Emerging (Re-Emerging) Tick-Borne Infections and the Dissemination of Ticks. In Ticks and Tick-borne Diseases: Geographical Distribution and Control Strategies in the Euro-Asia Region; CABI: Wallingford, UK, 2013; pp. 1–5. ISBN 978-1-84593-853-6. [Google Scholar]
- Occi, J.; Egizi, A.M.; Goncalves, A.; Fonseca, D.M. New Jersey-Wide Survey of Spotted Fever Group Rickettsia (Proteobacteria: Rickettsiaceae) in Dermacentor variabilis and Amblyomma americanum (Acari: Ixodida: Ixodidae). Am. J. Trop. Med. Hyg. 2020, 103, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Schulze, T.L.; Jordan, R.A. Meteorologically Mediated Diurnal Questing of Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae) Nymphs. J. Med. Entomol. 2003, 40, 395–402. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Life Stages | Amblyomma americanum | Dermacentor variabilis | Haemaphysalis longicornis | Ixodes scapularis | Total Ticks |
|---|---|---|---|---|---|
| Larvae | 887 (86%) | 0 | 8078 (89%) | 647 (81%) | 9612 (88%) |
| Nymphs | 95 (9%) | 0 | 819 (9%) | 41 (5%) | 955 (9%) |
| Adults | 50 (5%) | 10 (100%) | 151 (2%) | 106 (13%) | 317 (3%) |
| Total | 1032 | 10 | 9048 | 794 | 10,884 |
| Site | Amblyomma americanum | Haemaphysalis longicornis | Ixodes scapularis | Total Ticks |
|---|---|---|---|---|
| 1 | 0.022 | 0.039 a | 0.176 ab | 0.236 |
| 2 | 0.147 | 0.361 ab | 0.070 ab | 0.579 |
| 3 | 0.196 | 0.845 ab | 0.113 a | 1.154 |
| 4 | 0.101 | 0.459 ab | 0.004 b | 0.564 |
| 5 | 0.010 | 2.198 b | 0.026 ab | 2.238 |
| Statistic p-value | 3.90 (KW) 0.4196 | 10.45 (KW) 0.0335 * | 10.43 (KW) 0.0338 * | 1.79 (F) 0.1537 |
| Tick Species | Life Stage | Peak of Tick Activity | N | Temperature | Relative Humidity | Saturation Deficit | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| rs | 95% CI | p-Value | rs | 95% CI | p-Value | rs | 95% CI | p-Value | ||||
| Haemaphysalis longicornis | L | 18 August to 21 October | 48 | −0.092 | −0.374 to 0.205 | 0.5332 | −0.040 | −0.329 to 0.255 | 0.785 | −0.008 | −0.299 to 0.285 | 0.956 |
| N | 6 April to 16 June | 40 | 0.004 | −0.316 to 0.324 | 0.978 | 0.355 | 0.0395 to 0.606 | 0.024 * | −0.126 | −0.429 to 0.202 | 0.439 | |
| A | 15 July to 1 September | 40 | 0.351 | 0.035 to 0.603 | 0.026 * | −0.339 | −0.595 to −0.021 | 0.032 * | 0.369 | 0.056 to 0.617 | 0.019 * | |
| Amblyomma americanum | L | 29 July to 1 September | 30 | 0.303 | −0.075 to 0.605 | 0.104 | −0.140 | −0.485 to 0.242 | 0.459 | 0.207 | −0.176 to 0.536 | 0.272 |
| N | 3 May to 6 July | 41 | 0.220 | −0.103 to 0.502 | 0.166 | 0.251 | −0.070 to 0.525 | 0.113 | 0.155 | −0.169 to 0.449 | 0.332 | |
| A | 16 May to 2 July | 30 | −0.121 | −0.470 to 0.261 | 0.525 | −0.084 | −0.440 to 0.295 | 0.659 | −0.105 | −0.457 to 0.275 | 0.580 | |
| Ixodes scapularis | L | 6 July to 1 September | 45 | 0.029 | −0.275 to 0.328 | 0.849 | −0.018 | −0.318 to 0.285 | 0.908 | 0.004 | −0.297 to 0.306 | 0.976 |
| N | 3 May to 2 July | 36 | 0.139 | −0.208 to 0.455 | 0.420 | 0.112 | −0.234 to 0.433 | 0.515 | 0.068 | −0.276 to 0.396 | 0.693 | |
| A | 21 Oct to 24 November | 30 | 0.549 | 0.225 to 0.764 | 0.002 ** | −0.029 | −0.395 to 0.345 | 0.880 | 0.377 | 0.008 to 0.655 | 0.040 * | |
| Environmental Variables | Haemaphysalis longicornis (N = 40) | Ixodes scapularis (N = 30) | ||
|---|---|---|---|---|
| Nymphs | Adults | Adults | ||
| Temperature | Slope | 0.007 | 0.005 | |
| Std. Error | 0.003 | 0.002 | ||
| R2 | 0.11 | 0.19 | ||
| F (DFn, DFd) | 4.632 (1, 38) | 6.686 (1, 28) | ||
| p-value | 0.038 * | 0.015 * | ||
| Relative humidity | Slope | 0.012 | −0.002 | |
| Std. Error | 0.005 | 0.001 | ||
| R2 | 0.14 | 0.13 | ||
| F (DFn, DFd) | 6.132 (1, 38) | 5.759 (1, 38) | ||
| p-value | 0.018 * | 0.021 * | ||
| Saturation deficit | Slope | 0.005 | 0.010 | |
| Std. Error | 0.002 | 0.004 | ||
| R2 | 0.14 | 0.17 | ||
| F (DFn, DFd) | 6.283 (1, 38) | 5.565 (1, 28) | ||
| p-value | 0.017 * | 0.025 * | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, J.; Fonseca, D.M.; Toledo, A. Seasonal Dynamics of Tick Species in the Ecotone of Parks and Recreational Areas in Middlesex County (New Jersey, USA). Insects 2023, 14, 258. https://doi.org/10.3390/insects14030258
González J, Fonseca DM, Toledo A. Seasonal Dynamics of Tick Species in the Ecotone of Parks and Recreational Areas in Middlesex County (New Jersey, USA). Insects. 2023; 14(3):258. https://doi.org/10.3390/insects14030258
Chicago/Turabian StyleGonzález, Julia, Dina M. Fonseca, and Alvaro Toledo. 2023. "Seasonal Dynamics of Tick Species in the Ecotone of Parks and Recreational Areas in Middlesex County (New Jersey, USA)" Insects 14, no. 3: 258. https://doi.org/10.3390/insects14030258
APA StyleGonzález, J., Fonseca, D. M., & Toledo, A. (2023). Seasonal Dynamics of Tick Species in the Ecotone of Parks and Recreational Areas in Middlesex County (New Jersey, USA). Insects, 14(3), 258. https://doi.org/10.3390/insects14030258