
Functional Characterization of the Nuclear Receptor Gene SaE75 in the Grain Aphid, Sitobion avenae


 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Aphid Sampling and Colony Maintenance
2.2. Cloning of SaE75 Gene and Sequence Analysis
2.3. Temporal Differences in SaE75 Expression between Winged and Wingless Morphs
2.4. Treatment with 20E Analogue
2.5. In Vivo Characterization of SaE75 RNAi on Molting
2.6. Statistical Analysis
3. Results
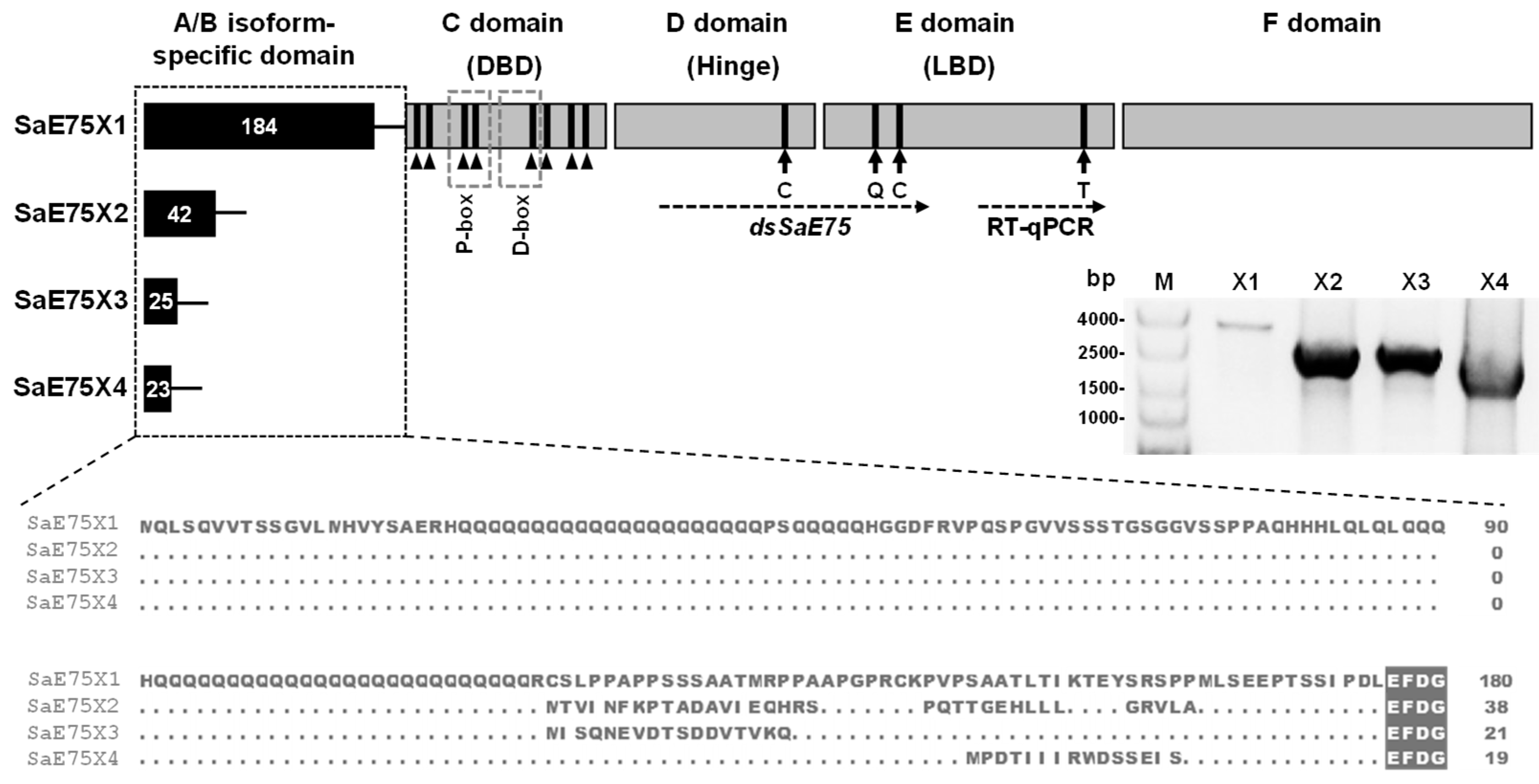
3.1. Molecular Cloning and Characterization of SaE75
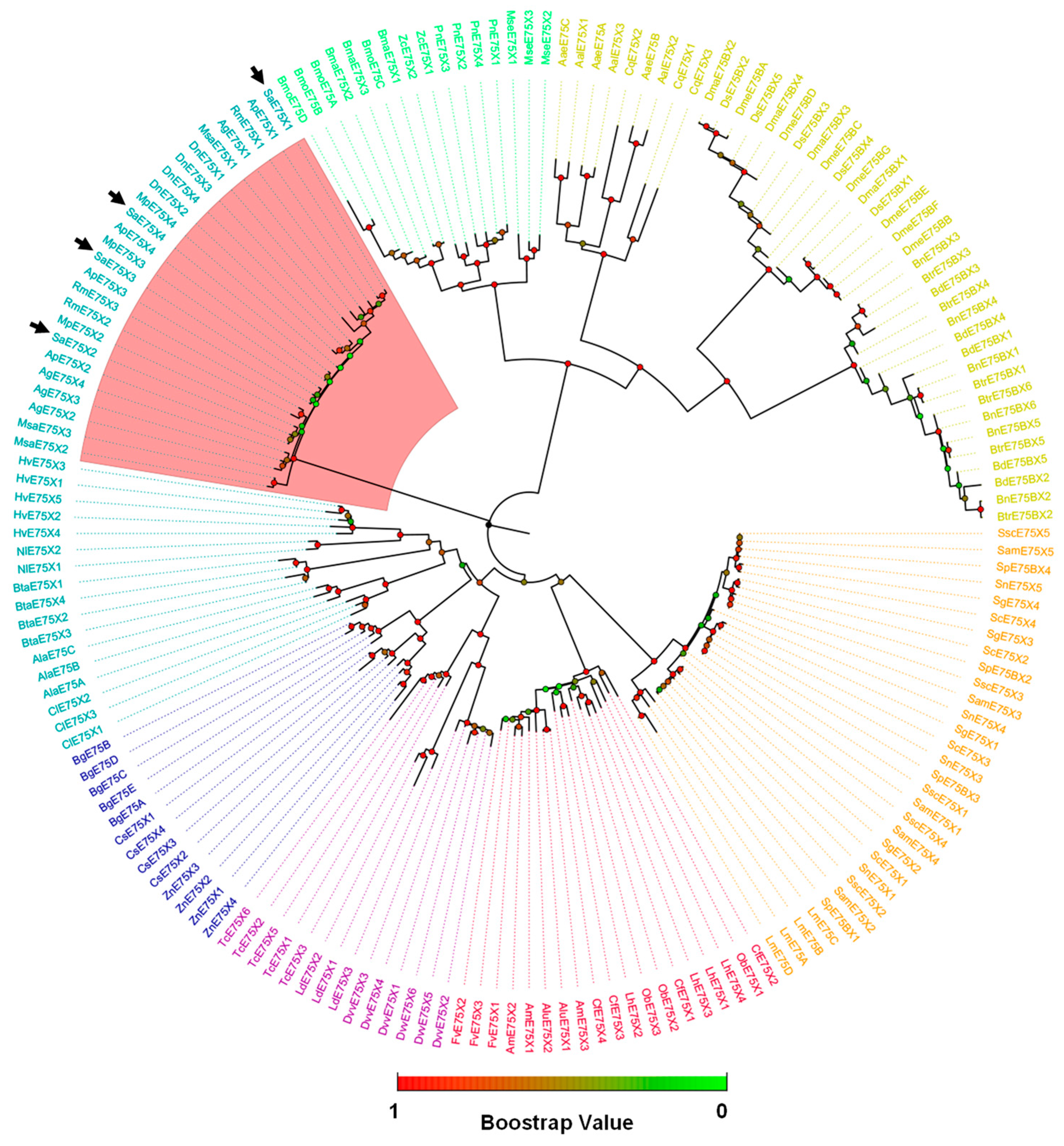
3.2. Phylogenetic Analysis
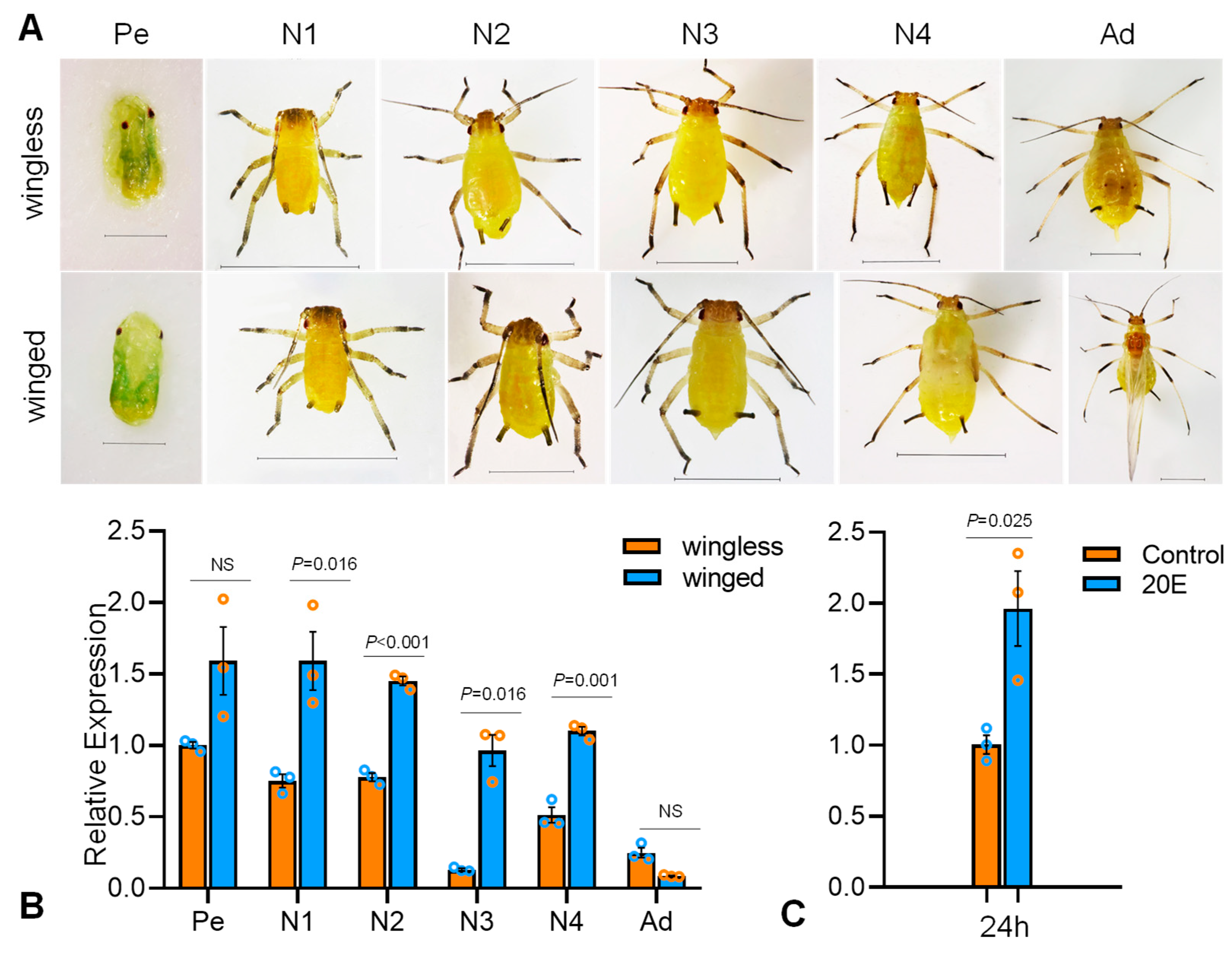
3.3. Expression Characteristics of SaE75
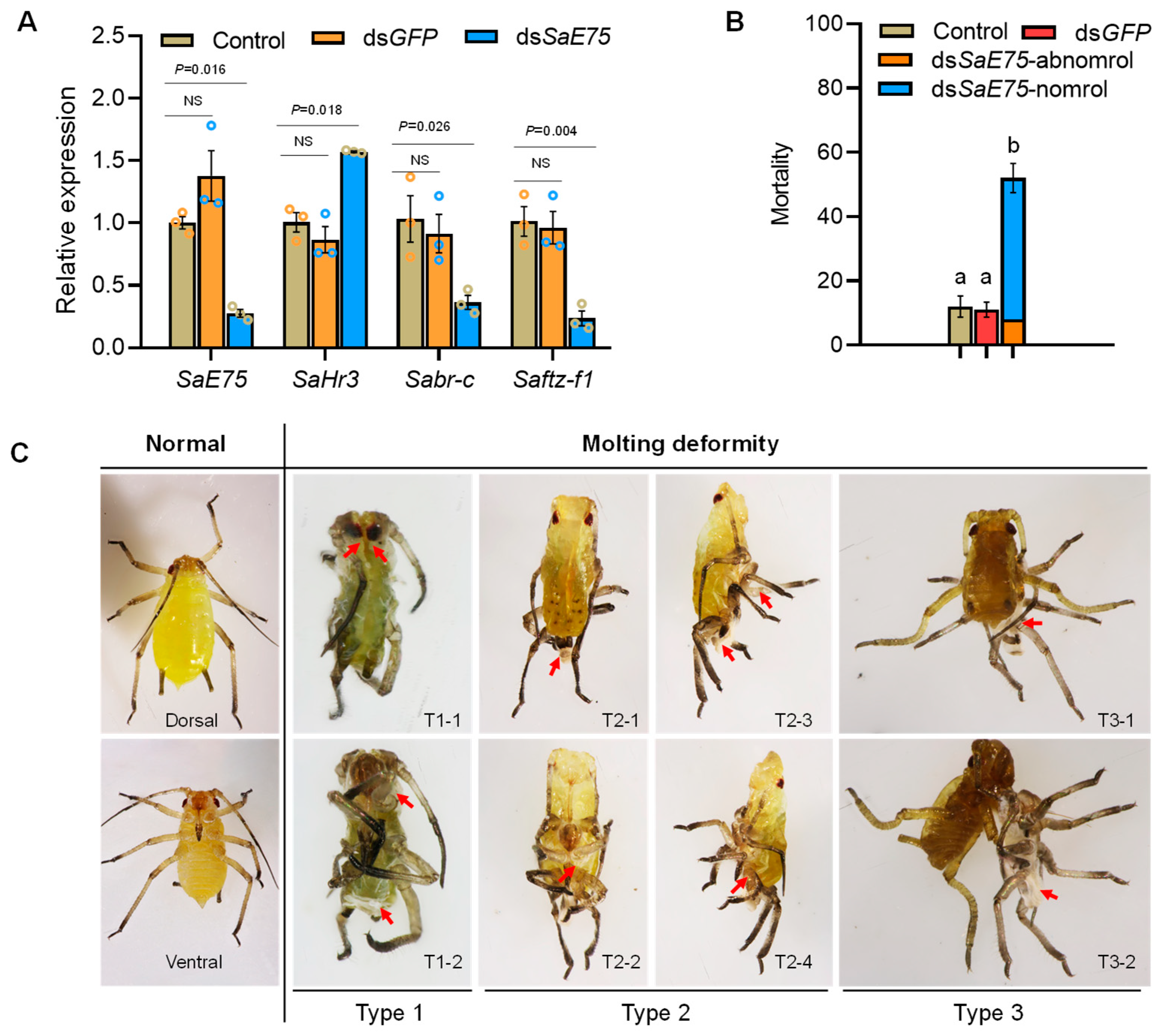
3.4. In Vivo Validation of SaE75 Impacts on Molting
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wenger, J.A.; Cassone, B.J.; Legeai, F.; Johnston, J.S.; Bansal, R.; Yates, A.D.; Coates, B.S.; Pavinato, V.A.C.; Michel, A. Whole genome sequence of the soybean aphid, Aphis glycines. Insect Biochem. Mol. Biol. 2020, 123, 102917. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, J.T. Aphids. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Academic Press: Cambridge, MA, USA, 2009; pp. 27–31. ISBN 978-0-12-374144-8. [Google Scholar]
- Brisson, J.A. Aphid wing dimorphisms: Linking environmental and genetic control of trait variation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 605–616. [Google Scholar] [CrossRef] [Green Version]
- Brisson, J.A.; Jaquiery, J.; Legeai, F.; Le Trionnaire, G.; Tagu, D. Genomics of phenotypic plasticity in aphids. In Management of Insect Pests to Agriculture, 2nd ed.; Czosnek, H., Ghanim, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 65–96. ISBN 978-3-319-24047-3. [Google Scholar]
- Harrison, J.S.; Mondor, E.B. Evidence for an invasive aphid “superclone”: Extremely low genetic diversity in Oleander aphid (Aphis nerii) populations in the southern United States. PLoS ONE 2011, 6, e17524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, P.; Li, X.; Gao, H.; Wang, C.; Li, M.; Zhang, Y.; Li, X.; Liu, E.; Zhu, X. Field evolved resistance to pyrethroids, neonicotinoids, organophosphates and macrolides in Rhopalosiphum padi (Linnaeus) and Sitobion avenae (Fabricius) from China. Chemosphere 2021, 269, 128747. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Gao, X. Multiple mechanisms responsible for differential susceptibilities of Sitobion avenae (Fabricius) and Rhopalosiphum padi (Linnaeus) to pirimicarb. Bull. Entomol. Res. 2009, 99, 611–617. [Google Scholar] [CrossRef]
- Zhang, F.; Li, X.; Zhang, Y.; Coates, B.; Zhou, X.J.; Cheng, D. Bacterial symbionts, Buchnera, and starvation on wing dimorphism in English grain aphid, Sitobion avenae (F.) (Homoptera: Aphididae). Front. Physiol. 2015, 6, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisson, J.A.; Ishikawa, A.; Miura, T. Wing development genes of the pea aphid and differential gene expression between winged and unwinged morphs. Insect Mol. Biol. 2010, 19, 63–73. [Google Scholar] [CrossRef]
- Vellichirammal, N.N.; Gupta, P.; Hall, T.A.; Brisson, J.A. Ecdysone signaling underlies the pea aphid transgenerational wing polyphenism. Proc. Natl. Acad. Sci. USA 2017, 114, 1419–1423. [Google Scholar] [CrossRef] [Green Version]
- Dubrovsky, E.B. Hormonal cross talk in insect development. Trends Endocrinol. Metab. 2005, 16, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Harmon, M.A.; Laudet, V.; Mangelsdorf, D.J.; Palmer, M.J. Isolation of a functional ecdysteroid receptor homologue from the ixodid tick Amblyomma americanum (L.). Insect Biochem. Mol. Biol. 1997, 27, 945–962. [Google Scholar] [CrossRef]
- Christiaens, O.; Iga, M.; Velarde, R.A.; Rougé, P.; Smagghe, G. Halloween genes and nuclear receptors in ecdysteroid biosynthesis and signalling in the pea aphid. Insect Mol. Biol. 2010, 19, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Buzón, V.; Carbó, L.R.; Estruch, S.B.; Fletterick, R.J.; Estébanez-Perpiñá, E. A conserved surface on the ligand binding domain of nuclear receptors for allosteric control. Mol. Cell. Endocrinol. 2012, 348, 394–402. [Google Scholar] [CrossRef]
- Robinson-Rechavi, M.; Escriva Garcia, H.; Laudet, V. The nuclear receptor superfamily. J. Cell Sci. 2003, 116, 585–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuclear Receptors Nomenclature Committee. A unified nomenclature system for the nuclear receptor superfamily. Cell 1999, 97, 161–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germain, P.; Staels, B.; Dacquet, C.; Spedding, M.; Laudet, V. Overview of nomenclature of nuclear receptors. Pharmacol. Rev. 2006, 58, 685–704. [Google Scholar] [CrossRef] [PubMed]
- Bialecki, M.; Shilton, A.; Fichtenberg, C.; Segraves, W.A.; Thummel, C.S. Loss of the ecdysteroid-inducible E75A orphan nuclear receptor uncouples molting from metamorphosis in Drosophila. Dev. Cell 2002, 3, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Segraves, W.A.; Hogness, D.S. The E75 ecdysone-inducible gene responsible for the 75B early puff in Drosophila encodes two new members of the steroid receptor superfamily. Genes Dev. 1990, 4, 204–219. [Google Scholar] [CrossRef] [Green Version]
- Dubrovskaya, V.A.; Berger, E.M.; Dubrovsky, E.B. Juvenile hormone regulation of the E75 nuclear receptor is conserved in Diptera and Lepidoptera. Gene 2004, 340, 171–177. [Google Scholar] [CrossRef]
- Priya, T.A.; Li, F.; Zhang, J.; Yang, C.; Xiang, J. Molecular characterization of an ecdysone inducible gene E75 of Chinese shrimp Fenneropenaeus chinensis and elucidation of its role in molting by RNA interference. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 156, 149–157. [Google Scholar] [CrossRef]
- Guo, W.C.; Liu, X.P.; Fu, K.Y.; Shi, J.F.; Lü, F.G.; Li, G.Q. Nuclear receptor ecdysone-induced protein 75 is required for larval-pupal metamorphosis in the Colorado potato beetle Leptinotarsa decemlineata (Say). Insect Mol. Biol. 2016, 25, 44–57. [Google Scholar] [CrossRef]
- Li, K.; Tian, L.; Guo, Z.; Guo, S.; Zhang, J.; Gu, S.H.; Palli, S.R.; Cao, Y.; Li, S. 20-Hydroxyecdysone (20E) primary response gene E75 isoforms mediate steroidogenesis autoregulation and regulate developmental timing in Bombyx. J. Biol. Chem. 2016, 291, 18163–18175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mané-Padrós, D.; Cruz, J.; Vilaplana, L.; Pascual, N.; Bellés, X.; Martín, D. The nuclear hormone receptor BgE75 links molting and developmental progression in the direct-developing insect Blattella germanica. Dev. Biol. 2008, 315, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, F.; Coates, B.; Wei, C.; Zhu, X.; Zhang, Y.; Zhou, X. Temporal analysis of microRNAs associated with wing development in the English grain aphid, Sitobion avenae (F.) (Homoptera: Aphidiae). Insect Biochem. Mol. Biol. 2022, 142, 103579. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative Toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Li, X.; Li, X.; Yan, W.; Coates, B.; Zhou, X.; Wang, C.; Haifeng, G.; Zhang, Y.; Zhu, X. Selection of reference genes for RT-qPCR analysis of wing dimorphism in English grain aphid, Sitobion avenae (Hemiptera: Aphididae). J. Econ. Entomol. 2022, 115, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ma, T.T.; Yang, Z.L.; Chang, M.L.; Huo, C.Y.; Kan, Y.C.; Li, D.D. Response of miRNAs related to wing differentiation and their predicted target genes to ecdysone and the confirmation of target gene of miR-92a-1-p5 in the pea aphid, Acyrthosiphon pisum (Hemiptera: Aphidae). Acta Entomol. Sin. 2021, 64, 419–427. [Google Scholar]
- Yan, S.; Qian, J.; Cai, C.; Ma, Z.Z.; Li, J.H.; Yin, M.Z.; Ren, B.Y.; Shen, J. Spray method application of transdermal dsRNA delivery system for efficient gene silencing and pest control on soybean aphid Aphis glycines. J. Pest Sci. 2020, 93, 449–459. [Google Scholar] [CrossRef]
- Li, J.H.; Qian, J.; Xu, Y.Y.; Yan, S.; Shen, J.; Yin, M.Z. A facile-synthesized star polycation constructed as a highly efficient gene vector in pest management. ACS Sustain. Chem. Eng. 2019, 7, 6316–6322. [Google Scholar] [CrossRef]
- Parthasarathy, R.; Tan, A.; Bai, H.; Palli, S.R. Transcription factor broad suppresses precocious development of adult structures during larval-pupal metamorphosis in the red flour beetle, Tribolium castaneum. Mech. Dev. 2008, 125, 299–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escriva, H.; Bertrand, S.; Laudet, V. The evolution of the nuclear receptor superfamily. Essays Biochem. 2004, 40, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Cáceres, L.; Necakov, A.S.; Schwartz, C.; Kimber, S.; Roberts, I.J.; Krause, H.M. Nitric oxide coordinates metabolism, growth, and development via the nuclear receptor E75. Genes Dev. 2011, 25, 1476–1485. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.A.; Zhao, X.D.; Sun, Y.; Hao, D.J.; Zhao, J.; Jiang, Y.P.; Bai, L.X.; Xiao, L.B. The nuclear hormone receptor E75A regulates vitellogenin gene (Al-Vg) expression in the mirid bug Apolygus lucorum. Insect Mol. Biol. 2018, 27, 188–197. [Google Scholar] [CrossRef]
- Liu, X.J.; Guo, J.; Zhang, X.Y.; Ma, E.B.; Zhang, J.Z. Molecular characteristics and function analysis of nuclear receptor gene LmE75 in Locusta migratoria. Sci. Agric. Sin. 2020, 53, 2219–2231. [Google Scholar]
- Subramanian, S.; Shankarganesh, K. Insect Hormones (as Pesticides). In Ecofriendly Pest Management for Food Security, 1st ed.; Omkar, Ed.; Academic Press: Pittsburgh, PA, USA, 2016; pp. 613–650. ISBN 978-0-12-803265-7. [Google Scholar]
- Li, K. Functional Analysis of the Genes Involved in Molting and Metamorphosis Signal Pathways in Nilaparvata Lugens; Chinese Academy of Agricultural Sciences: Hangzhou, China, 2016. [Google Scholar]
- Pierceall, W.E.; Li, C.; Biran, A.; Miura, K.; Raikhel, A.S.; Segraves, W.A. E75 expression in mosquito ovary and fat body suggests reiterative use of ecdysone-regulated hierarchies in development and reproduction. Mol. Cell. Endocrinol. 1999, 150, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Swevers, L.; Ito, K.; Iatrou, K. The BmE75 nuclear receptors function as dominant repressors of the nuclear receptor BmHR3A. J. Biol. Chem. 2002, 277, 41637–41644. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.; Yan, Y.; Wei, G.; Merchant, A.; Gu, Y.; Zhou, X.; Zhu, X.; Zhang, Y.; Li, X. Functional Characterization of the Nuclear Receptor Gene SaE75 in the Grain Aphid, Sitobion avenae. Insects 2023, 14, 383. https://doi.org/10.3390/insects14040383
Zheng H, Yan Y, Wei G, Merchant A, Gu Y, Zhou X, Zhu X, Zhang Y, Li X. Functional Characterization of the Nuclear Receptor Gene SaE75 in the Grain Aphid, Sitobion avenae. Insects. 2023; 14(4):383. https://doi.org/10.3390/insects14040383
Chicago/Turabian StyleZheng, Haixia, Yi Yan, Guohua Wei, Austin Merchant, Yaxin Gu, Xuguo Zhou, Xun Zhu, Yunhui Zhang, and Xiangrui Li. 2023. "Functional Characterization of the Nuclear Receptor Gene SaE75 in the Grain Aphid, Sitobion avenae" Insects 14, no. 4: 383. https://doi.org/10.3390/insects14040383
APA StyleZheng, H., Yan, Y., Wei, G., Merchant, A., Gu, Y., Zhou, X., Zhu, X., Zhang, Y., & Li, X. (2023). Functional Characterization of the Nuclear Receptor Gene SaE75 in the Grain Aphid, Sitobion avenae. Insects, 14(4), 383. https://doi.org/10.3390/insects14040383