
Evaluating the Sublethal Effects of Origanum vulgare Essential Oil and Carvacrol on the Biological Characteristics of Culex pipiens biotype molestus (Diptera: Culicidae)


 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals Tested
2.2. Mosquito Colony
2.3. Dose–response Bioassays (LC50 Determination)
2.4. Sublethal Effects after Exposure to Larvicidal LC50 Concentrations
2.4.1. Survival and Longevity of Larvae and Pupae
2.4.2. Adult Longevity, Fecundity, Fertility, and Wing Length
2.4.3. Data Analysis
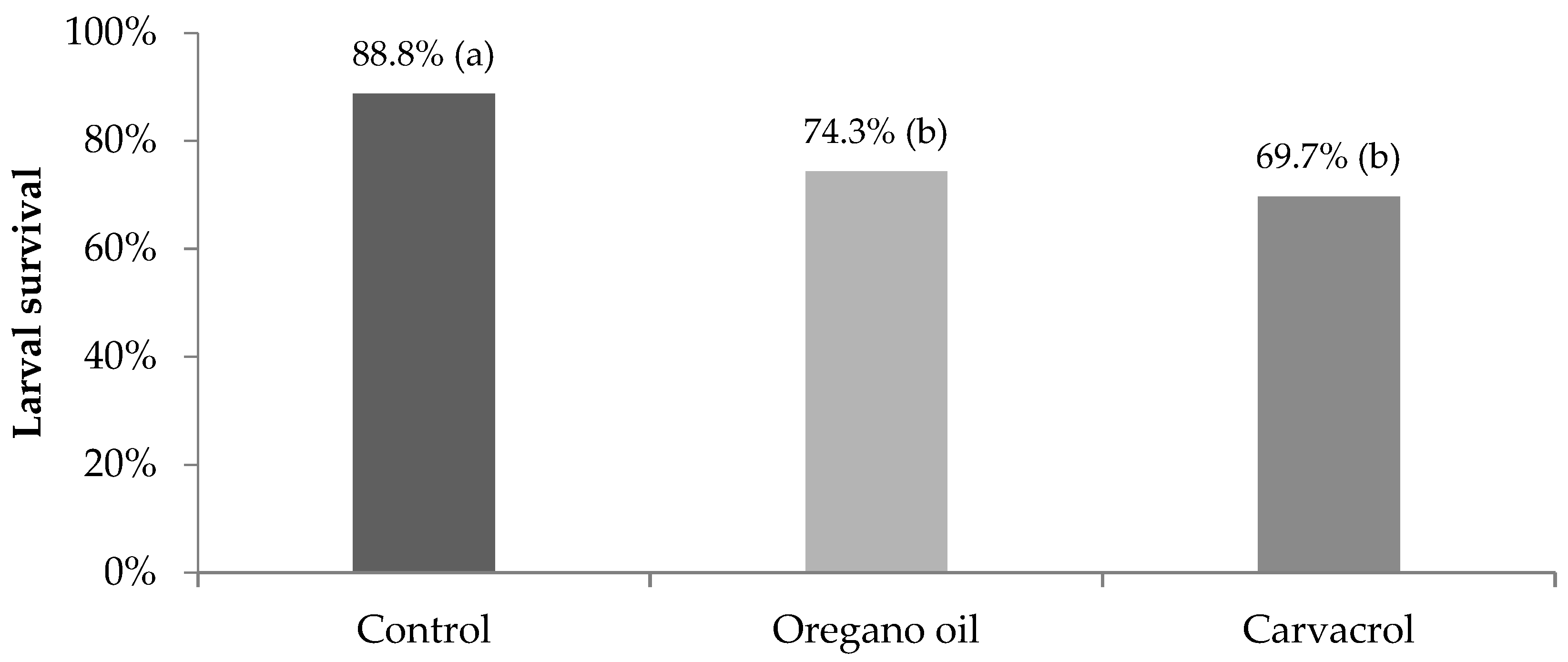
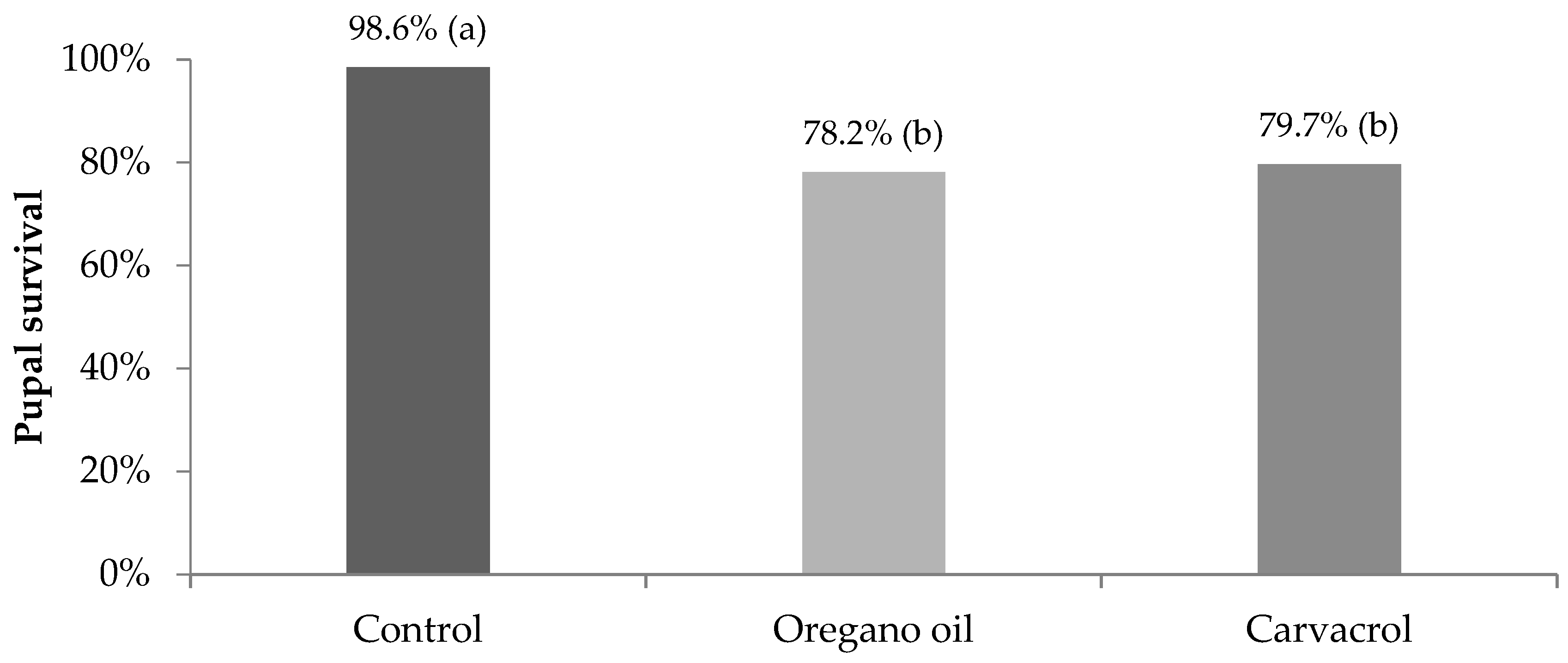
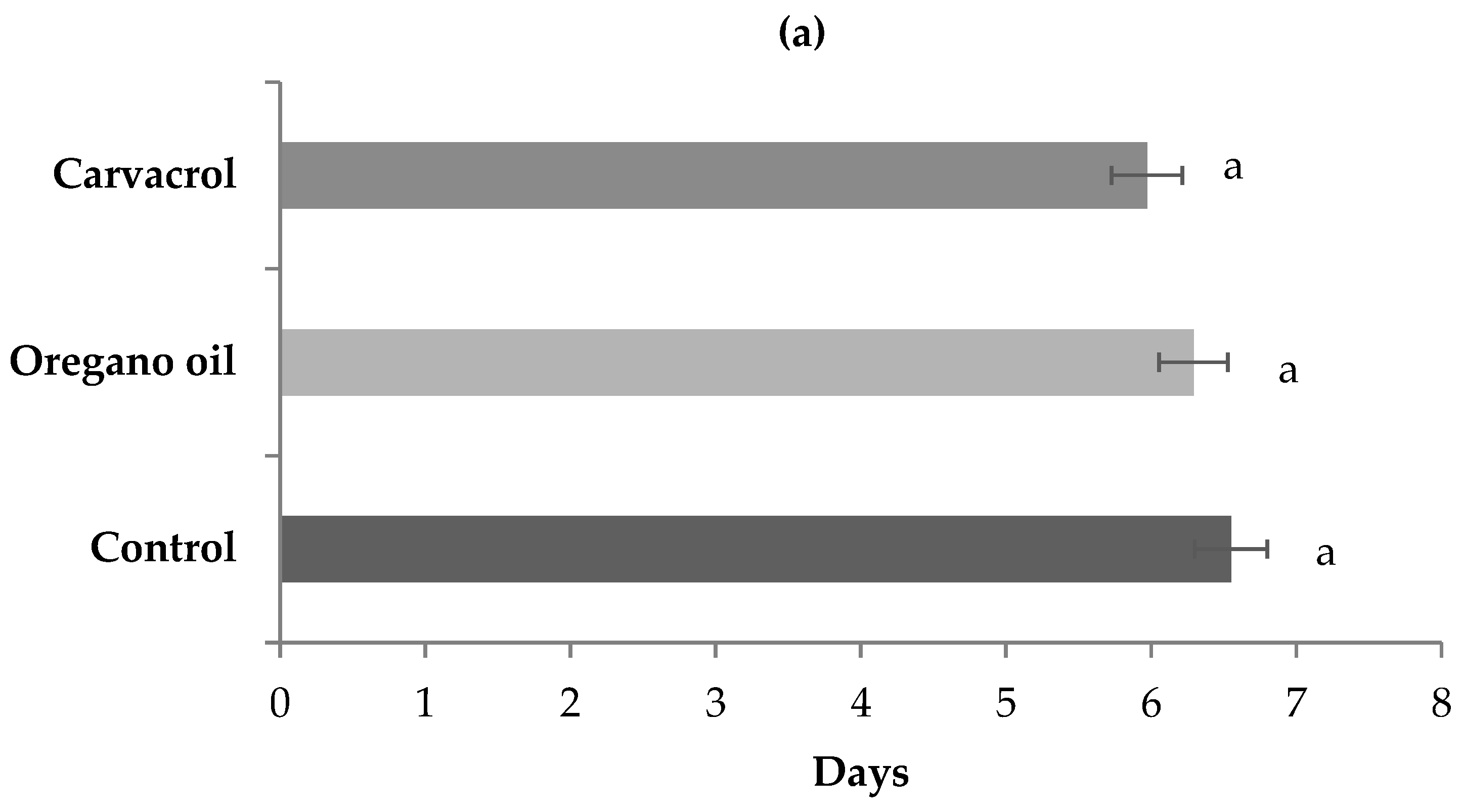
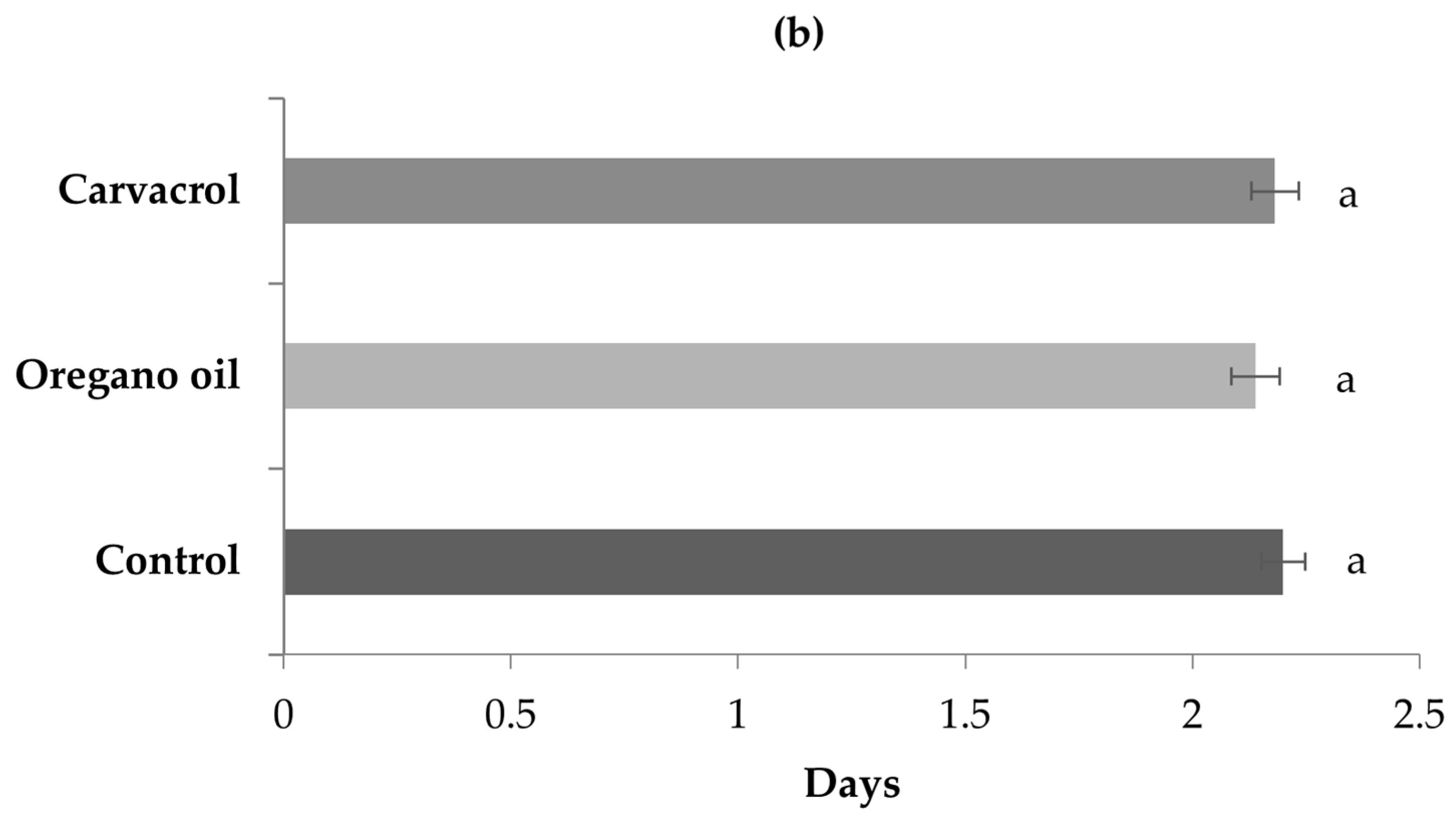
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2010; p. 577. [Google Scholar]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. BioMed Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Bolzoni, L.; Chadwick, E.A.; Capelli, G.; Montarsi, F.; Grisenti, M.; de la Puente, J.M.; Muñoz, J.; Figuerola, J.; Soriguer, R.; et al. Understanding West Nile virus ecology in Europe: Culex pipiens host feeding preference in a hotspot of virus emergence. Parasit Vectors 2015, 8, 213. [Google Scholar] [CrossRef] [PubMed]
- García-Carrasco, J.M.; Muñoz, A.R.; Olivero, J.; Segura, M.; Real, R. Predicting the spatio-temporal spread of West Nile virus in Europe. PLoS Negl. Trop. Dis. 2021, 15, e0009022. [Google Scholar] [CrossRef] [PubMed]
- Tsioka, K.; Gewehr, S.; Pappa, S.; Kalaitzopoulou, S.; Stoikou, K.; Mourelatos, S.; Papa, A. West Nile Virus in Culex mosquitoes in central Macedonia, Greece, 2022. Viruses 2023, 15, 224. [Google Scholar] [CrossRef] [PubMed]
- Bellini, R.; Zeller, H.; Van Bortel, W. A review of the vector management methods to prevent and control outbreaks of West Nile virus infection and the challenge for Europe. Parasit. Vectors 2014, 7, 323. [Google Scholar] [CrossRef]
- Osório, H.C.; Zé-Zé, L.; Amaro, F.; Nunes, A.; Alves, M.J. Sympatric occurrence of Culex pipiens (Diptera, Culicidae) biotypes pipiens, molestus and their hybrids in Portugal, Western Europe: Feeding patterns and habitat determinants. Med. Vet. Entomol. 2014, 28, 103–109. [Google Scholar] [CrossRef]
- Vogels, C.B.; Fros, J.J.; Göertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of northern European Culex pipiens biotypes and hybrids for West Nile virus is differentially affected by temperature. Parasit. Vectors. 2016, 9, 393. [Google Scholar] [CrossRef]
- Martínez-de la Puente, J.; Ferraguti, M.; Ruiz, S.; Roiz, D.; Soriguer, R.C.; Figuerola, J. Culex pipiens forms and urbanization: Effects on blood feeding sources and transmission of avian Plasmodium. Malar. J. 2016, 15, 589. [Google Scholar] [CrossRef]
- Brugman, V.A.; Hernández-Triana, L.M.; Medlock, J.M.; Fooks, A.R.; Carpenter, S.; Johnson, N. The role of Culex pipiens L. (Diptera: Culicidae) in virus transmission in Europe. Int. J. Environ. Res. Public Health 2018, 15, 389. [Google Scholar] [CrossRef]
- European Chemicals Agency (ECHA). Available online: https://echa.europa.eu/el/information-on-chemicals/biocidal-products (accessed on 10 January 2023).
- Nauen, R. Insecticide resistance in disease vectors of public health importance. Pest Manag. Sci. 2007, 63, 628–633. [Google Scholar] [CrossRef]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Grigoraki, L.; Puggioli, A.; Mavridis, K.; Douris, V.; Montanari, M.; Bellini, R.; Vontas, J. Striking diflubenzuron resistance in Culex pipiens, the prime vector of West Nile Virus. Sci. Rep. 2017, 7, 11699. [Google Scholar] [CrossRef]
- Porretta, D.; Fotakis, E.A.; Mastrantonio, V.; Chaskopoulou, A.; Michaelakis, A.; Kioulos, I.; Weill, M.; Urbanelli, S.; Vontas, J.; Bellini, R. Focal distribution of diflubenzuron resistance mutations in Culex pipiens mosquitoes from Northern Italy. Acta Trop. 2019, 193, 106–112. [Google Scholar] [CrossRef]
- Said-Al Ahl, H.A.H.; Hikal, W.M.; Tkachenko, K.G. Essential oils with potential as insecticidal agents: A review. Int. J. Environ. Plan. Manag. 2017, 3, 23–33. [Google Scholar]
- Boulogne, I.; Petit, P.; Ozier-Lafontaine, H.; Desfontaines, L.; Loranger-Merciris, G. Insecticidal and antifungal chemicals produced by plants: A review. Environ. Chem. Lett. 2012, 10, 325–347. [Google Scholar] [CrossRef]
- Vargas, M.V. An update on published literature (period 1992–2010) and botanical categories on plant essential oils with effects on mosquitoes (Diptera: Culicidae). Bol. Malariol. Sal. Amb. 2012, 52, 143–193. [Google Scholar]
- Piplani, M.; Bhagwat, D.P.; Singhvi, G.; Sankaranarayanan, M.; Balana-Fouce, R.; Vats, T.; Chander, S. Plant-based larvicidal agents: An overview from 2000 to 2018. Exp. Parasitol. 2019, 199, 92–103. [Google Scholar] [CrossRef]
- Sengül Demirak, M.S.; Canpolat, E. Plant-based bioinsecticides for mosquito control: Impact on insecticide resistance and disease transmission. Insects 2022, 13, 162. [Google Scholar] [CrossRef]
- Conti, B.; Canale, A.; Bertoli, A.; Gozzini, F.; Pistelli, L. Essential oil composition and larvicidal activity of six Mediterranean aromatic plants against the mosquito Aedes albopictus (Diptera: Culicidae). Parasitol. Res. 2010, 107, 1455–1462. [Google Scholar] [CrossRef]
- Govindarajan, M.; Sivakumar, R.; Rajeswary, M.; Yogalakshmi, K. Chemical composition and larvicidal activity of essential oil from Ocimum basilicum (L.) against Culex tritaeniorhynchus, Aedes albopictus and Anopheles subpictus (Diptera: Culicidae). Exp. Parasitol. 2013, 134, 7–11. [Google Scholar] [CrossRef]
- Pavela, R. Essential oils for the development of eco-friendly mosquito larvicides: A review. Ind. Crop. Prod. 2015, 76, 174–187. [Google Scholar] [CrossRef]
- Ntalli, N.; Koliopoulos, G.; Giatropoulos, A.; Menkissoglu-Spiroudi, U. Plant secondary metabolites against arthropods of medical importance. Phytochem. Rev. 2019, 18, 1255–1275. [Google Scholar] [CrossRef]
- Senthil-Nathan, S. A review of resistance mechanisms of synthetic insecticides and botanicals, Phytochemicals, and essential oils as alternative larvicidal agents against mosquitoes. Front Physiol. 2020, 10, 1591. [Google Scholar] [CrossRef] [PubMed]
- Rattan, R.S. Mechanism of action of insecticidal secondary metabolites of plant origin. Crop Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef]
- Traboulsi, A.F.; Taoubi, K.; El-Haj, S.; Bessiere, J.M.; Rammal, S. Insecticidal properties of essential plant oils against the mosquito Culex pipiens molestus (Diptera: Culicidae). Pest Manag. Sci. 2002, 58, 491–495. [Google Scholar] [CrossRef]
- Cetin, H.; Yanikoglu, A. A study of the larvicidal activity of Origanum (Labiatae) species from south west Turkey. J. Vector Ecol. 2006, 31, 118–122. [Google Scholar] [CrossRef]
- Michaelakis, A.; Theotokatos, S.A.; Koliopoulos, G.; Chorianopoulos, N.G. Essential oils of Satureja species: Insecticidal effect on Culex pipiens larvae (Diptera: Culicidae). Molecules 2007, 12, 2567–2578. [Google Scholar] [CrossRef]
- Pavela, R. Larvicidal property of essential oils against Culex quinquefasciatus Say (Diptera: Culicidae). Ind. Crop. Prod. 2009, 30, 311–315. [Google Scholar] [CrossRef]
- Govindarajan, M.; Rajeswary, M.; Hoti, S.L.; Benelli, G. Larvicidal potential of carvacrol and terpinen-4-ol from the essential oil of Origanum vulgare (Lamiaceae) against Anopheles stephensi, Anopheles subpictus, Culex quinquefasciatus and Culex tritaeniorhynchus (Diptera: Culicidae). Res. Vet. Sci. 2016, 104, 77–82. [Google Scholar] [CrossRef]
- Bouguerra, N.; Tine-Djebbar, F.; Soltani, N. Oregano essential oil as potential mosquito larvicides. Transylv. Rev. 2019, 39, 9612–9619. [Google Scholar]
- Youssefi, M.R.; Tabari, M.A.; Esfandiari, A.; Kazemi, S.; Moghadamnia, A.A.; Sut, S.; Dall’Acqua, S.; Benelli, G.; Maggi, F. Efficacy of two monoterpenoids, carvacrol and thymol, and their combinations against eggs and larvae of the West Nile vector Culex pipiens. Molecules 2019, 24, 1867. [Google Scholar] [CrossRef]
- Enan, E.E. Insecticidal activity of essential oils: Octopaminergic sites of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2001, 130, 325–337. [Google Scholar] [CrossRef]
- Jukic, M.; Politeo, O.; Maksimovic, M.; Milos, M.; Milos, M. In vitro acetylcholinesterase inhibitory properties of thymol, carvacrol and their derivatives thymoquinone and thymohydroquinone. Phytother. Res. 2007, 21, 259–261. [Google Scholar] [CrossRef]
- Tong, F.; Coats, J.R. Effects of monoterpenoid insecticides on [3H]-TBOB binding in house fly GABA receptor and 36 Cl- uptake in American cockroach ventral nerve cord. Pestic. Biochem. Physiol. 2010, 98, 317–324. [Google Scholar] [CrossRef]
- Anderson, J.A.; Coats, J.R. Acetylcholinesterase inhibition by nootkatone and carvacrol in arthropods. Pestic. Biochem. Physiol. 2012, 102, 124–128. [Google Scholar] [CrossRef]
- Evergetis, E.; Bellini, R.; Balatsos, G.; Michaelakis, A.; Carrieri, M.; Veronesi, R.; Papachristos, D.P.; Puggioli, A.; Kapsaski-Kanelli, V.-N.; Haroutounian, S.A. From bio-prospecting to field assessment: The case of carvacrol rich essential oil as a potent mosquito larvicidal and repellent agent. Front. Ecol. Evol. 2018, 6, 204. [Google Scholar] [CrossRef]
- Giatropoulos, A.; Bellini, R.; Pavlopoulos, D.T.; Balatsos, G.; Karras, V.; Mourafetis, F.; Papachristos, D.P.; Karamaouna, F.; Carrieri, M.; Veronesi, R.; et al. Efficacy evaluation of oregano essential oil mixed with Bacillus thuringiensis israelensis and diflubenzuron against Culex pipiens and Aedes albopictus in road drains of Italy. Insects 2022, 13, 977. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential oils as ecofriendly biopesticides? Challenges and constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef]
- Pavela, R. Lethal and sublethal effects of thyme oil (Thymus vulgaris L.) on the house fly (Musca domestica Lin.). J. Essent. Oil Bear. Plants 2007, 10, 346–356. [Google Scholar] [CrossRef]
- Pavela, R.; Vrchotova, N.; Triska, J. Mosquitocidal activities of thyme oils (Thymus vulgaris L.) against Culex quinquefasciatus (Diptera: Culicidae). Parasitol. Res. 2009, 105, 1365–1370. [Google Scholar] [CrossRef] [PubMed]
- Giatropoulos, A.; Karamaouna, F.; Ampatzi, A.; Papachristos, D.; Michaelakis, A. Sublethal effects of oregano essential oil and its major compound carvacrol on biological parameters of Aedes albopictus (Diptera: Culicidae). Exp. Parasitol. 2022, 242, 108392. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Guidelines for Laboratory and Field Testing of Mosquito Larvicides World Health Organization (WHO) Communicable Disease Control, Prevention and Eradication WHO Pesticide Evaluation Scheme (WHOPES); World Health Organization (WHO): Geneva, Switzerland, 2005; pp. 1–41. [Google Scholar]
- van-den-Huevel, M.J. The effect of rearing temperature on the wing length, thorax length, leg length and ovariole number of the adult mosquito, Aedes aegypti (L.). Trans. R. Entomol. Soc. Lond. 1963, 115, 197–216. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: London, UK, 1971. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry, 3rd ed.; W. H. Freeman: New York, NY, USA, 1995. [Google Scholar]
- Ndung’u, M.; Torto, B.; Knols, B.G.J.; Hassanali, A. Laboratory evaluation of some eastern African Meliaceae as sources of larvicidal botanicals for Anopheles gambiae. Int. J. Trop. Insect Sci. 2004, 24, 311–318. [Google Scholar] [CrossRef]
- Shaalan, E.A.; Canyon, D.; Younes, M.W.; Abdel-Wahab, H.; Mansour, A.H. A review of botanical phytochemicals with mosquitocidal potential. Environ. Int. 2005, 31, 1149–1166. [Google Scholar] [CrossRef]
- Muema, J.M.; Bargul, J.L.; Njeru, S.N.; Onyango, J.O.; Imbahale, S.S. Prospects for malaria control through manipulation of mosquito larval habitats and olfactory-mediated behavioural responses using plant-derived compounds. Parasit. Vectors 2017, 10, 184. [Google Scholar] [CrossRef]
- Abdelgaleil, S.A.M.; El-Sabrout, A.M. Composition, toxicity and developmental potential of three essential oils on the West Nile virus mosquito, Culex pipiens L. Int. J. Pest Manag. 2020, 69, 175–183. [Google Scholar] [CrossRef]
- Giatropoulos, A.; Papachristos, D.P.; Kimbaris, A.; Koliopoulos, G.; Polissiou, M.G.; Emmanouel, N.; Michaelakis, A. Evaluation of bioefficacy of three citrus essential oils against the dengue vector Aedes albopictus (Diptera: Culicidae) in correlation to their components enantiomeric distribution. Parasitol. Res. 2012, 111, 2253–2263. [Google Scholar] [CrossRef]
- Zahran, H.E.D.M.; Abdelgaleil, S.A.M. Insecticidal and developmental inhibitory properties of monoterpenes on Cx. pipiens L. (Diptera: Culicidae). J. Asia Pacific Entomol. 2011, 14, 46–51. [Google Scholar] [CrossRef]
- Dakhil, M.A.; Morsy, T.A. The larvicidal activities of the peel oils of three citrus fruits against Cx. pipiens. J. Egypt. Soc. Parasitol. 1999, 29, 347–352. [Google Scholar]
- Pavela, R.; Maggi, F.; Mazzara, E.; Torresi, J.; Cianfaglione, K.; Benelli, G.; Canale, A. Prolonged sublethal effects of essential oils from non-wood parts of nine conifers on key insect pests and vectors. Ind. Crop. Prod. 2021, 168, 113590. [Google Scholar] [CrossRef]
- Ochola, J.B.; Mutero, C.M.; Marubu, R.M.; Haller, B.F.; Hassanali, A.; Lwande, W. Mosquitoes larvicidal activity of Ocimum kilimandscharicum oil formulation under laboratory and field-simulated conditions. Insects 2022, 13, 203. [Google Scholar] [CrossRef]
- Kamiabi, F.; Jaal, Z.; Keng, C.L. Bioefficacy of crude extract of Cyperus aromaticus (Family: Cyperaceae) cultured cells, against Aedes aegypti and Aedes albopictus mosquitoes. Asian Pac. J. Trop. Biomed. 2013, 3, 767–775. [Google Scholar] [CrossRef]
- Zuharah, W.F.; Ahbirami, R.; Dieng, H.; Thiagaletchumi, M.; Fadzly, N. Evaluation of sublethal effects of Ipomoea cairica Linn. extract on life history traits of Dengue vectors. Rev. Inst. Med. Trop. Sao Paulo 2016, 58, 44. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Giordan, C.; Casettarie, L.; Curzi, G.; Cappellacci, L.; Petrelli, R.; Maggi, F. Acute and sub-lethal toxicity of eight essential oils of commercial interest against the filariasis mosquito Culex quinquefasciatus and the housefly Musca domestica. Ind. Crop. Prod. 2018, 112, 668–680. [Google Scholar] [CrossRef]
- Muthukrishnan, J.; Pushpalatha, E. Effects of plant extracts on fecundity and fertility of mosquitoes. J. Appl. Ent. 2001, 125, 31–35. [Google Scholar] [CrossRef]
- Conti, B.; Flamini, G.; Cioni, P.L.; Ceccarini, L.; Leonardi, M.M.; Benelli, G. Mosquitocidal essential oils: Are they safe against non-target aquaticorganisms? Parasitol. Res. 2014, 113, 251–259. [Google Scholar] [CrossRef]
- Arslan, Ö.C.; Parlak, H.; Boyacıoglu, M.; Karaaslan, M.A. Acute toxicty of several essential oils on Daphnia magna (Straus, 1816). Ege J. Fish. Aquat. Sci. 2014, 31, 137–143. [Google Scholar]
- Pavela, R. Insecticidal properties of Pimpinella anisum essential oils against the Culex quinquefasciatus and the non-target organism Daphnia magna. J. Asia-Pacif. Entomol. 2014, 17, 287–293. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Material | Slope (±SEM) | LC50 (95% CL) a | LC90 (95% CL) a | x2 | d.f. |
|---|---|---|---|---|---|
| Oregano oil | 5.11 ± 0.43 | 19.86 (16.97–23.48) | 35.36 (28.60–53.81) | 106.539 b | 22 |
| Carvacrol | 2.77 ± 0.30 | 6.41 (5.78–7.02) | 18.57 (15.47–24.31) | 21.165 | 26 |
| Parameter | Control | Oregano Oil | Carvacrol | p Values |
|---|---|---|---|---|
| Sex ratio (males:females) | 1.69:1 (70) | 1.53:1 (43) | 1.75:1 (55) | 0.799 |
| Male longevity (days) | 25.8 ± 2.52 a (26) | 23.6 ± 3.0 ab (16) | 17.4 ± 2.53 b (19) | 0.041 |
| Female longevity (days) | 32.9 ± 2.9 (26) | 34.8 ± 2.5 (16) | 29.8 ± 3.0 (19) | 0.397 |
| Pre-oviposition period (days) | 5.7 ± 0.7 (23) | 5.2 ± 0.5 (13) | 4.9 ± 0.6 (15) | 0.634 |
| Fecundity (eggs per female) | 45.4 ± 3.8 (26) | 42.6 ± 6.6 (16) | 32.3 ± 4.9 (19) | 0.101 |
| Fertility (larvae per female) | 42.4 ± 4.4 (26) | 37.8 ± 6.2 (16) | 28.9 ± 5.1 (19) | 0.092 |
| Fertile females (%) | 84.6 (26) | 81.3 (16) | 68.4 (19) | 0.690 |
| Wing length of males (mm) | 2.67 ± 0.02 (23) | 2.69 ± 0.013 (16) | 2.68 ± 0.03 (17) | 0.986 |
| Wing length of females (mm) | 3.20 ± 0.019 (25) | 3.17 ± 0.022 (14) | 3.18 ± 0.017 (19) | 0.378 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giatropoulos, A.; Koliopoulos, G.; Pantelakis, P.-N.; Papachristos, D.; Michaelakis, A. Evaluating the Sublethal Effects of Origanum vulgare Essential Oil and Carvacrol on the Biological Characteristics of Culex pipiens biotype molestus (Diptera: Culicidae). Insects 2023, 14, 400. https://doi.org/10.3390/insects14040400
Giatropoulos A, Koliopoulos G, Pantelakis P-N, Papachristos D, Michaelakis A. Evaluating the Sublethal Effects of Origanum vulgare Essential Oil and Carvacrol on the Biological Characteristics of Culex pipiens biotype molestus (Diptera: Culicidae). Insects. 2023; 14(4):400. https://doi.org/10.3390/insects14040400
Chicago/Turabian StyleGiatropoulos, Athanasios, George Koliopoulos, Pavlos-Nektarios Pantelakis, Dimitrios Papachristos, and Antonios Michaelakis. 2023. "Evaluating the Sublethal Effects of Origanum vulgare Essential Oil and Carvacrol on the Biological Characteristics of Culex pipiens biotype molestus (Diptera: Culicidae)" Insects 14, no. 4: 400. https://doi.org/10.3390/insects14040400
APA StyleGiatropoulos, A., Koliopoulos, G., Pantelakis, P. -N., Papachristos, D., & Michaelakis, A. (2023). Evaluating the Sublethal Effects of Origanum vulgare Essential Oil and Carvacrol on the Biological Characteristics of Culex pipiens biotype molestus (Diptera: Culicidae). Insects, 14(4), 400. https://doi.org/10.3390/insects14040400