
CRISPR/Cas9-Mediated Mutagenesis of Abdominal-A and Ultrabithorax in the Asian Corn Borer, Ostrinia furnacalis



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Phylogenetic Analysis of OfAbd-A and OfUbx Genes
2.1.1. Phylogenetic Tree
2.1.2. Multiple Alignment
2.2. Temporal and Spatial Expression Profiles of OfAbd-A and OfUbx
2.2.1. Sample Collection
2.2.2. Reverse Transcription Quantitative Real-Time PCR (RT-qPCR) Analysis
2.3. CRISPR/Cas9-Targeted Mutagenesis in O. furnacalis
2.3.1. Molecular Cloning and Target Selection
2.3.2. Synthesis of Cas9 mRNA and sgRNAs
2.3.3. Embryo Microinjection
2.4. Genomic DNA Extraction to Identify Successful Mutants
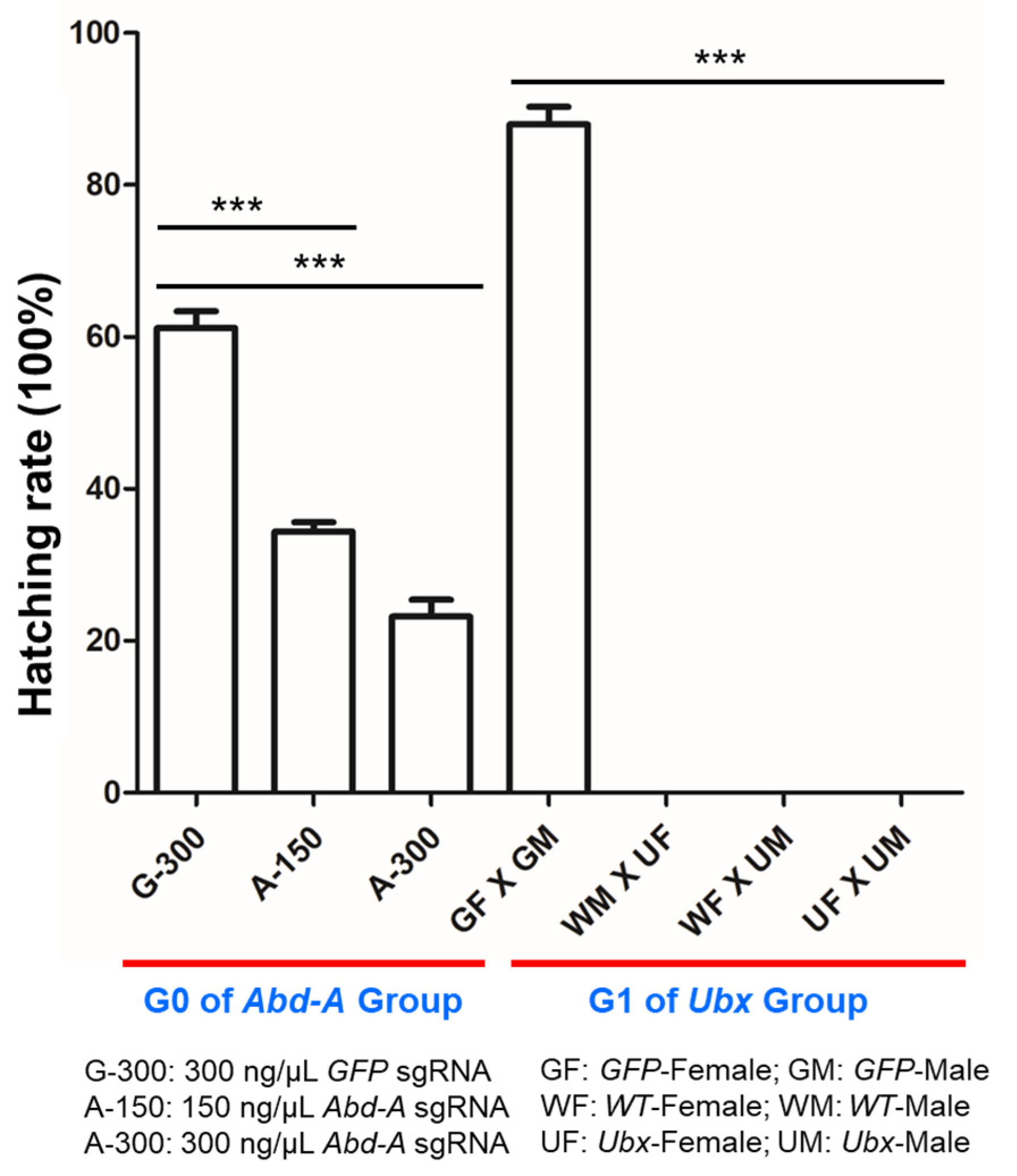
2.5. Analysis of Hatch Rate after Mutagenesis
2.6. Statistical Analysis
3. Results
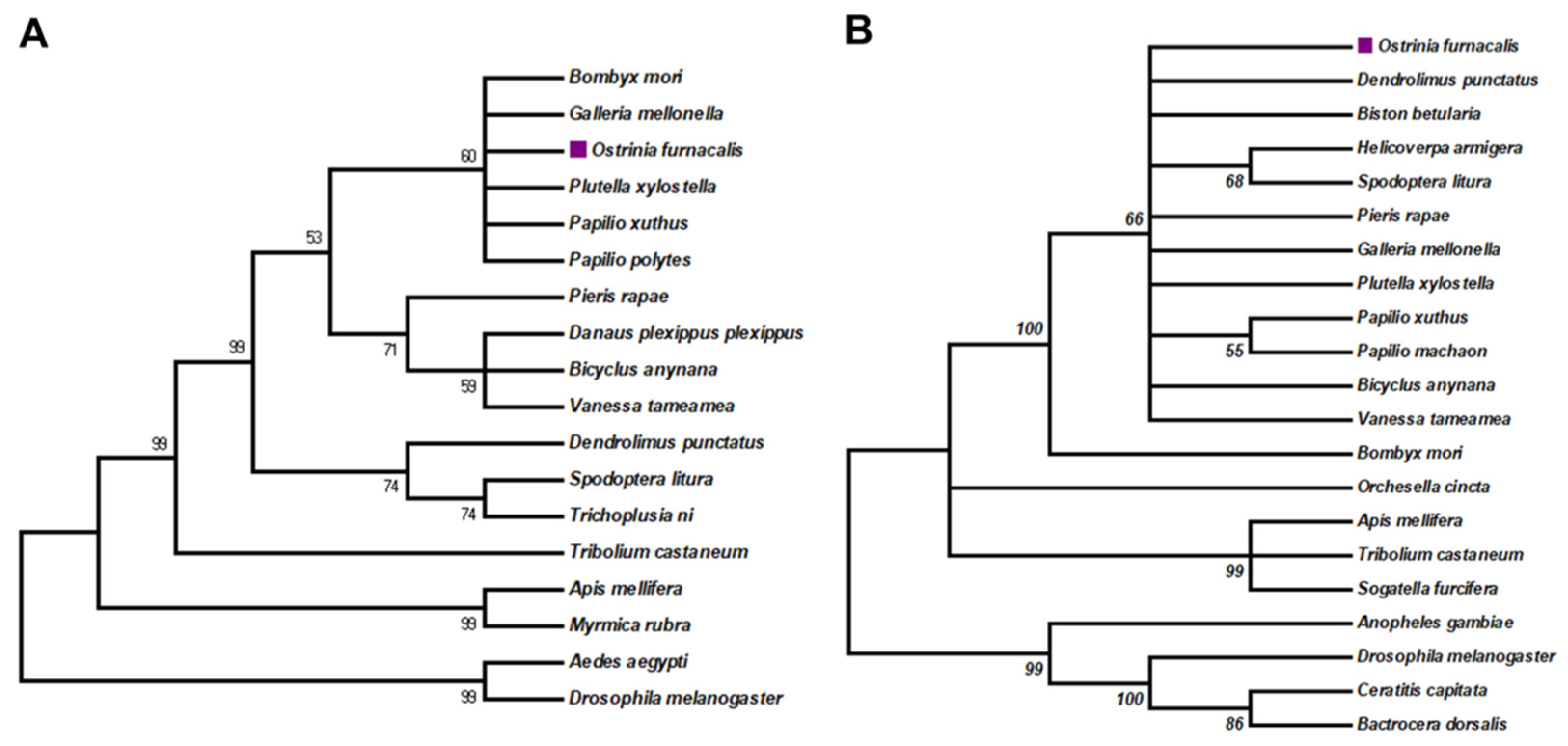
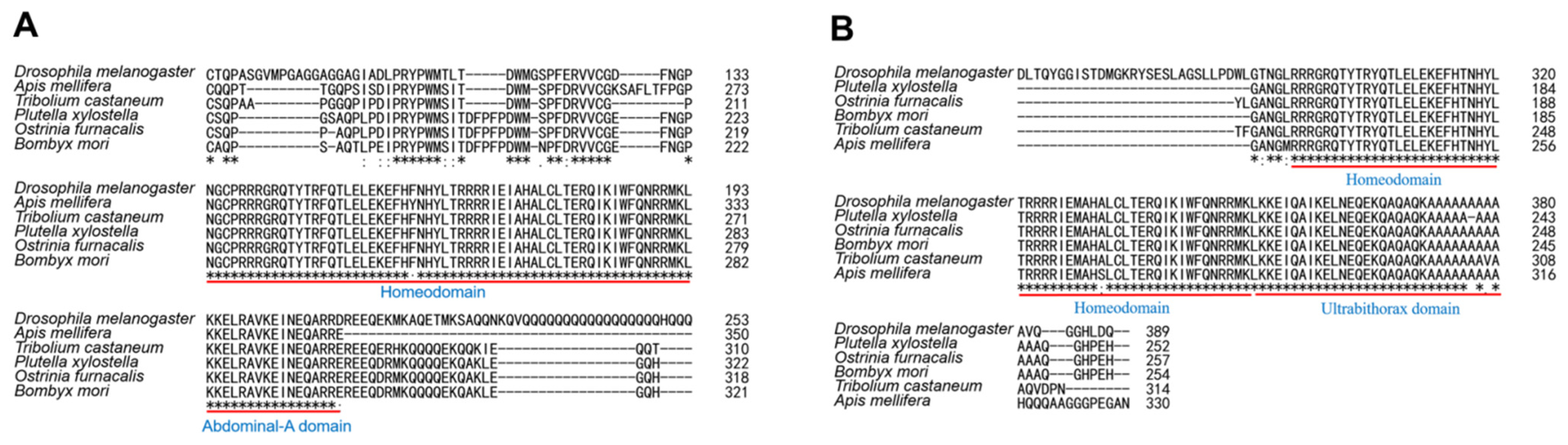
3.1. Phylogenetic Analysis of OfAbd-A and OfUbx
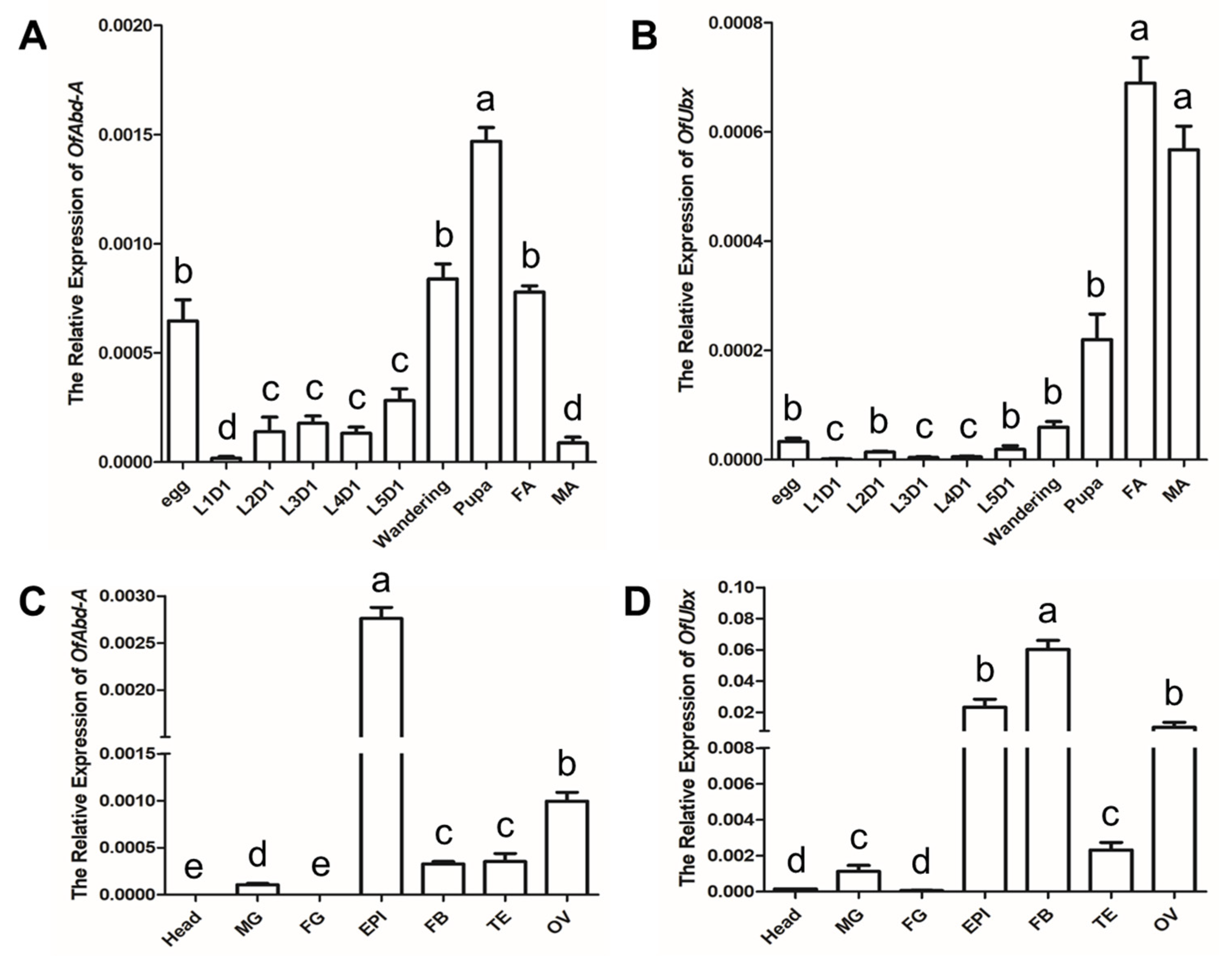
3.2. Temporal–Spatial Distribution of OfAbd-A and OfUbx
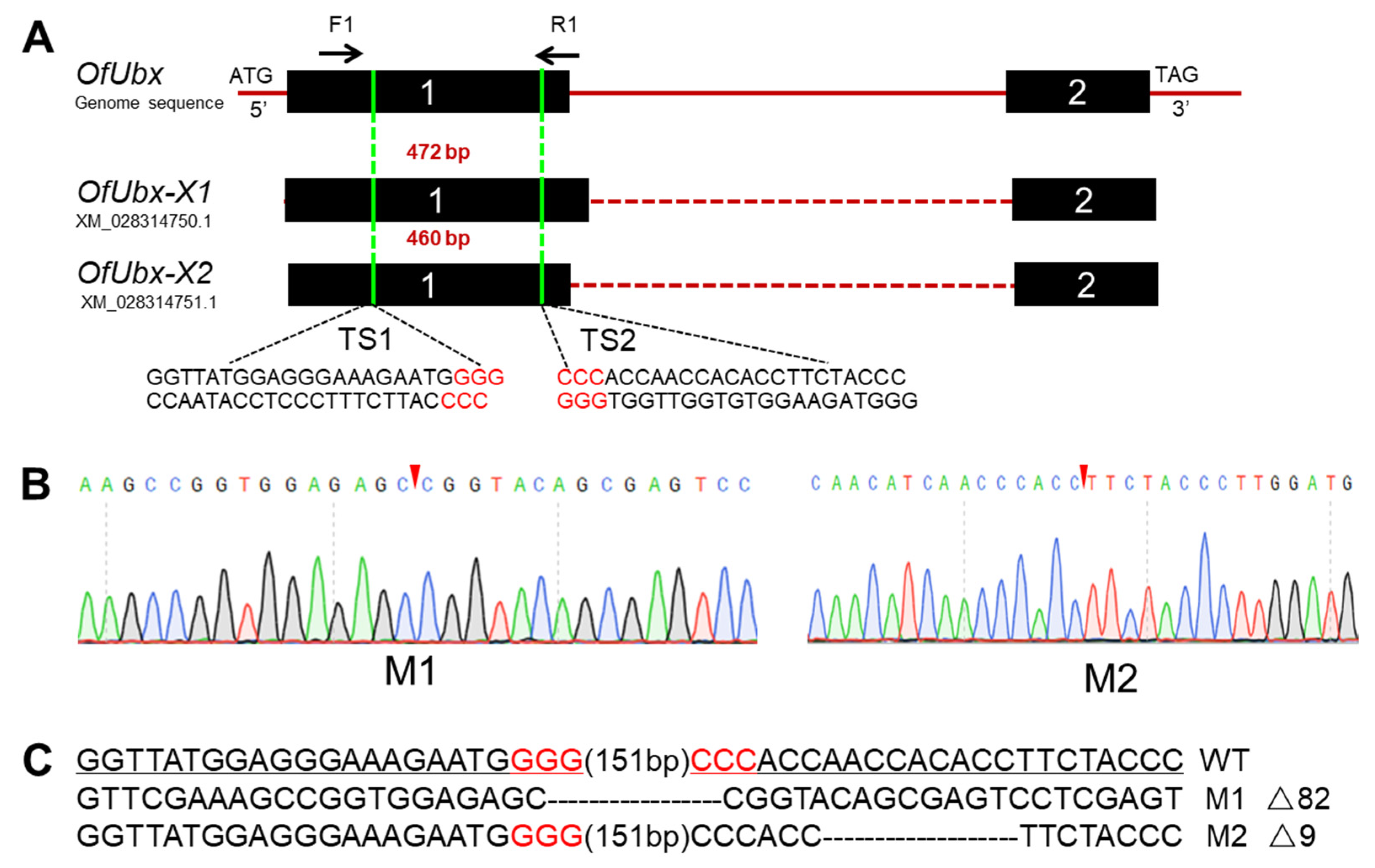
3.3. CRISPR/Cas9-Mediated Mutagenesis in O. furnacalis
3.4. Phenotypic Impacts of OfAbd-A and OfUbx Mutagenesis
3.4.1. Morphological Impacts
3.4.2. Physiological Impacts
3.4.3. Pleiotropic Impacts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nafus, D.M.; Schreiner, I. Review of the biology and control of the Asian corn borer, Ostrinia furnacalis (Lep: Pyralidae). Trop. Pest Manag. 1991, 37, 41–56. [Google Scholar] [CrossRef]
- Afidchao, M.M.; Musters, C.J.; de Snoo, G.R. Asian corn borer (ACB) and non-ACB pests in GM corn (Zea mays L.) in the Philippines. Pest Manag. Sci. 2013, 69, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Wu, Y.; He, K.L.; Bai, S.X. Effects of transgenic Bt maize pollen on longevity and fecundity of Trichogramma ostriniae in laboratory conditions. Bull. Insectol. 2007, 60, 49–55. [Google Scholar]
- Wang, C.L.; Wang, H.X.; Gui, C.M.; Lu, H. Studies on the control of the Asian corn borer, Ostrinia furnacalis (Guenee), with Trichogramma ostriniae. In US. National Acedemy of Science Joint Symposium on Biological Control of Insects; Adkisson, P.L., Ma, S.J., Eds.; Science Press: Beijing, China, 1984; pp. 268–273. [Google Scholar]
- Hassan, S.A.; Guo, M.F. Selection of effective strains of egg parasites of the genus Trichogramma (Hym., Trichogrammatidae) to control the European corn borer Ostrinia nubilalis Hb. (Lep., Pyralidae). J. Appl. Entomol. 1991, 111, 335–341. [Google Scholar] [CrossRef]
- Hoffmann, M.P.; Wright, M.G.; Pitcher, S.A.; Gardner, J. Inoculative releases of Trichogramma ostriniae for suppression of Ostrinia nublilalis (European corn borer) in sweet corn: Field biology and population dynamics. Biol. Control 2002, 25, 249–258. [Google Scholar] [CrossRef]
- Wright, M.G.; Kuhar, T.P.; Hoffmann, M.P.; Chenus, S.A. Effect of inoculative releases of Trichogramma Ostriniae on populations of Ostrinia nubilalis and damage to sweet corn and field corn. Biol. Control 2002, 23, 149–155. [Google Scholar] [CrossRef]
- Munkvold, G.P.; Hellmich, R.L.; Rice, L.G. Comparison of Fumonisin Concentrations in Kernels of Transgenic Bt Maize Hybrids and Nontransgenic Hybrids. Plant Dis. 1999, 83, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Hurley, T.M.; Secchi, S.; Babcock, B.A.; Hellmichet, R.L. Managing the risk of European corn borer resistance to Bt corn. Environ. Resour. Econ. 2002, 22, 537–558. [Google Scholar] [CrossRef]
- Ferry, N.; Edwards, M.G.; Gatehouse, J.; Capell, T.; Christou, P.; Gatehouse, A.M. Transgenic plants for insect pest control: A forward looking scientific perspective. Transgenic Res. 2006, 15, 13–19. [Google Scholar] [CrossRef]
- Thompson, G.D.; Dalmacia, S.C.; Criador, A.R.; Alvarez, E.R.; Hechanova, R.F. Field Performance of TC1507 Transgenic Corn Hybrids Against Asian Corn Borer in the Philippines. Philipp. Agric. Sci. 2010, 93, 375–383. [Google Scholar]
- Xu, L.N.; Wang, Y.Q.; Wang, Z.Y.; Hu, B.J.; Ling, Y.H.; He, K.L. Transcriptome differences between Cry1Ab resistant and susceptible strains of Asian corn borer. BMC Genom. 2015, 16, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crespo, A.L.S.; Spencer, T.A.; Alves, A.P.; Hellmich, R.L.; Blankenship, E.E.; Magalhaes, L.C.; Siegfried, B.D. On-plant survival and inheritance of resistance to Cry1Ab toxin from Bacillus thuringiensis in a field-derived strain of European corn borer, Ostrinia nubilalis. Pest Manag. Sci. 2009, 65, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Crespo, A.L.; Rodrigo-Simón, A.; Siqueira, H.A.; Pereira, E.J.; Ferre, J.; Siegfried, B.D. Cross-resistance and mechanism of resistance to Cry1Ab toxin from Bacillus thuringiensis in a field-derived strain of European corn borer, Ostrinia nubilalis. J. Invert. Pathol. 2011, 107, 185–192. [Google Scholar] [CrossRef]
- Smith, L.J.; Farhan, Y.; Schaafsma, W.A. Practical resistance of Ostrinia nubilalis (Lepidoptera: Crambidae) to Cry1F Bacillus thuringiensis maize discovered in Nova Scotia, Canada. Sci. Rep. 2019, 9, 18247. [Google Scholar] [CrossRef] [Green Version]
- Carroll, S.B. Endless forms: The evolution of gene regulation and morphological diversity. Cell 2000, 101, 577–580. [Google Scholar] [CrossRef] [Green Version]
- Lewis, E.B. A gene complex controlling segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef]
- Krumlauf, R. Hox genes in vertebrate development. Cell 1994, 78, 191–201. [Google Scholar] [CrossRef]
- McGinnis, W.; Krumlauf, R. Homeobox genes and axial patterning. Cell 1992, 68, 283–302. [Google Scholar] [CrossRef]
- Reuter, R.; Scott, M.P. Expression and function of the homoeotic genes Antennapedia and Sex combs reduced in the embryonic midgut of Drosophila. Development 1990, 109, 289–303. [Google Scholar] [CrossRef]
- Maeda, R.K.; Karch, F. The bithorax complex of Drosophila an exceptional Hox cluster. Curr. Top. Dev. Biol. 2009, 88, 1–33. [Google Scholar]
- Liu, F.; Li, X.; Zhao, M.; Guo, M.; Han, K.; Dong, X.; Zhao, J.; Cai, W.; Zhang, Q.; Hua, H. Ultrabithorax is a key regulator for the dimorphism of wings, a main cause for the outbreak of planthoppers in rice. Natl. Sci. Rev. 2020, 7, 1181–1189. [Google Scholar] [CrossRef]
- Lawrence, P.A.; Morata, G. Homeobox genes: Their function in Drosophila segmentation and pattern formation. Cell 1994, 78, 181–189. [Google Scholar] [CrossRef]
- Ponzielli, R.; Astier, M.; Chartier, A.; Gallet, A.; Sémériva, M. Heart tube patterning in Drosophila requires integration of axial and segmental information provided by the Bithorax Complex genes and hedgehog signaling. Development 2002, 129, 4509–4521. [Google Scholar] [CrossRef]
- Vachon, G.; Cohen, B.; Pfeifle, C.; McGuffin, M.E.; Botas, J.; Cohen, S.M. Homeotic genes of the Bithorax complex repress limb development in the abdomen of the Drosophila embryo through the target gene Distal-less. Cell 1992, 71, 437–450. [Google Scholar] [CrossRef]
- Harding, K.; Wedeen, C.; McGinnis, W.; Levine, M. Spatially regulated expression of homeotic genes in Drosophila. Science 1985, 229, 1236–1242. [Google Scholar] [CrossRef]
- Cumberledge, S.U.; Szabad, J.A.; Sakonju, S.H. Gonad formation and development requires the abd-A domain of the bithorax complex in Drosophila melanogaster. Development 1992, 115, 395–402. [Google Scholar] [CrossRef]
- Foronda, D. Requirement of Abdominal-A and Abdominal-B in the developing genitalia of Drosophila breaks the posterior downregulation rule. Development 2006, 133, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, M.F. Differential expression of the Drosophila BX-C in polytene chromosomes in cells of larval fat bodies: A cytological approach to identifying in vivo targets of the homeotic Ubx, Abd-A and Abd-B proteins. Development 2003, 130, 3683–3689. [Google Scholar] [CrossRef] [Green Version]
- Mathies, L.D.; Kerridge, S.; Scott, M.P. Role of the teashirt gene in Drosophila midgut morphogenesis: Secreted proteins mediate the action of homeotic genes. Development 1994, 120, 2799–2809. [Google Scholar] [CrossRef]
- Perrin, L.; Monier, B.; Ponzielli, R.; Astier, M.; Semeriva, M. Drosophila cardiac tube organogenesis requires multiple phases of Hox activity. Dev. Biol. 2004, 272, 419–431. [Google Scholar] [CrossRef] [Green Version]
- Bi, H.L.; Xu, J.; Tan, A.J.; Huang, Y.P. CRISPR/Cas9-mediated targeted gene mutagenesis in Spodoptera litura. Insect Sci. 2016, 23, 469–477. [Google Scholar] [CrossRef]
- Roch, F.; Akam, M. Ultrabithorax and the control of cell morphology in Drosophila halteres. Development 2000, 127, 97–107. [Google Scholar] [CrossRef]
- Averof, M.; Patel, N.H. Crustacean appendage evolution associated with changes in Hox gene expression. Nature 1997, 388, 682–686. [Google Scholar] [CrossRef]
- Kaufman, T.C.; Lewis, R.; Wakimoto, B. Cytogenetic Analysis of Chromosome 3 in Drosophila melanogaster: The Homoeotic Gene Complex in Polytene Chromosome Interval 84a-B. Genetics 1980, 94, 115–133. [Google Scholar] [CrossRef]
- Garcia-Bellido, A. Genetic control of wing disc development in Drosophila. Ciba Found. Symp. 1975, 29, 161–182. [Google Scholar]
- Struhl, G. Genes controlling segmental specification in the Drosophila thorax. Proc. Natl. Acad. Sci. USA 1982, 79, 7380–7384. [Google Scholar] [CrossRef] [Green Version]
- Prasad, N.; Tarikere, S.; Khanale, D.; Habib, F.; Shashidhara, L.S. A comparative genomic analysis of targets of Hox protein Ultrabithorax amongst distant insect species. Sci. Rep. 2016, 6, 27885. [Google Scholar] [CrossRef] [Green Version]
- Gebelein, B.; Culi, J.; Ryoo, H.D.; Zhang, W.; Mann, R.S. Specificity of Distalless repression and limb primordia development by abdominal Hox proteins. Dev. Cell. 2002, 3, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Ronshaugen, M.; McGinnis, N.; McGinnis, W. Hox protein mutation and macroevolution of the insect body plan. Nature 2002, 415, 914–917. [Google Scholar] [CrossRef]
- Pan, M.H.; Wang, X.Y.; Chai, C.L.; Zhang, C.D.; Lu, C.; Xiang, Z.H. Identification and function of Abdominal-A in the silkworm, Bombyx mori. Insect Mol. Biol. 2009, 18, 155–160. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, G.Q.; Zhang, Q.; Chen, X.E.; Cao, Y.H.; Wang, Y.H.; Qi, M.M.; Wu, N.N.; Qian, L.S.; Zhu, C.X.; Huang, Y.P.; et al. The draft genome of the Asian corn borer yields insights into ecological adaptation of a devastating maize pest. Insect Biochem. Mol. Biol. 2021, 138, 103638. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, Y.; Huang, J.; Jin, W.; Yang, Y.; Wu, Y. CRISPR-Mediated Knockout of the ABCC2 Gene in Ostrinia furnacalis Confers High-Level Resistance to the Bacillus thuringiensis Cry1Fa Toxin. Toxins 2020, 12, 246. [Google Scholar] [CrossRef]
- You, L.; Bi, H.L.; Wang, Y.H.; Li, X.W.; Chen, X.E.; Li, Z.Q. CRISPR/Cas9-based mutation reveals Argonaute 1 is essential for pigmentation in Ostrinia furnacalis. Insect Sci. 2019, 26, 1020–1028. [Google Scholar] [CrossRef]
- Merrill, V.K.; Turner, F.R.; Kaufman, T.C. A genetic and developmental analysis of mutations in the Deformed locus in Drosophila melanogaster. Dev. Biol. 1987, 122, 379–395. [Google Scholar] [CrossRef]
- Balavoine, G.; de Rosa, R.; Adoutte, A. Hox clusters and bilaterian phylogeny. Mol. Phylogenet. Evol. 2002, 24, 366–373. [Google Scholar] [CrossRef]
- Duncan, I. The bithorax complex. Annu. Rev. Genet. 1987, 21, 285–319. [Google Scholar] [CrossRef]
- Angelini, D.R.; Liu, P.Z.; Hughes, C.L.; Kaufman, T.C. Hox gene function and interaction in the milkweed bug Oncopeltus fasciatus (Hemiptera). Dev. Biol. 2005, 287, 440–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, C.L.; Kaufman, T.C. Hox genes and the evolution of the arthropod body plan. Evol. Dev. 2002, 4, 459–499. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.C.; Lemons, D.; McGinnis, W. Modulating Hox gene functions during animal body patterning. Nat. Rev. Genet. 2005, 6, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Konopova, B.; Akam, M. The Hox genes Ultrabithorax and abdominal-A specify three different types of abdominal appendage in the springtail Orchesella cincta (Collembola). EvoDevo 2014, 5, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, L.K.; Mahaffey, J.W. Insect homeotic complex genes and development, lessons from Drosophila and beyond. In Comprehensive Insect Science; Elsevier: London, UK, 2005; Volume 1, pp. 247–303. [Google Scholar]
- Chauvet, S.; Maurel-Zaffran, C.; Miassod, R.; Jullien, N.; Pradel, J.; Aragnol, D. dlarp, a new candidate Hox target in Drosophila whose orthologue in mouse is expressed at sites of epithelium/mesenchymal interactions. Dev. Dyn. 2000, 218, 401–413. [Google Scholar] [CrossRef]
- Khila, A.; Abouheif, E.; Rowe, L. Evolution of a novel appendage ground plan in water striders is driven by changes in the Hox gene Ultrabithorax. PLoS Genet. 2009, 5, e1000583. [Google Scholar] [CrossRef] [Green Version]
- Castelli-Gair, J.; Akam, M. How the Hox gene Ultrabithorax specifies two different segments: The significance of spatial and temporal regulation within metameres. Development 1995, 121, 2973–2982. [Google Scholar] [CrossRef]
- Mahfooz, N.S.; Li, H.; Popadić, A. Differential expression patterns of the hox gene are associated with differential growth of insect hind legs. Proc. Natl. Acad. Sci. USA. 2004, 101, 4877–4882. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Yang, F.Y.; Xu, X.J.; Kumar, U.; He, W.Y.; You, M.S. Genetic control of Plutella xylostella in omics era. Arch. Insect Biochem. Physiol. 2019, 102, e21621. [Google Scholar] [CrossRef]
- Paulo, F.D.; Williamson, M.E.; Arp, A.P.; Li, F.; Sagel, A.; Skoda, S.R.; Sanchez-Gallego, J.; Vasquez, M.; Quintero, G.; Pérez de León, A.A.; et al. Specific Gene Disruption in the Major Livestock Pests Cochliomyia hominivorax and Lucilia cuprina Using CRISPR/Cas9. G3-Genes Genomes Genet. 2019, 9, 3045–3055. [Google Scholar] [CrossRef] [Green Version]
- Bi, H.L.; Xu, X.; Li, X.W.; Zhang, Y.; Huang, Y.P.; Li, K.; Xu, J. CRISPR Disruption of BmOvo Resulted in the Failure of Emergence and Affected the Wing and Gonad Development in the Silkworm Bombyx mori. Insects 2019, 10, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, W.Z.; Zhai, Y.Q.; Yang, Y.H.; Wu, Y.D.; Wang, X.L. Cadherin Protein Is Involved in the Action of Bacillus thuringiensis Cry1Ac Toxin in Ostrinia furnacalis. Toxins 2021, 13, 658. [Google Scholar] [CrossRef] [PubMed]
- Weatherbee, S.D.; Nijhout, H.F.; Grunert, L.W.; Halder, G.; Galant, R.; Selegue, J.; Carroll, S. Ultrabithorax function in butterfly wings and the evolution of insect wing patterns. Curr. Biol. 1999, 9, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, J. Hox and Wings. Bioessays 2005, 27, 673–675. [Google Scholar] [CrossRef]
- Pavlopoulos, A.; Akam, M. Hox gene Ultrabithorax regulates distinct sets of target genes at successive stages of Drosophila haltere morphogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 2855–2860. [Google Scholar] [CrossRef] [Green Version]
- Abzhanov, A.; Holtzman, S.; Kaufman, T.C. The Drosophila proboscis is specified by two Hox genes, proboscipedia and Sex combs reduced, via repression of leg and antennal appendage genes. Development 2001, 128, 2803–2814. [Google Scholar] [CrossRef]
- Martin, A.; Serano, J.M.; Jarvis, E.; Bruce, H.S.; Wang, J.; Ray, S.; Barker, C.A.; O’Connell, L.C.; Patel, N.H. CRISPR/Cas9 Mutagenesis Reveals Versatile Roles of Hox Genes in Crustacean Limb Specification and Evolution. Curr. Biol. 2015, 26, 14–26. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Arias, A.; Ingham, P.W.; Scott, M.P.; Akam, M.E. The spatial and temporal deployment of Dfd and Scr transcripts throughout development of Drosophila. Development 1987, 100, 673–683. [Google Scholar] [CrossRef]
- Chen, P.; Tong, X.L.; Li, D.D.; Fu, M.Y.; He, S.Z.; Hu, H.; Xiang, Z.H.; Lu, C.; Dai, F.Y. Antennapedia is involved in the development of thoracic legs and segmentation in the silkworm, Bombyx mori. Heredity 2013, 111, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.P.; Mishra, R.K. Role of Abd-A and Abd-B in development of abdominal epithelia breaks posterior prevalence rule. PLoS Genet. 2014, 10, e1004717. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Hallerman, E.M.; Peng, Y.; Li, Y. Development of Bt Rice and Bt Maize in China and Their Efficacy in Target Pest Control. Int. J. Mol. Sci. 2016, 17, 1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Zeng, B.; Noland Jeffery, E.; Zhou, X. The coming of RNA-based pest controls. J. Plant. Protection. 2015, 42, 673–690. [Google Scholar]
- Enserink, M.; Hines, J.H.; Vignieri, S.N.; Wigginton, N.S. Smarter pest control. The pesticide paradox. Introduction. Science 2013, 341, 728–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Yan, S.C.; Huang, Y.P.; Tan, A.J.; Stanley, D.W.; Song, Q.S. Genetic transformation mediated by piggyBac in the Asian corn borer, Ostrinia furnacalis (Lepidoptera: Crambidae). Arch. Insect Biochem. Physiol. 2012, 80, 140–150. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sgRNA | Conc (ng/μL) | Injected 1 | Hatched 2 | L Mutant 3 | Pupation 3 | P Mutant 4 | Adult 5 | A Mutant 6 |
|---|---|---|---|---|---|---|---|---|
| OfAbd-A | 300 | 785 | 165 (21.0) | 92 (55.8) | 87 (52.7) | 29 (33.3) | - | - |
| 150 | 596 | 197 (33.1) | 75 (38.1) | 128 (65.1) | 18 (24.0) | - | - | |
| Ubx | 300 | 486 | 179 (36.8) | - | 118 (65.9) | 53 (44.9) | 62 (52.5) | 18 (29.3) |
| 150 | 568 | 234 (41.2) | - | 160 (68.4) | 58 (36.2) | 95 (59.4) | 24 (25.3) | |
| GFP | 300 | 245 | 152 (62.0) | - | 108 (71.1) | - | 70 (64.8) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bi, H.; Merchant, A.; Gu, J.; Li, X.; Zhou, X.; Zhang, Q. CRISPR/Cas9-Mediated Mutagenesis of Abdominal-A and Ultrabithorax in the Asian Corn Borer, Ostrinia furnacalis. Insects 2022, 13, 384. https://doi.org/10.3390/insects13040384
Bi H, Merchant A, Gu J, Li X, Zhou X, Zhang Q. CRISPR/Cas9-Mediated Mutagenesis of Abdominal-A and Ultrabithorax in the Asian Corn Borer, Ostrinia furnacalis. Insects. 2022; 13(4):384. https://doi.org/10.3390/insects13040384
Chicago/Turabian StyleBi, Honglun, Austin Merchant, Junwen Gu, Xiaowei Li, Xuguo Zhou, and Qi Zhang. 2022. "CRISPR/Cas9-Mediated Mutagenesis of Abdominal-A and Ultrabithorax in the Asian Corn Borer, Ostrinia furnacalis" Insects 13, no. 4: 384. https://doi.org/10.3390/insects13040384
APA StyleBi, H., Merchant, A., Gu, J., Li, X., Zhou, X., & Zhang, Q. (2022). CRISPR/Cas9-Mediated Mutagenesis of Abdominal-A and Ultrabithorax in the Asian Corn Borer, Ostrinia furnacalis. Insects, 13(4), 384. https://doi.org/10.3390/insects13040384