
The Dynamic Distribution of Wolbachia and Rickettsia in AsiaII1 Bemisia tabaci


 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Host Plant and Whitefly Cultures
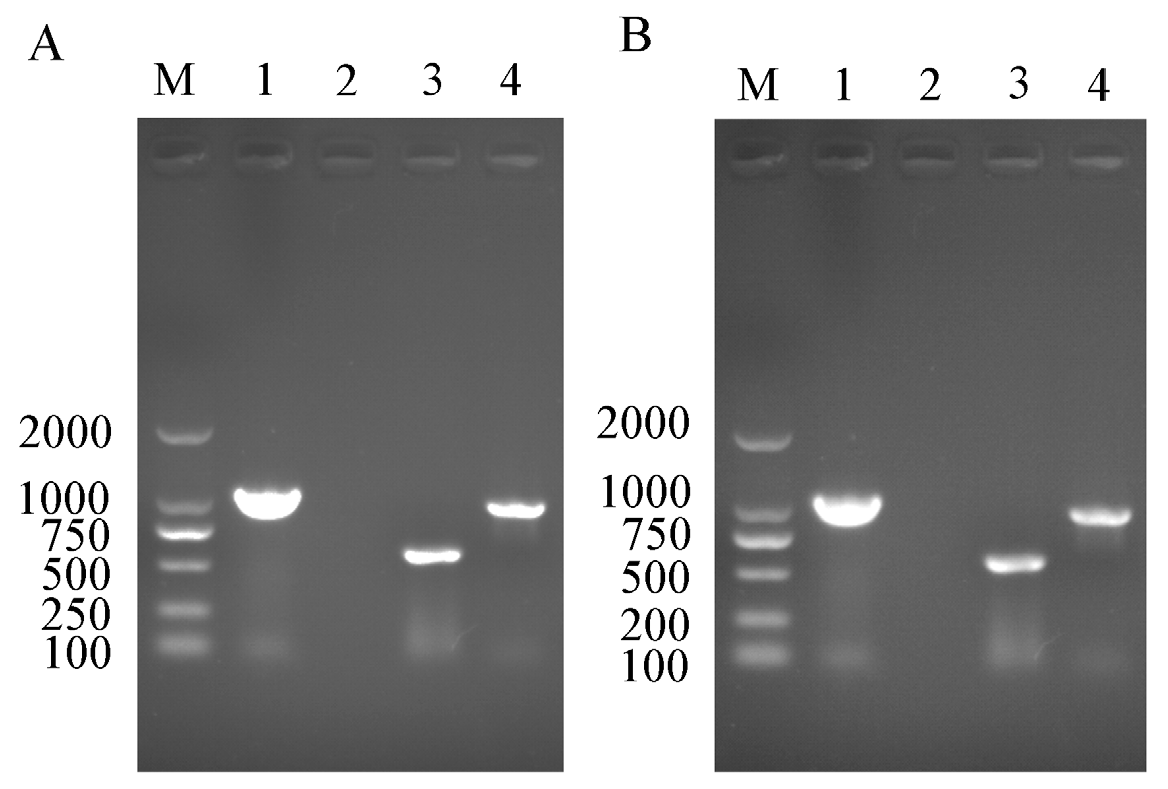
2.2. PCR Detection of Rickettsia and Wolbachia in Asia II1 Whitefly
2.3. Infection Monitoring and Maternal Transmission Efficiency of Rickettsia and Wolbachia in Laboratory Lines
2.4. Quantitative Detection of Rickettsia and Wolbachia
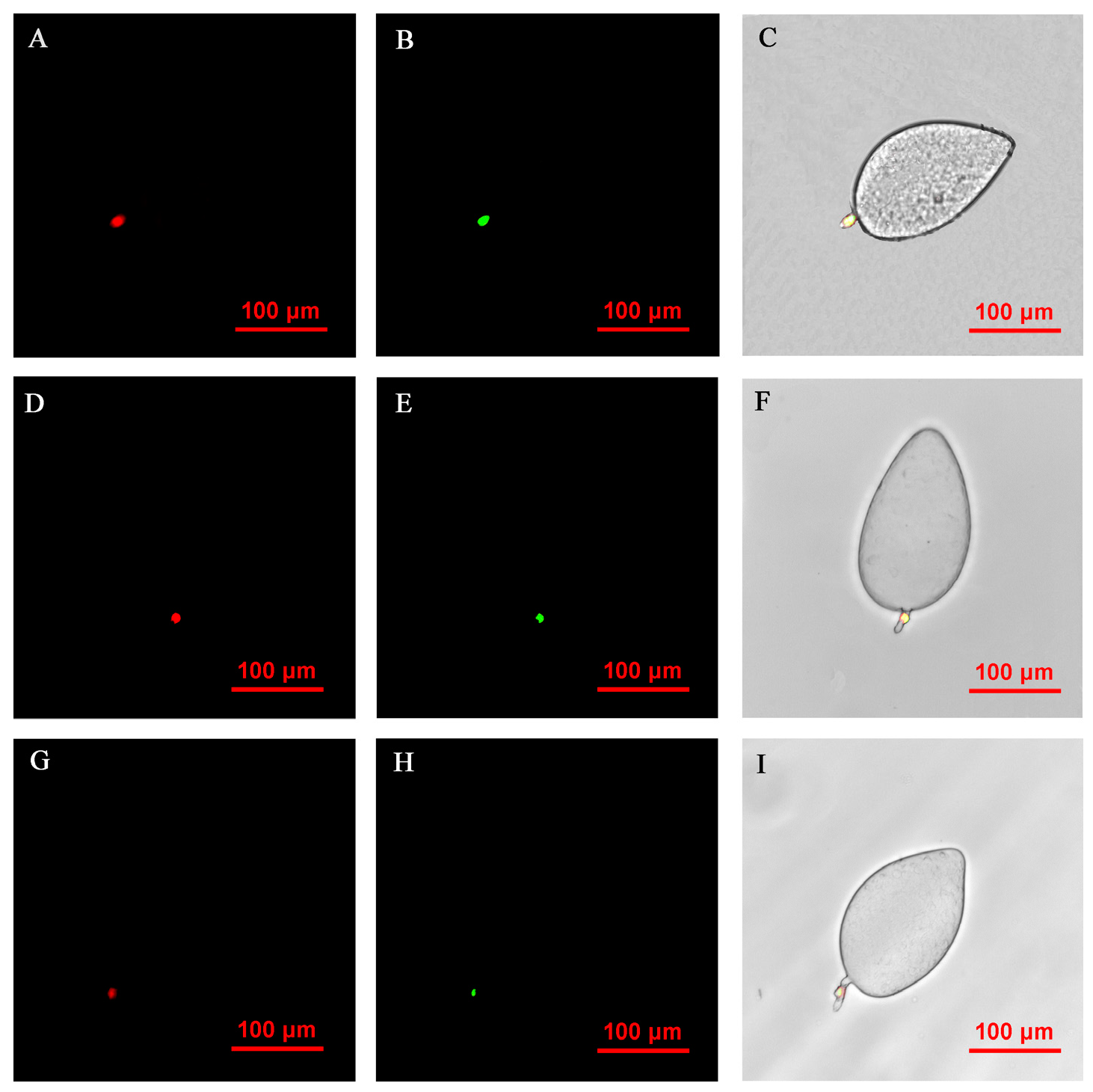
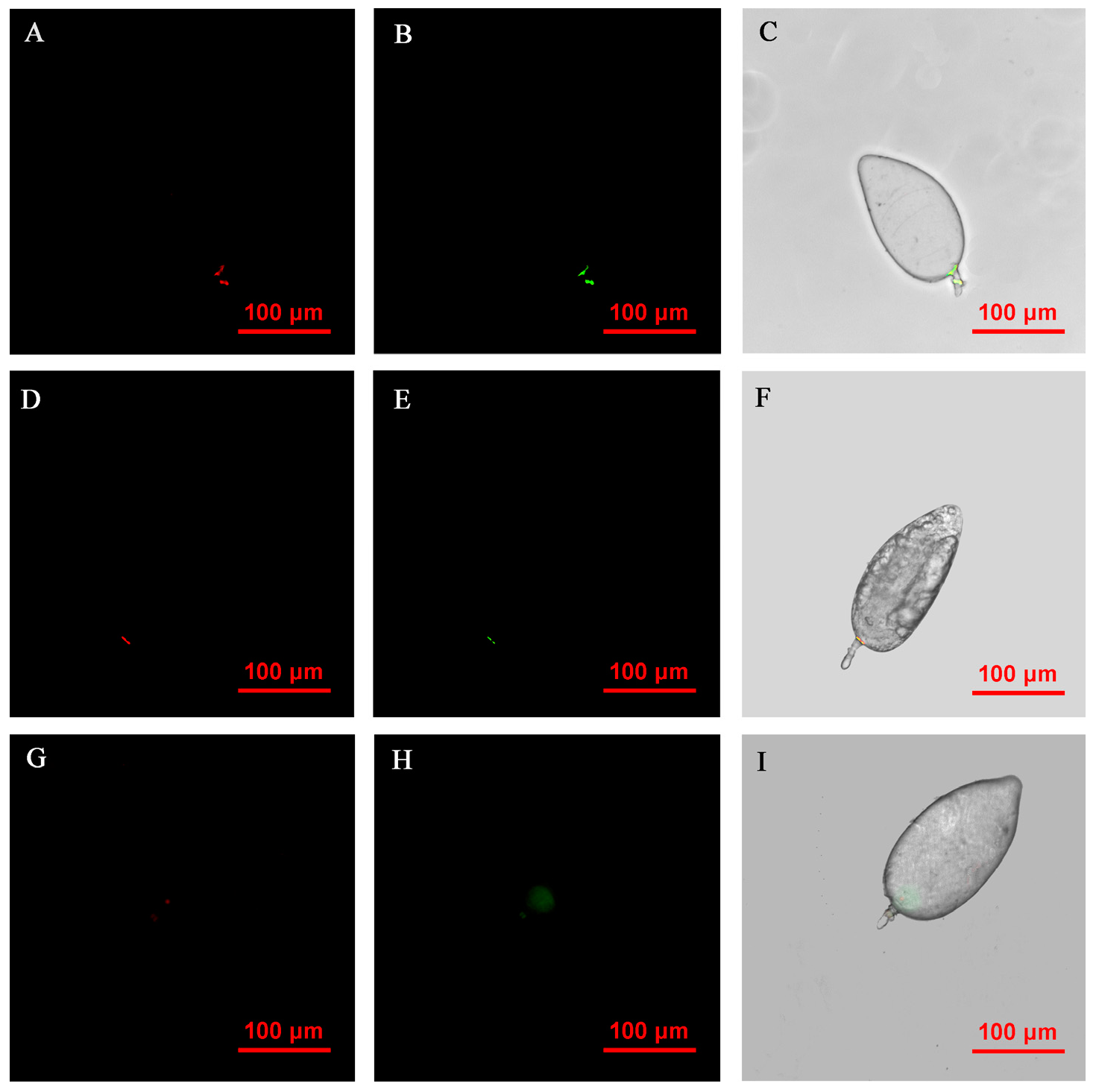
2.5. Fluorescence In Situ Hybridization Microscopy of Rickettsia and Wolbachia in Asia II1 Whiteflies
2.6. Data Analysis
3. Results
3.1. Rickettsia and Wolbachia Detection in Different Asia II1 Whiteflies
3.2. Maternal Transmission Efficiency of Rickettsia and Wolbachia of AsiaII1 IWR Line
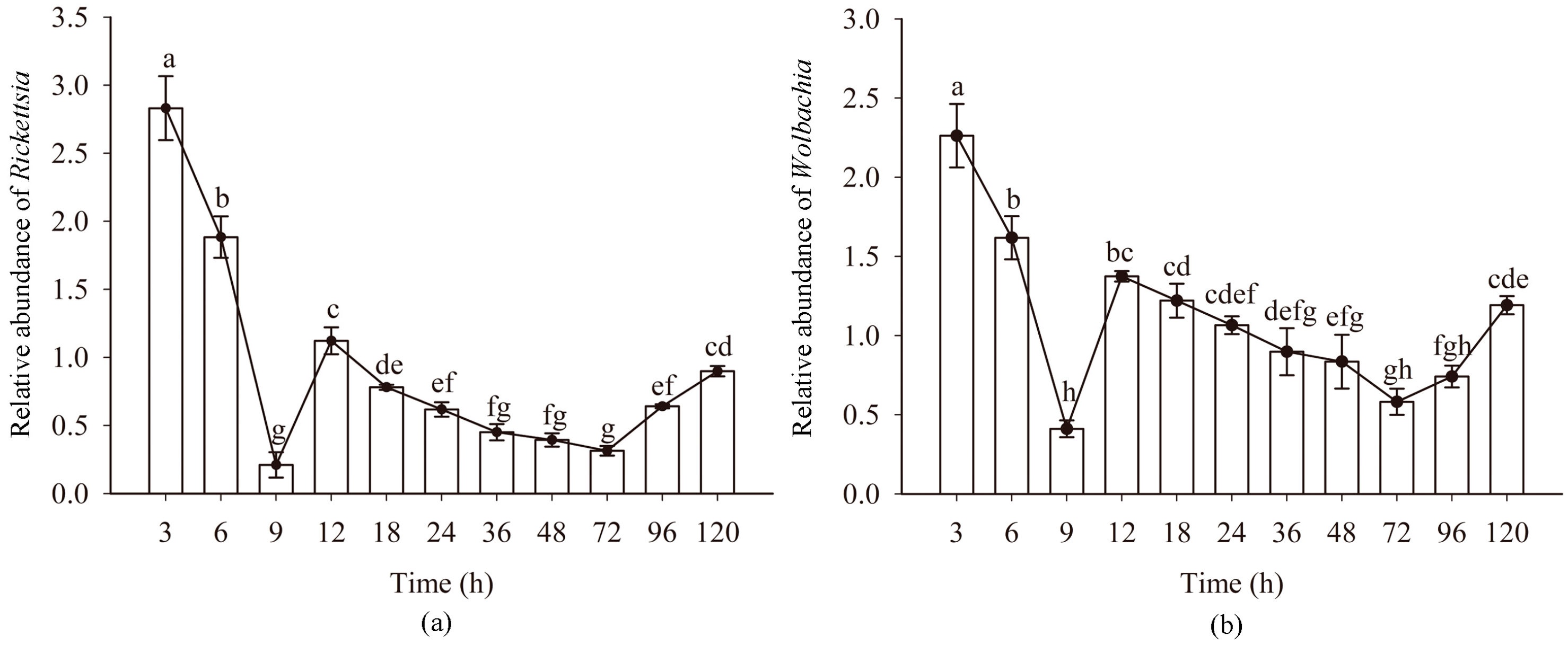
3.3. Quantitative Detection of Rickettsia and Wolbachia in Eggs of Asia II1 Whitefly
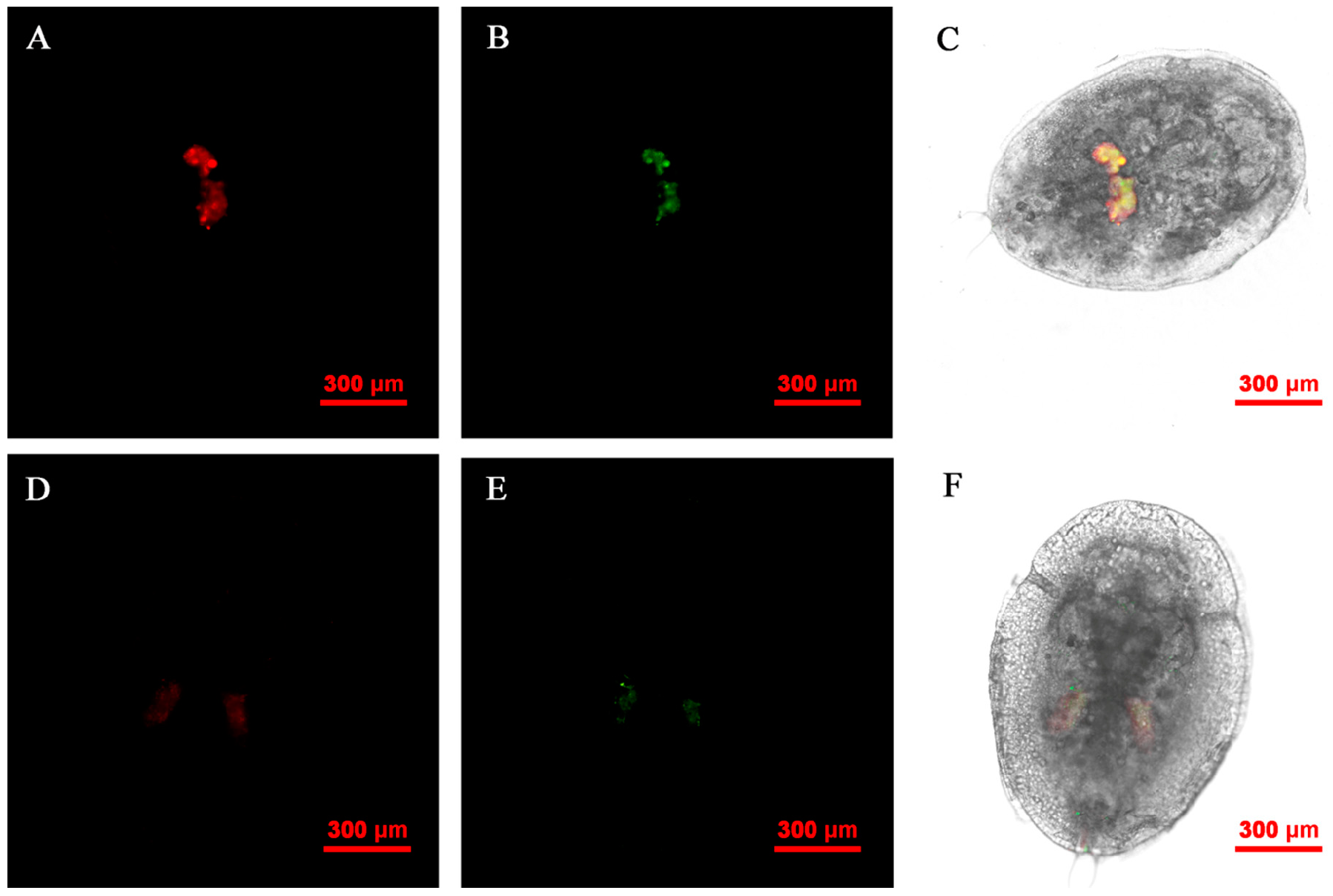
3.4. FISH Visualization of Rickettsia and Wolbachia in Eggs at Different Times
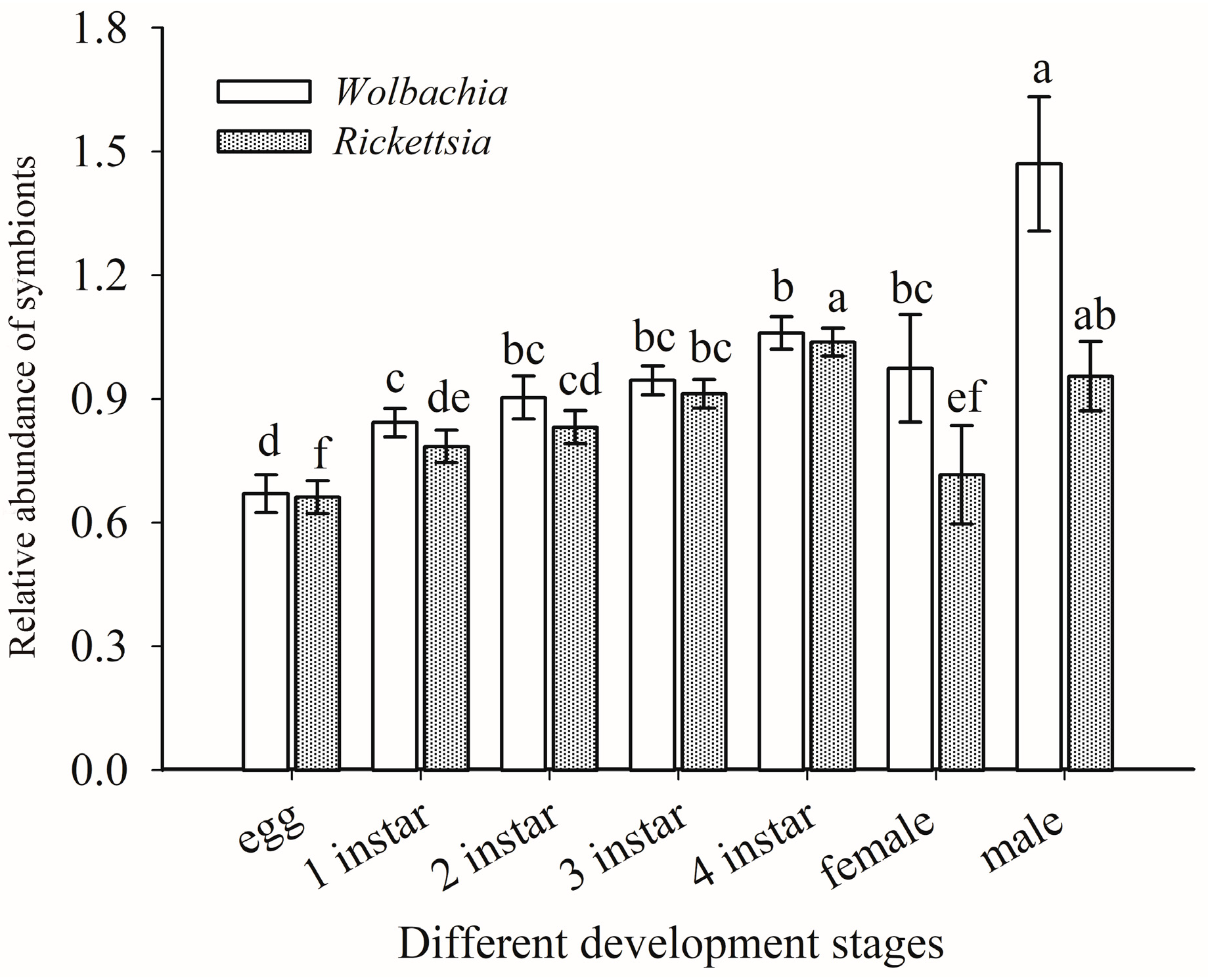
3.5. Quantitative Detection of Rickettsia and Wolbachia in Asia II1 Whiteflies
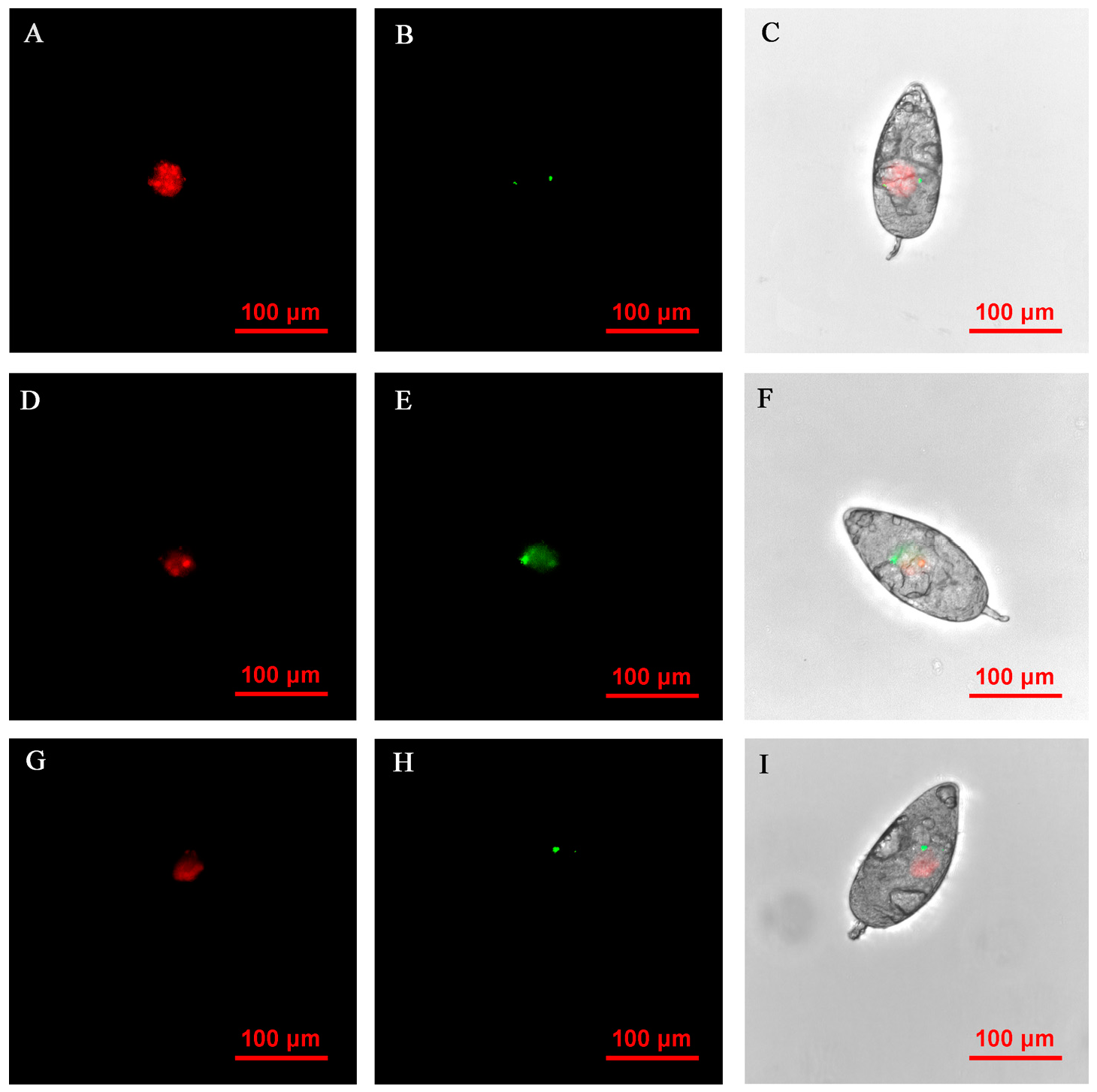
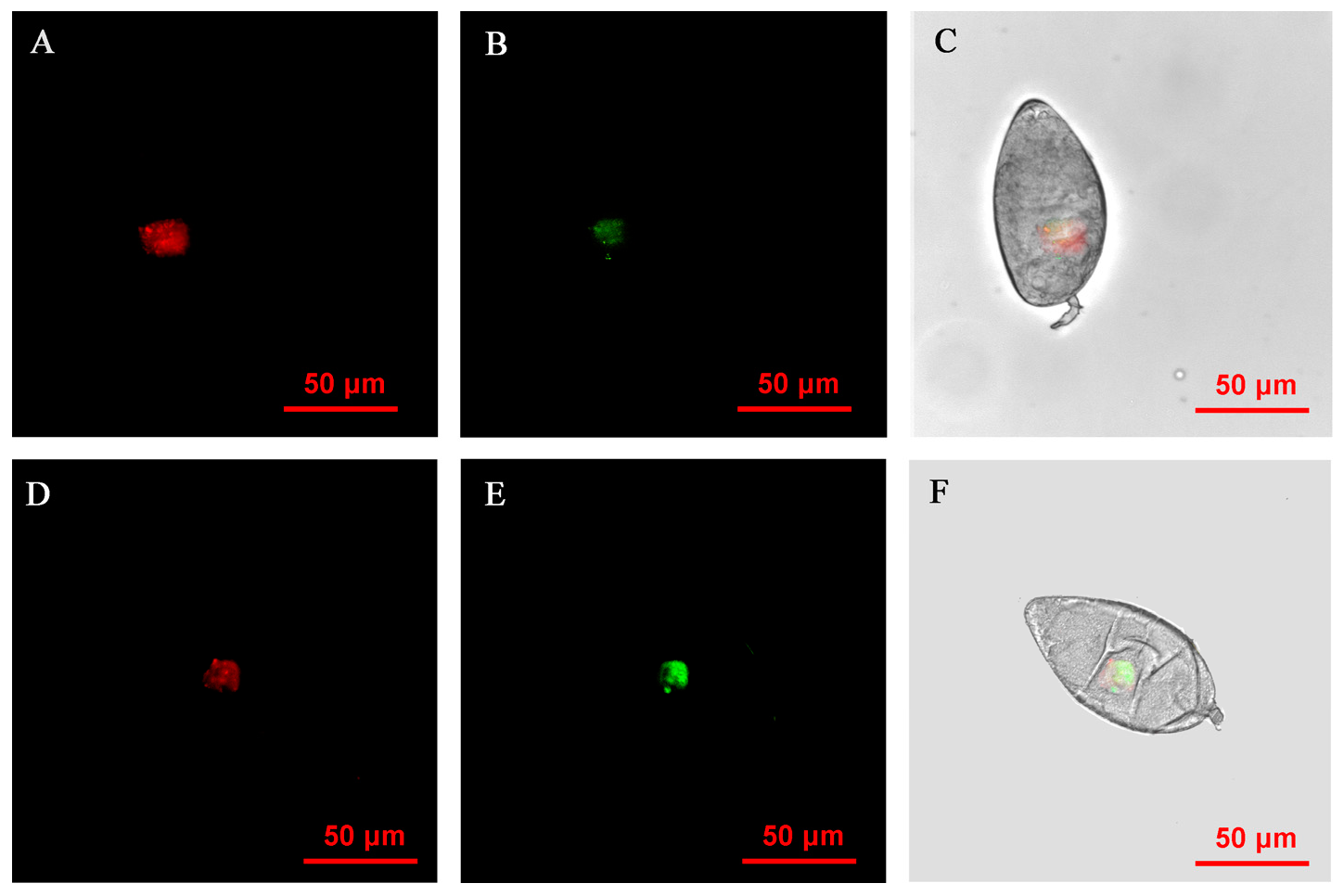
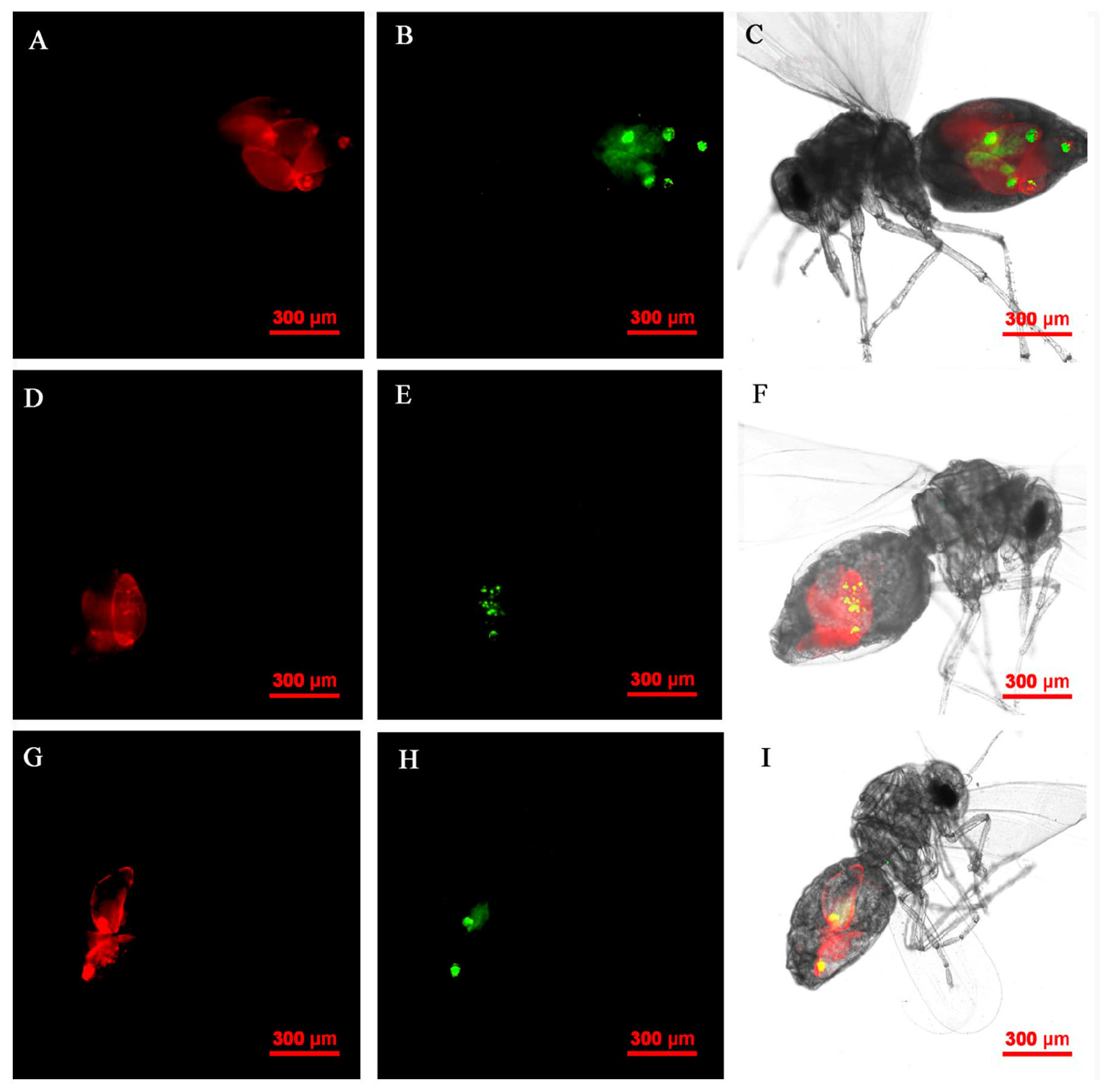
3.6. FISH Visualization of Rickettsia and Wolbachia in Nymphs and Adults
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weinert, L.A.; Araujo-Jnr, E.V.; Ahmed, M.Z.; Welch, J.J. The incidence of bacterial endosymbionts in terrestrial arthropods. Proc. Biol. Sci. 2015, 282, 20150249. [Google Scholar] [CrossRef]
- Wilson, A.C.C.; Duncan, R.P. Signatures of host/symbiont genome coevolution in insect nutritional endosymbioses. Proc. Natl. Acad. Sci. USA 2015, 112, 10255–10261. [Google Scholar] [CrossRef]
- Morrow, J.L.; Hall, A.A.G.; Riegler, M. Symbionts in waiting: The dynamics of incipient endosymbiont complementation and replacement in minimal bacterial communities of psyllids. Microbiome 2017, 5, 58. [Google Scholar] [CrossRef]
- Lv, N.; Lei, W.; Wen, S.; Liu, C.Z.; Qiu, B.L. Effects of endosymbiont disruption on the nutritional dynamics of the pea aphid Acyrthosiphon pisum. Insects 2018, 9, 161. [Google Scholar] [CrossRef]
- Pan, X.L.; Zhou, G.L.; Wu, J.H.; Bian, G.W.; Lu, P.; Raikhel, A.S. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, 13–14. [Google Scholar] [CrossRef] [PubMed]
- Maire, J.; Vincent-Monégat, C.; Masson, F.; Zaidman-Rémy, A.; Heddi, A. An IMD-like pathway mediates both endosymbiont control and host immunity in the cereal weevil Sitophilus spp. Microbiome 2018, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.F.; Bing, X.L.; Zhao, D.S.; Guo, Y.; Xi, Z.Y.; Hoffmann, A.A.; Zhang, K.J.; Huang, H.J.; Gong, J.T.; Zhang, X.; et al. Wolbachia supplement biotin and riboflavin to enhance reproduction in planthoppers. ISME J. 2020, 14, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.; Sun, X.; Fei, Z.; Douglas, A.E. Maternal inheritance of a single somatic animal cell displayed by the bacteriocyte in the whitefly Bemisia tabaci. Curr. Biol. 2018, 28, 459–465. [Google Scholar] [CrossRef]
- Shan, H.W.; Liu, Y.Q.; Luan, J.B.; Liu, S.S. New insight into the transovarial transmission of the symbiont Rickettsia in whitefly. Sci. China Life Sci. 2021, 64, 1174–1186. [Google Scholar] [CrossRef]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Mugerwa, H.; Colvin, J.; Alicai, T.; Omongo, A.C.; Kabaalu, P.; Visendi, P. Genetic diversity of whitefly (Bemisia spp.) on crop and uncultivated plants in Uganda: Implications for the control of this devastating pest species complex in Africa. J. Pest. Sci. 2021, 94, 1307–1330. [Google Scholar] [CrossRef]
- Luan, J.B.; Shan, H.W.; Isermann, P.; Huang, J.H.; Lammerding, J.; Liu, S.S.; Douglas, A.E. Cellular and molecular remodelling of a host cell for vertical transmission of bacterial symbionts. Proc. Biol. Sci. 2016, 283, 20160580. [Google Scholar] [CrossRef] [PubMed]
- Perotti, M.A.; Clarke, H.K.; Turner, B.D.; Braig, H.R. Rickettsia as obligate and mycetomic bacteria. FASEB J. 2006, 20, 2372–2374. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.C.; Sakurai, M.; Bonhomme, J.; Tsuchida, T.; Koga, R.; Fukatsu, T. Elimination of a specialized facultative symbiont does not affect the reproductive mode of its aphid host. Ecol. Entomol. 2007, 32, 296–301. [Google Scholar] [CrossRef]
- Kontsedalov, S.E.; Zchori-Fein, E.; Chiel, Y.; Gottlieb, M.; Inbar, M.; Ghanim, M. The presence of Rickettsia is associated with increased susceptibility of Bemisia tabaci (Homoptera: Aleyrodidae) to insecticides. Pest Manag. Sci. 2008, 64, 789–792. [Google Scholar] [CrossRef]
- Zhou, X.F.; Li, Z.X. Establishment of the cytoplasmic incompatibility-inducing Wolbachia strain wMel in an important agricultural pest insect. Sci. Rep. 2016, 6, 39200. [Google Scholar] [CrossRef]
- Giorgini, M.; Bernardo, U.; Monti, M.M.; Nappo, A.G.; Gebiola, M. Rickettsia symbionts cause parthenogenetic reproduction in the parasitoid wasp pnigalio soemius (Hymenoptera: Eulophidae). Appl. Environ. Microbiol. 2010, 76, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Bourtzis, K. Wolbachia-based technologies for insect pest population control. Adv. Exp. Med. Biol. 2008, 627, 104–113. [Google Scholar]
- Christodoulou, M. Biological vector control of mosquito-borne diseases. Lancet Infect Dis. 2011, 11, 84–85. [Google Scholar] [CrossRef] [PubMed]
- Hancock, P.A.; Sinkins, S.P.; Godfray, H.C. Strategies for introducing Wolbachia to reduce transmission of mosquito-borne diseases. PLoS Negl. Trop Dis. 2011, 5, e1024. [Google Scholar] [CrossRef]
- Hughes, G.L.; Rasgon, J.L. Transinfection: A method to investigate Wolbachia-host interactions and control arthropd-borne diseases. Insect Mol. Biol. 2014, 23, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Strunov, A.; Kiseleva, E.; Gottlieb, Y. Spatial and temporal distribution of pathogenic Wolbachia strain wMelPop in Drosophila melanogaster central nervous system under different temperature conditions. J. Invertebr. Pathol. 2013, 114, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Ahmed, M.; Lv, N.; Shi, P.Q.; Wang, X.M.; Huang, J.L. Plant-mediated horizontal transmission of Wolbachia between whiteflies. ISME J. 2017, 11, 1019–1028. [Google Scholar] [CrossRef]
- Liu, Y.; Fan, Z.Y.; An, X.; Shi, P.Q.; Ahmed, M.Z.; Qiu, B.L. A single-pair method to screen Rickettsia-infected and uninfected whitefly Bemisia tabaci populations. J. Microbiol. Methods 2020, 168, 105795. [Google Scholar] [CrossRef]
- Ahmed, M.Z.; Ren, S.X.; Xue, X.; Li, X.X.; Jin, G.H.; Qiu, B.L. Prevalence of endosymbionts in Bemisia tabaci populations and their in vivo sensitivity to antibiotics. Curr. Microbiol. 2010, 61, 322–328. [Google Scholar] [CrossRef]
- Gottlieb, Y.; Ghanim, M.; Chiel, E.; Gerling, D.; Portnoy, V. Identification and localization of a Rickettsia sp. in Bemisia tabaci (Homoptera: Aleyrodidae). Appl. Environ. Microbiol. 2006, 72, 3646–3652. [Google Scholar] [CrossRef]
- Zhou, W.; Rousset, F.; O’Neill, S. Phylogeny and PCR-based classification of Wolbachia strains using WSP gene sequences. P. Roy. Soc. B-Biol. Sci. 1998, 265, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.; Morrow, J.L.; Spooner-Hart, R.N.; Riegler, M. Independent cytoplasmic incompatibility induced by Cardinium and Wolbachia maintains endosymbiont coinfections in haplodiploid thrips populations. Evolution 2017, 71, 995–1008. [Google Scholar] [CrossRef]
- Ghanim, M.; Kontsedalov, S. Susceptibility to insecticides in the Q biotype of Bemisia tabaci is correlated with bacterial symbiont densities. Pest Manag. Sci. 2009, 65, 939–942. [Google Scholar] [CrossRef]
- Pan, H.P.; Chu, D.; Liu, B.M.; Xie, W.; Wang, S.L.; Wu, Q.J.; Xu, Y.X.; Zhang, Y.J. Relative amount of symbionts in insect hosts changes with host-plant adaptation and insecticide resistance. Environ. Entomol. 2013, 42, 74–78. [Google Scholar] [CrossRef]
- Lv, N.; Peng, J.; Chen, X.Y.; Guo, C.F.; Sang, W.; Wnag, X.M. Antagnostic interaction between male-killing and cytoplasmic incompatibility induced by Cardinium and Wolbachia in the whitefly, Bemisia tabaci. Insect Sci. 2021, 28, 330–346. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, Y.; Ghanim, M.; Gueguen, G.; Kontsedalov, S.; Zchori-Fein, E. Inherited intracellular ecosystem: Symbiotic bacteria share bacteriocytes in whiteflies. FASEB J. 2008, 22, 2591–2599. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Liu, S.S.; Colvin, J.; De Barro, P.J. Species concepts as applied to the whitefly Bemisia tabaci systematics: How many species are there? J. Integr. Agr. 2012, 11, 176–186. [Google Scholar] [CrossRef]
- Tang, X.T.; Cai, L.; Shen, Y.; Du, Y.Z. Diversity and evolution of the endosymbionts of Bemisia tabaci in China. Peer J. 2018, 6, e5516. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.Q.; He, Z.; Li, S.j.; An, X.; Lv, N.; Ghanim, M.; Qiu, B.L. Wolbachia has two different localization patterns in whitefly Bemisia tabaci Asia II 7 species. PLoS ONE 2016, 11, e0162558. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.X.; Song, Y.L.; Zhang, Y.K.; Hoffmann, A.A.; Zhou, J.C.; Sun, J.T. Incidence of facultative bacterial endosymbionts in spider mites associated with local environment and host plant. Appl. Environ. Microb. 2018, 84, e02546-17. [Google Scholar] [CrossRef]
- Shan, H.W.; Lu, Y.H.; Bing, X.L.; Liu, S.S.; Liu, Y.Q. Differential responses of the whitefly Bemisia tabaci symbionts to unfavorable low and high temperatures. Microb. Ecol. 2014, 68, 472–482. [Google Scholar] [CrossRef]
- Ren, S.L.; Li, Y.H.; Ou, D.; Guo, Y.J.; Qiu, B.L. Localization and dynamics of Wolbachia infection in Asian citrus psyllid Diaphorina citri, the insect vector of the causal pathogens of Huanglongbing. Microbiol. Open 2018, 7, e00561. [Google Scholar] [CrossRef] [PubMed]
- Marisa, S.; Katja, Z.; Smiljana Goreta, B.; Svetlana, K.; Murad, G. Co-infection and localization of secondary symbionts in two whitefly species. BMC Microbiol. 2010, 10, 142. [Google Scholar]
- Zchori-Fein, E.; Brown, J.K. Diversity of prokaryotes associated with Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2002, 6, 711–718. [Google Scholar] [CrossRef]
- Chiel, E.; Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Katzir, N.; Inbar, M.; Ghanim, M. Biotype-dependent secondary symbiont communities in sympatric populations of Bemisia tabaci. B Entomol. Res. 2007, 97, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Weeks, A.R.; Velten, R.; Stouthamer, R. Incidence of a new sex-ratio-distorting endosymbiotic bacterium among arthropods. Proc. R. Soc. Lond. B. 2003, 270, 1857–1865. [Google Scholar] [CrossRef] [PubMed]
- Thao, M.L.; Baumann, P. Evolutionary relationships of primary prokaryotic endosymbionts of whiteflies and their hosts. Appl. Environ. Microb. 2004, 70, 3401–3406. [Google Scholar] [CrossRef] [PubMed]
- Everett, K.D.E.; Thao, M.L.; Horn, M.; Dyszynski, G.E.; Baumann, P. Novel chlamydiae in whiteflies and scale insects: Endosymbionts ‘Candidatus Fritschea bemisiae’ strain Falk and ‘Candidatus Fritschea eriococci’ strain Elm. Int. J. Syst. Evol. Micr. 2005, 55, 1581–1587. [Google Scholar] [CrossRef] [PubMed]
- Bing, X.L.; Yang, J.; Zchori-Fein, E.; Wang, X.W.; Liu, S.S. Characterization of a newly discovered symbiont of the whitefly bemisia tabaci (Hemiptera: Aleyrodidae). Appl. Environ. Microb. 2012, 79, 569–575. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Whitefly Samples | n Females | n Offspring Per Female | Total N Tested | Rickettsia | Wolbachia | ||||
|---|---|---|---|---|---|---|---|---|---|
| n+ | n− | % | n+ | n− | % | ||||
| ♀IWR | 10 | 10 | 100 | 100 | 0 | 100 | 100 | 0 | 100 |
| ♀IWR × IWR♂ | 10 | 10 | 100 | 100 | 0 | 100 | 100 | 0 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, N.; Peng, J.; He, Z.-Q.; Wen, Q.; Su, Z.-Q.; Ali, S.; Liu, C.-Z.; Qiu, B.-L. The Dynamic Distribution of Wolbachia and Rickettsia in AsiaII1 Bemisia tabaci. Insects 2023, 14, 401. https://doi.org/10.3390/insects14040401
Lv N, Peng J, He Z-Q, Wen Q, Su Z-Q, Ali S, Liu C-Z, Qiu B-L. The Dynamic Distribution of Wolbachia and Rickettsia in AsiaII1 Bemisia tabaci. Insects. 2023; 14(4):401. https://doi.org/10.3390/insects14040401
Chicago/Turabian StyleLv, Ning, Jing Peng, Zi-Qi He, Qin Wen, Zheng-Qin Su, Shaukat Ali, Chang-Zhong Liu, and Bao-Li Qiu. 2023. "The Dynamic Distribution of Wolbachia and Rickettsia in AsiaII1 Bemisia tabaci" Insects 14, no. 4: 401. https://doi.org/10.3390/insects14040401
APA StyleLv, N., Peng, J., He, Z. -Q., Wen, Q., Su, Z. -Q., Ali, S., Liu, C. -Z., & Qiu, B. -L. (2023). The Dynamic Distribution of Wolbachia and Rickettsia in AsiaII1 Bemisia tabaci. Insects, 14(4), 401. https://doi.org/10.3390/insects14040401