


The Male Reproductive System of the Kissing Bug, Rhodnius prolixus Stål, 1859 (Hemiptera: Reduviidae: Triatominae): Arrangements of the Muscles and the Myoactivity of the Selected Neuropeptides


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemicals
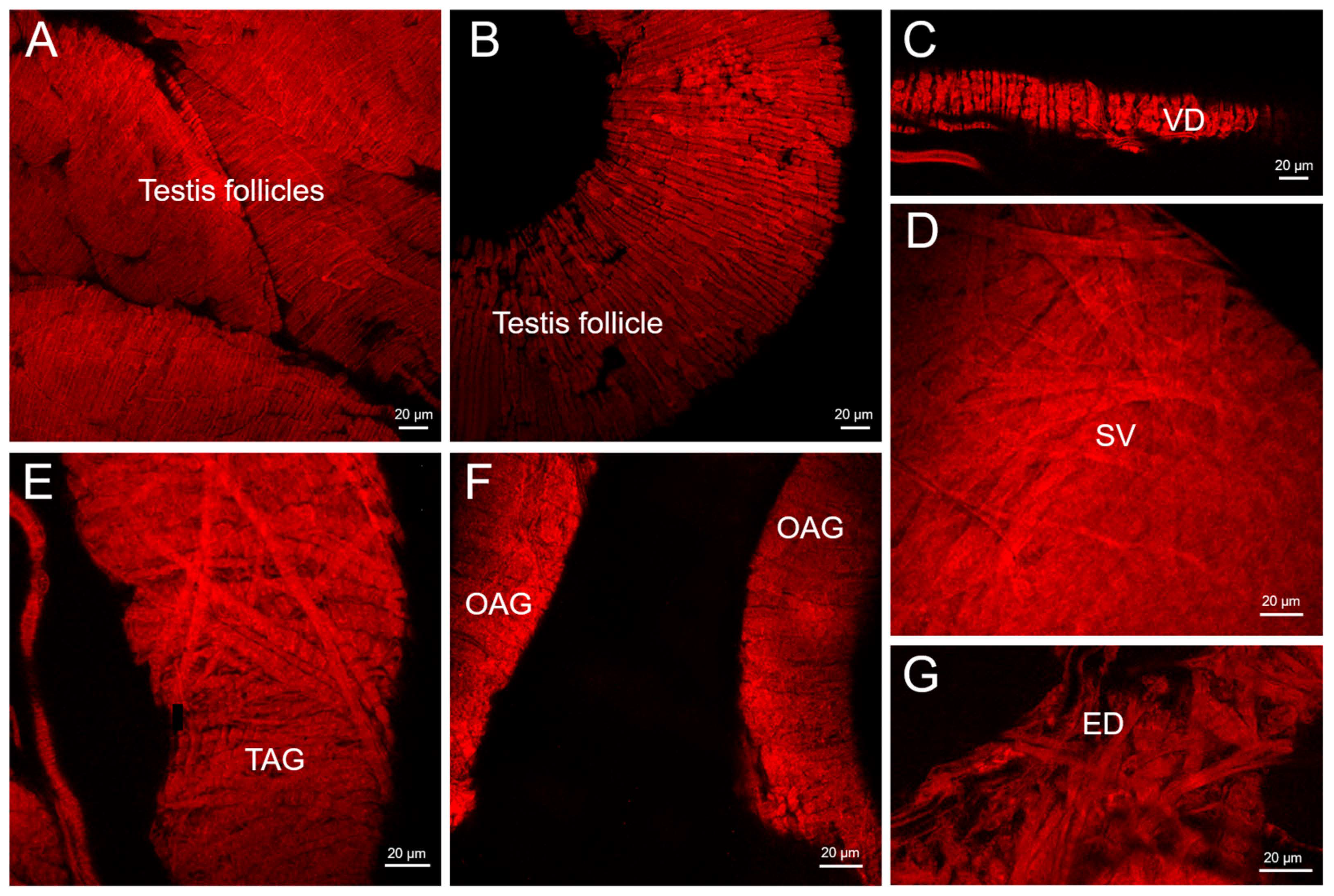
2.3. Phalloidin Staining
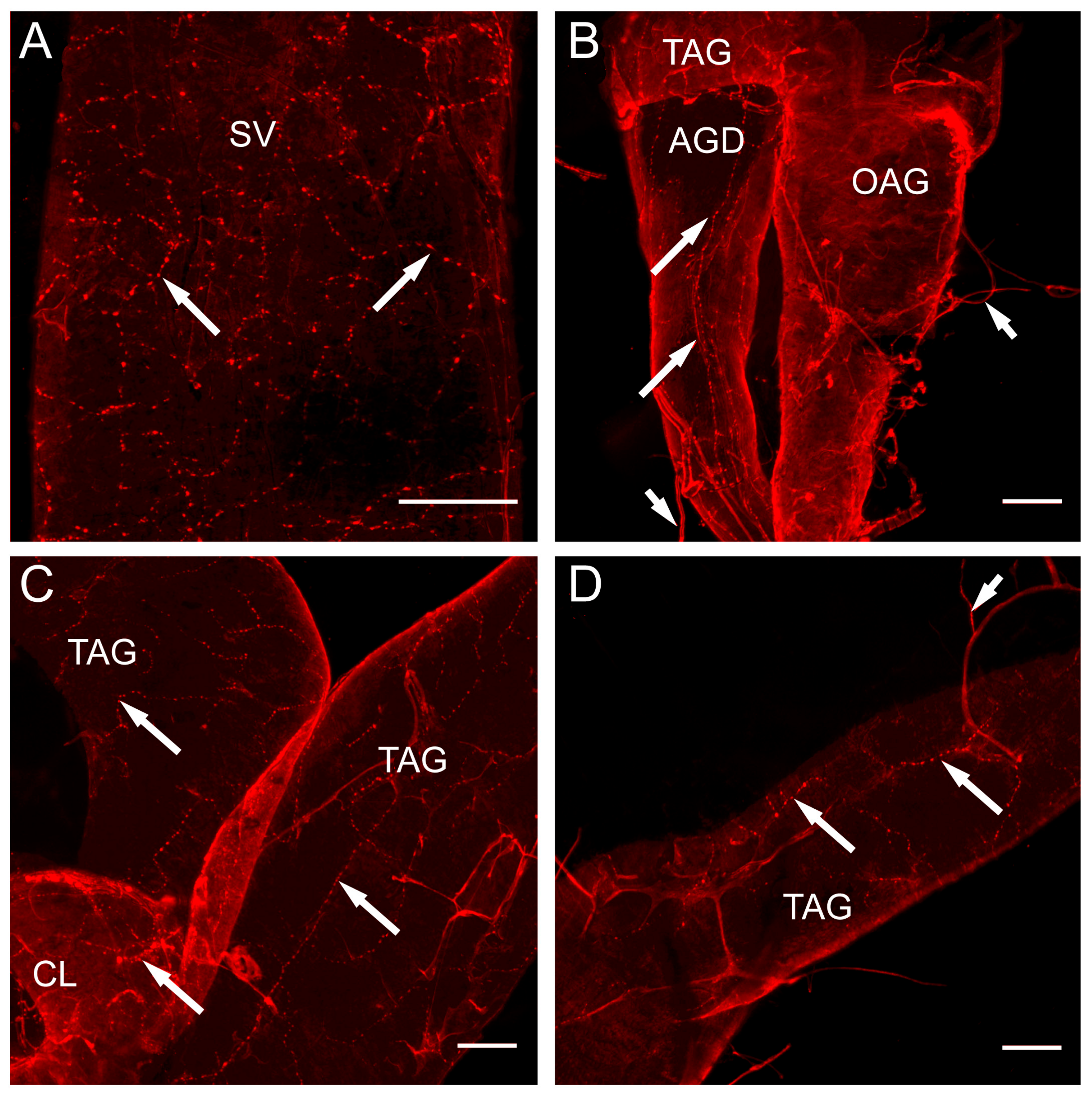
2.4. Whole Mount Immunohistochemistry
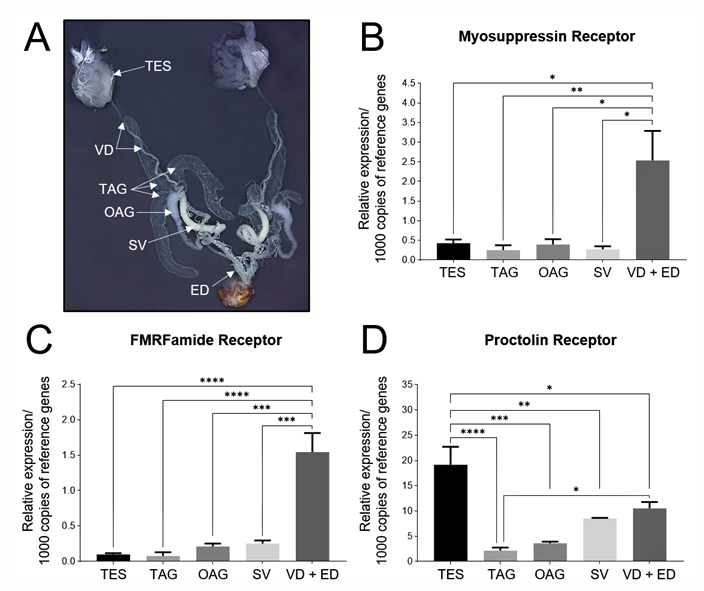
2.5. RNA Extraction and Quantitative Polymerase Chain Reaction (qPCR)
2.6. Contraction Assays
2.7. Statistical Analyses
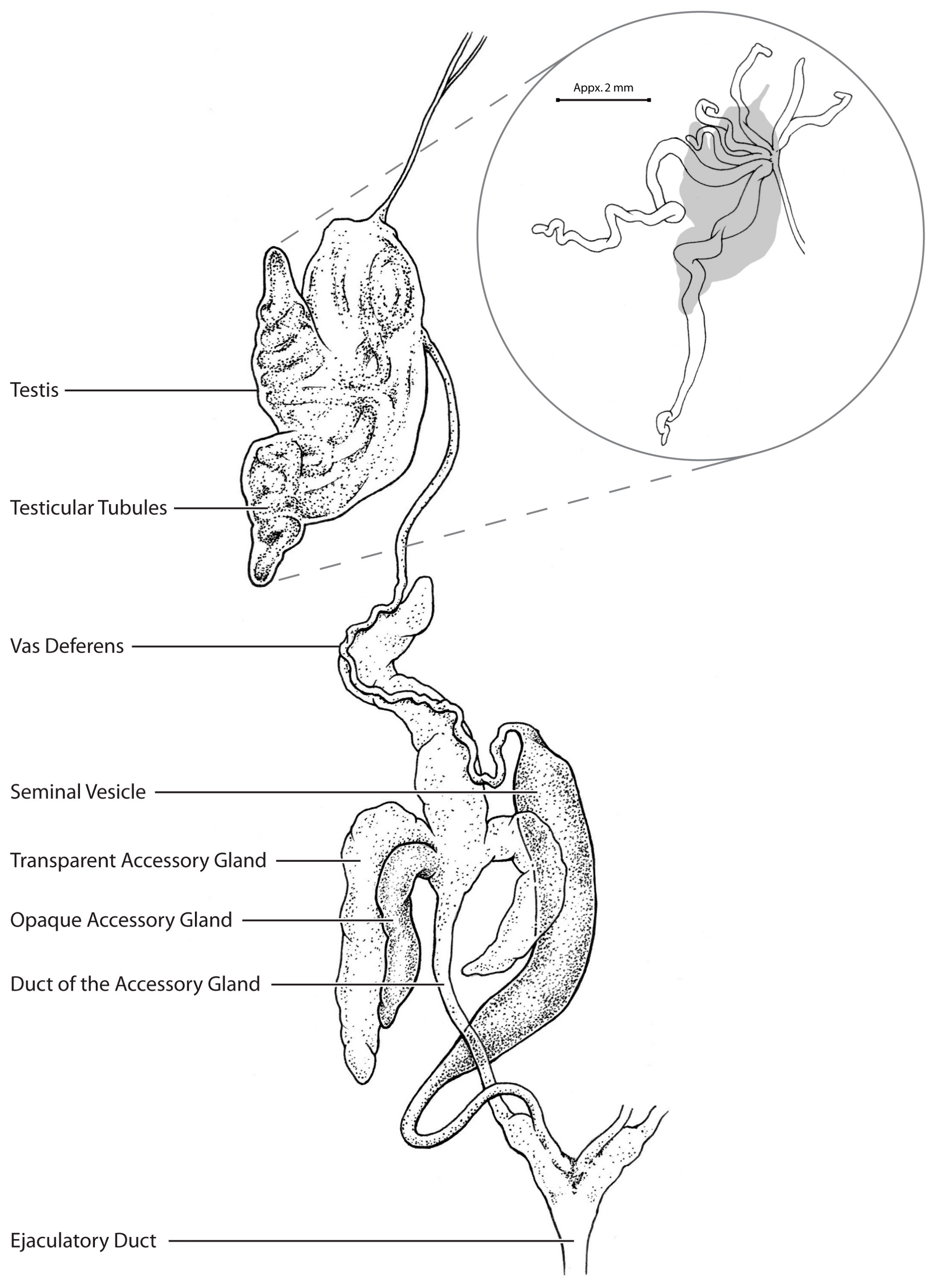
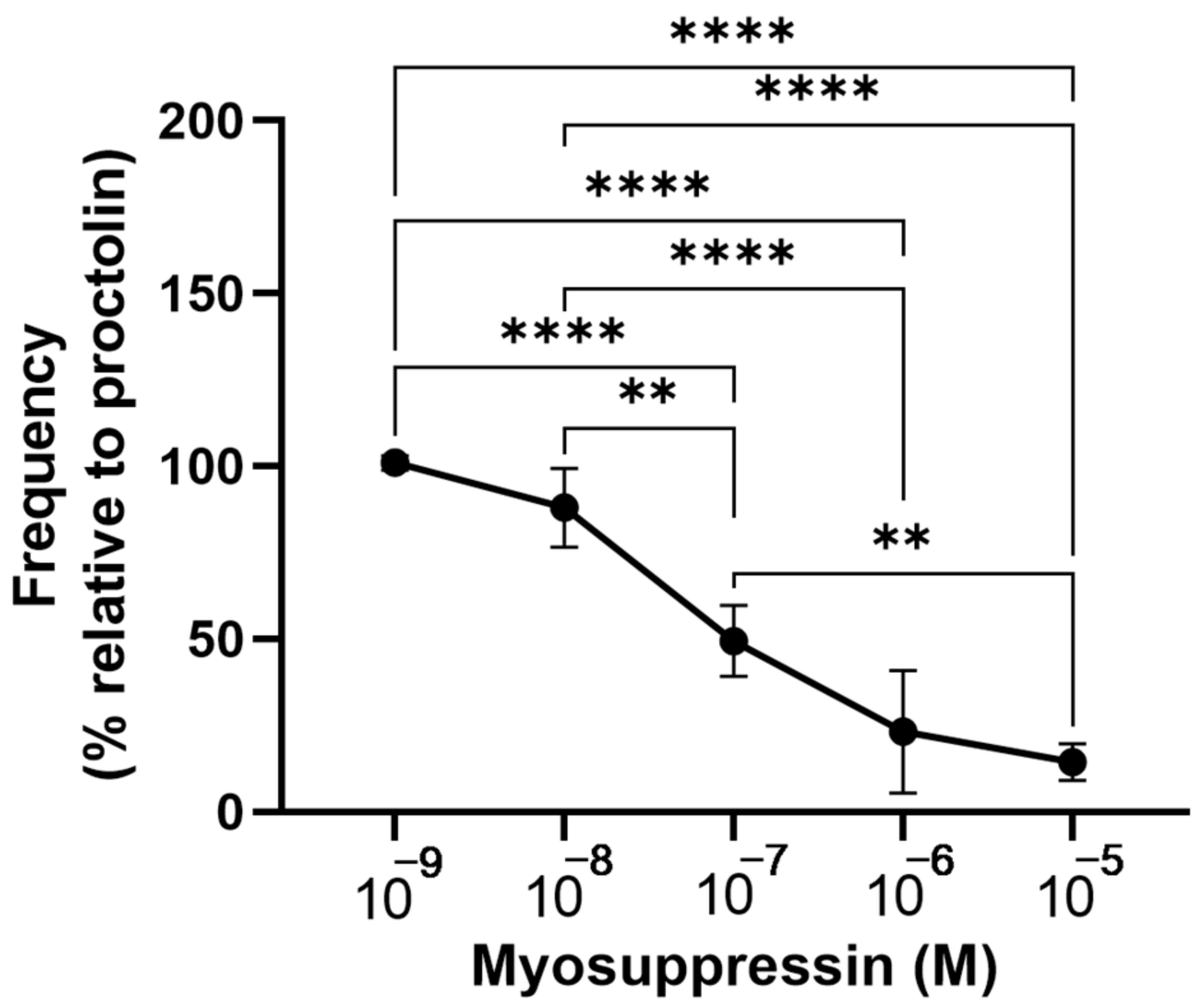
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wigglesworth, V.B. Factors controlling moulting and ‘metamorphosis’ in an insect. Nature 1934, 133, 725–726. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. Source of moulting hormone in Rhodnius. Nature 1951, 168, 558. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. The Principles of Insect Physiology, 7th ed.; John Wiley & Sons Inc.: New York, NY, USA, 1972. [Google Scholar]
- Lange, A.B.; Leyria, J.; Orchard, I. The hormonal and neural control of egg production in the historically important model insect, Rhodnius prolixus: A review, with new insights in this post-genomic era. Gen. Comp. Endocrinol. 2022, 321–322, 114030. [Google Scholar] [CrossRef] [PubMed]
- WHO. Chagas Disease (American Trypanosomiasis). Available online: https://www.who.int/health-topics/chagas-disease#tab=tab_1 (accessed on 28 February 2023).
- Buxton, P.A. The biology of a blood-sucking bug, Rhodnius prolixus. Trans. Ent. Soc. Lond. 1930, 78, 227–256. [Google Scholar] [CrossRef]
- Davey, K.G. The interaction of feeding and mating in the hormonal control of egg production in Rhodnius prolixus. J. Insect Physiol. 2007, 53, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Lange, A.B. The female reproductive system and control of oviposition in the Locusta migratoria migratorioides. Can. J. Zool. 2009, 87, 649–661. [Google Scholar] [CrossRef]
- Lange, A.B. Neural mechanisms coordinating the female reproductive system in the locust. Front. Biosci. 2009, 14, 4401–4415. [Google Scholar] [CrossRef] [Green Version]
- Lange, A.B. A review of the involvement of proctolin as a cotransmitter and local neurohormone in the oviduct of the locust, Locusta migratoria. Peptides 2002, 23, 2063–2070. [Google Scholar] [CrossRef]
- Lange, A.B.; da Silva, R. Neural and hormonal control of muscular activity of the spermatheca in the locust, Locusta migratoria. Peptides 2007, 28, 174–184. [Google Scholar] [CrossRef]
- Anholt, R.R.R.H.; O’Grady, P.; Wolfner, M.F.; Harbison, S.T. Evolution of reproductive behavior. Genetics 2020, 214, 49–73. [Google Scholar] [CrossRef] [Green Version]
- White, M.A.; Chen, D.S.; Wolfner, M.A. She’s got nerve: Roles of octopamine in insect female reproduction. J. Neurogenet. 2021, 35, 132–153. [Google Scholar] [CrossRef]
- Davey, K.G. Copulation and egg-production in Rhodnius prolixus: The role of the spermathecae. J. Exp. Biol. 1965, 42, 373–378. [Google Scholar] [CrossRef]
- Davey, K.G. Some consequences of copulation in Rhodnius prolixus. J. Insect Physiol. 1967, 13, 1629–1636. [Google Scholar] [CrossRef]
- Chiang, R.G.; Chiang, J.A. The Physiology of Sperm Transfer and Egg Production in Vectors of Chagas Disease with Particular Reference to Rhodnius prolixus. In Triatominae—The Biology of Chagas Disease Vectors; Guarneri, A., Lorenzo, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 285–385. [Google Scholar]
- Chiang, R.G.; Chiang, J.A.; Hoogendoorn, H.; Lima, M.M. Exploring the role of Rhodtestolin, A cardio-inhibitor from the testes of Rhodnius prolixus, in relation to the structure and function of reproductive organs in insect vectors of Chagas disease. Insects 2013, 4, 593–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, E.M.; Wolfner, M.F. Seminal proteins but not sperm induce morphological changes in the Drosophila melanogaster female reproductive tract during sperm storage. J. Insect Physiol. 2007, 53, 319–331. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, B.R.; Avila, F.W.; Wolfner, M.F. Insect male reproductive glands and their products. In Encyclopedia of Reproduction, 2nd ed.; Skinner, M.K., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 137–144. [Google Scholar] [CrossRef]
- Freitas, S.P.C.; Gonçalves, T.C.M.; Serrão, J.E.; Costa, J.; Santos-Mallet, J.R. Male reproductive system structure and accessory glands ultrastructure of two species of Triatoma (Hemiptera, Reduviidae, Triatominae). Micron 2010, 41, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Özyurt, N.; Candan, S.; Suludere, Z.; Amutkan, D. Morphology and histology of the male reproductive system in Graphosoma lineatum (Heteroptera: Pentatomidae) based on optical and scanning electron microscopy. J. Entomol. Zool. Stud. 2013, 1, 40–46. [Google Scholar]
- Chiang, R.G.; Chiang, J.A. Functional anatomy of the external and internal reproductive structures in insect vectors of Chagas Disease with particular reference to Rhodnius prolixus. In Biological Control of Pest and Vector Insects; Shields, V.D.C., Ed.; InTech: Rigeka, Croatia, 2017; pp. 299–324. [Google Scholar]
- Alevi, K.C.C.; Oliveira, J.; Rosa, J.A.; Azeredo-Oliveira, M.T.V. Coloration of the testicular peritoneal sheath as a synapomorphy of triatomines (Hemiptera, Reduviidae). Biota Neotrop. 2014, 14, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Alevi, K.C.C.; Castro, N.F.C.; Oliveira, J.; Rosa, J.A.; Azeredo-Oliveira, M.T.V. Cystic spermatogenesis in three species of the prolixus complex (Hemiptera: Triatominae). Ital. J. Zool. 2015, 82, 172–178. [Google Scholar]
- Leopold, R.A. The role of male accessory glands in insect reproduction. Annu. Rev. Entomol. 1976, 21, 199–221. [Google Scholar] [CrossRef]
- Gillott, C. Male accessory gland secretions: Modulators of female reproductive physiology and behavior. Annu. Rev. Entomol. 2003, 48, 163–184. [Google Scholar] [CrossRef] [PubMed]
- Insausti, T.C. Nervous system of Triatoma infestans. J. Morphol. 1994, 221, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Lange, A.B.; Alim, U.; Vandersmissen, H.P.; Mizoguchi, A.; Vanden Broeck, J.; Orchard, I. The distribution and physiological effects of the myoinhibiting peptides in the kissing bug, Rhodnius prolixus. Front. Neurosci. 2012, 6, 98. [Google Scholar] [CrossRef] [Green Version]
- Orchard, I.; Lee, D.H.; da Silva, R.; Lange, A.B. The proctolin gene and biological effects of proctolin in the blood-feeding bug, Rhodnius prolixus. Front. Neuroendocrinol. 2011, 2, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, A.B. The presence of proctolin in the reproductive system of Rhodnius prolixus. J. Insect Physiol. 1990, 36, 345–351. [Google Scholar] [CrossRef]
- Orchard, I.; Lange, A.B.; Bendena, W.G. FMRFamide-related peptides: A multifunctional family of structurally related neuropeptides in insects. Adv. Insect Physiol. 2001, 28, 267–329. [Google Scholar]
- Sedra, L.; Lange, A.B. The female reproductive system of the kissing bug, Rhodnius prolixus: Arrangements of muscles, distribution and myoactivity of two endogenous FMRFamide-like peptides. Peptides 2014, 53, 140–147. [Google Scholar] [CrossRef]
- Marciniak, P.; Urbański, A.; Kudlewska, M.; Szymczak, M.; Rosiński, G. Peptide hormones regulate the physiological functions of reproductive organs in Tenebrio molitor males. Peptides 2017, 98, 35–42. [Google Scholar] [CrossRef]
- Marciniak, P.; Rosiński, G. Comparison of proctolin and FMRFamide actions on the motility of male and female beetle reproductive tracts. Invertebr. Rep. Dev. 2010, 54, 1–6. [Google Scholar] [CrossRef]
- Marciniak, P.; Urbański, A.; Lubawy, J.; Szymczak, M.; Pacholska-Bogalska, J.; Chowański, S.; Kuczer, M.; Rosiński, G. Short neuropeptide F signaling regulates functioning of male reproductive system in Tenebrio molitor beetle. J. Comp. Physiol. 2020, 190, 521–534. [Google Scholar] [CrossRef]
- Al-Dailami, A.N.; Leyria, J.; Orchard, I.; Lange, A.B. Exploring the role of glycoprotein hormone GPA2/GPB5 in the medically important insect, Rhodnius prolixus. Peptides 2022, 149, 170710. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Ons, S.; Sterkel, M.; Diambra, L.; Urlaub, H.; Rivera-Pomar, R. Neuropeptide precursor gene discovery in the Chagas disease vector Rhodnius prolixus. Insect Mol. Biol. 2011, 20, 29–44. [Google Scholar] [CrossRef]
- Ons, S.; Richter, F.; Urlaub, H.; Pomar, R.R. The neuropeptidome of Rhodnius prolixus brain. Proteomics 2009, 9, 788–792. [Google Scholar] [CrossRef]
- Lee, D.; Taufique, H.; da Silva, R.; Lange, A.B. An unusual myosuppressin from the blood-feeding bug, Rhodnius prolixus. J. Exp. Biol. 2012, 215, 2088–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orchard, I.; Lange, A.B. FMRFamide-like peptides (FLPs). In The Handbook of Biologically Active Peptides; Kastin, E., Ed.; Elsevier Publishers: Amsterdam, The Netherlands, 2013; pp. 237–246. [Google Scholar]
- Lee, D.H.; James, T.E.; Lange, A.B. Identification, characterization and expression of a receptor for the unusual myosuppressin in the blood-feeding bug, Rhodnius prolixus. Insect Mol. Biol. 2015, 24, 129–137. [Google Scholar] [CrossRef]
- Lange, A.B.; Orchard, I.; Te Brugge, V.A. Evidence for the involvement of a SchistoFLRF-amide-like peptide in the neural control of locust oviduct. J. Comp. Physiol. 1991, 168, 383–391. [Google Scholar] [CrossRef]
- Starratt, A.N.; Brown, B.E. Structure of pentapeptide proctolin, a proposed neurotransmitter in insects. Life Sci. 1975, 17, 1253–1256. [Google Scholar] [CrossRef]
- Kimura, T.; Yasuyama, K.; Yamaguchi, T. Proctolingergic innervation of the accessory gland in male crickets (Gryllus bimaculatus): Detection of proctolin and some pharmacological properties of myogenically and neurogenically evoked contractions. J. Insect Physiol. 1989, 35, 251–264. [Google Scholar] [CrossRef]
- Nässel, D.R.; Winther, A.M.E. Drosophila neuropeptides in regulation of physiology and behavior. Prog. Neurobiol. 2010, 92, 42–104. [Google Scholar] [CrossRef] [PubMed]
- Orchard, I.; Belanger, J.H.; Lange, A.B. Proctolin: A review with emphasis on insects. J. Neurobiol. 1989, 20, 470–496. [Google Scholar] [CrossRef] [Green Version]
- Lange, A.B.; Orchard, I. Proctolin. In The Handbook of Biologically Active Peptides; Kastin, E., Ed.; Elsevier Publishers: Amsterdam, The Netherlands, 2013; pp. 304–309. [Google Scholar] [CrossRef] [Green Version]
- Mesquita, R.D.; Vionette-Amaral, R.J.; Lowenberger, C.; Rivera-Pomar, R.; Monteiro, F.A.; Minx, P.; Spieth, J.; Carvalho, A.B.; Panzera, F.; Lawson, D.; et al. Genome of Rhodnius prolixus, an insect vector of Chagas disease, reveals unique adaptations to hematophagy and parasite infection. Proc. Natl. Acad. Sci. USA 2015, 112, 14936–14941. [Google Scholar] [CrossRef] [Green Version]
- Davey, K.G. Spermatophore production in Rhodnius prolixus. J. Cell Sci. 1959, s3-100, 221–230. [Google Scholar] [CrossRef]
- Cook, B.J.; Meola, S. Nerve-muscle networks in the accessory gland tubules of a male insect: Structural and physiological properties. J. Insect Physiol. 1987, 33, 791–801. [Google Scholar] [CrossRef]
- Rankin, S.M.; TeBrugge, V.A.; Murray, J.A.; Schuler, A.M.; Tobe, S.S. Effects of selected neuropeptides, mating status and castration on male reproductive tract movements and immunolocalization of neuropeptides in earwigs. Comp. Biochem. Physiol. 2009, 152, 83–90. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lange, A.B.; Kisana, A.; Leyria, J.; Orchard, I. The Male Reproductive System of the Kissing Bug, Rhodnius prolixus Stål, 1859 (Hemiptera: Reduviidae: Triatominae): Arrangements of the Muscles and the Myoactivity of the Selected Neuropeptides. Insects 2023, 14, 324. https://doi.org/10.3390/insects14040324
Lange AB, Kisana A, Leyria J, Orchard I. The Male Reproductive System of the Kissing Bug, Rhodnius prolixus Stål, 1859 (Hemiptera: Reduviidae: Triatominae): Arrangements of the Muscles and the Myoactivity of the Selected Neuropeptides. Insects. 2023; 14(4):324. https://doi.org/10.3390/insects14040324
Chicago/Turabian StyleLange, Angela B., Anika Kisana, Jimena Leyria, and Ian Orchard. 2023. "The Male Reproductive System of the Kissing Bug, Rhodnius prolixus Stål, 1859 (Hemiptera: Reduviidae: Triatominae): Arrangements of the Muscles and the Myoactivity of the Selected Neuropeptides" Insects 14, no. 4: 324. https://doi.org/10.3390/insects14040324
APA StyleLange, A. B., Kisana, A., Leyria, J., & Orchard, I. (2023). The Male Reproductive System of the Kissing Bug, Rhodnius prolixus Stål, 1859 (Hemiptera: Reduviidae: Triatominae): Arrangements of the Muscles and the Myoactivity of the Selected Neuropeptides. Insects, 14(4), 324. https://doi.org/10.3390/insects14040324