

Provisioning Australian Seed Carrot Agroecosystems with Non-Floral Habitat Provides Oviposition Sites for Crop-Pollinating Diptera

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
- Will eristaline syrphid flies use provisioned habitat pools as oviposition sites in a commercial field setting otherwise unsuitable for larval development?
- Which of the two habitats resulted in the greatest number of eggs and larvae?
- What were the specific features within the habitat pools that resulted in the greatest oviposition?
2. Materials and Methods
2.1. Study Sites
2.2. Deploying the Habitat Pools
2.3. Surveying the Immature Life Stages of Eristaline Flies
2.4. Rearing Eristaline Flies from Pools
2.5. Statistical Analyses
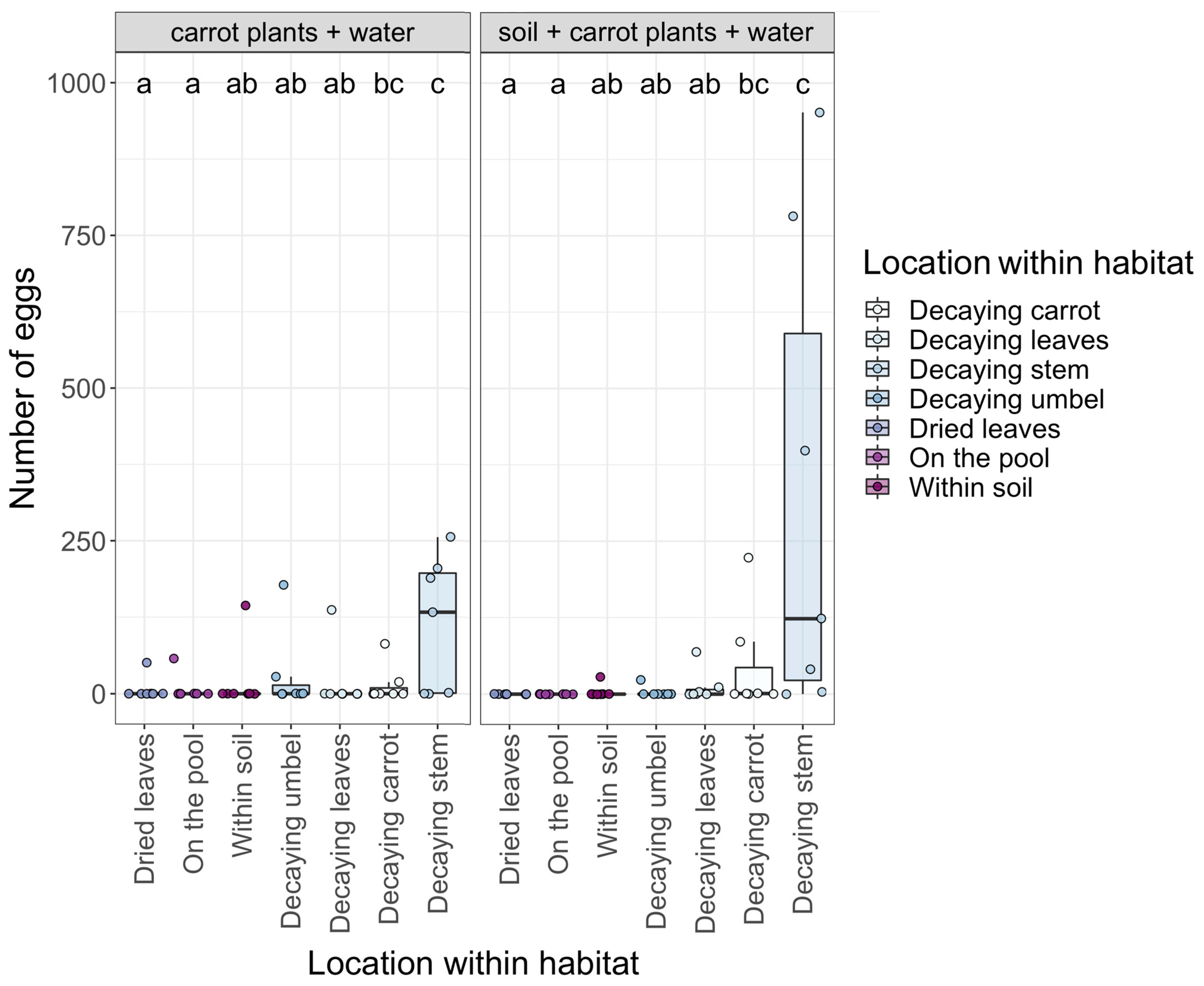
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fijen, T.P.M.; Read, S.F.J.; Walker, M.K.; Gee, M.; Nelson, W.R.; Howlett, B.G. Different landscape features within a simplified agroecosystem support diverse pollinators and their service to crop plants. Landsc. Ecol. 2022, 37, 1787–1799. [Google Scholar] [CrossRef]
- Finch, J.; Gilpin, A.-M.; Cook, J. Fishing for flies: Testing the efficacy of “stink stations” for promoting blow flies as pollinators in mango orchards. J. Pollinat. Ecol. 2023, 32, 79–100. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Seymour, C.L.; Nicolson, S.W.; Veldtman, R. Creating patches of native flowers facilitates crop pollination in large agricultural fields: Mango as a case study. J. Appl. Ecol. 2012, 49, 1373–1383. [Google Scholar] [CrossRef]
- Knapp, M.; Rezac, M. Even the smallest non-crop habitat islands could be beneficial: Distribution of carabid beetles and spiders in agricultural landscape. PLoS ONE 2015, 10, e0123052. [Google Scholar] [CrossRef] [PubMed]
- Altieri, M.A. The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 19–31. [Google Scholar] [CrossRef]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar] [CrossRef]
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; Vandermeer, J.; Whitbread, A. Global food security, biodiversity conservation and the future of agricultural intensification. Biol. Conserv. 2012, 151, 53–59. [Google Scholar] [CrossRef]
- Sunderland, K.; Samu, F. Effects of agricultural diversification on the abundance, distribution, and pest control potential of spiders: A review. Entomol. Exp. Appl. 2000, 95, 1–13. [Google Scholar] [CrossRef]
- Wilson, J.D.; Morris, A.J.; Arroyo, B.E.; Clark, S.C.; Bradbury, R.B. A review of the abundance and diversity of invertebrate and plant foods of granivorous birds in northern Europe in relation to agricultural change. Agric. Ecosyst. Environ. 1999, 75, 13–30. [Google Scholar] [CrossRef]
- Rischen, T.; Frenzel, T.; Fischer, K. Biodiversity in agricultural landscapes: Different non-crop habitats increase diversity of ground-dwelling beetles (Coleoptera) but support different communities. Biodivers. Conserv. 2021, 30, 3965–3981. [Google Scholar] [CrossRef]
- González, E.; Štrobl, M.; Janšta, P.; Hovorka, T.; Kadlec, T.; Knapp, M. Artificial temporary non-crop habitats support parasitoids on arable land. Biol. Conserv. 2022, 265, 109409. [Google Scholar] [CrossRef]
- Landis, D.A. Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 2017, 18, 1–12. [Google Scholar] [CrossRef]
- Schellhorn, N.A.; Bianchi, F.; Hsu, C.L. Movement of Entomophagous Arthropods in Agricultural Landscapes: Links to Pest Suppression. Annu. Rev. Entomol. 2014, 59, 559–581. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.E.; Peisley, R.K.; Rader, R.; Luck, G.W. Pollinators, pests, and predators: Recognizing ecological trade-offs in agroecosystems. Ambio 2016, 45, 4–14. [Google Scholar] [CrossRef]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- Scheper, J.; Bommarco, R.; Holzschuh, A.; Potts, S.G.; Riedinger, V.; Roberts, S.P.M.; Rundlof, M.; Smith, H.G.; Steffan-Dewenter, I.; Wickens, J.B.; et al. Local and landscape-level floral resources explain effects of wildflower strips on wild bees across four European countries. J. Appl. Ecol. 2015, 52, 1165–1175. [Google Scholar] [CrossRef]
- Jonsson, A.M.; Ekroos, J.; Danhardt, J.; Andersson, G.K.S.; Olsson, O.; Smith, H.G. Sown flower strips in southern Sweden increase abundances of wild bees and hoverflies in the wider landscape. Biol. Conserv. 2015, 184, 51–58. [Google Scholar] [CrossRef]
- Haenke, S.; Scheid, B.; Schaefer, M.; Tscharntke, T.; Thies, C. Increasing syrphid fly diversity and density in sown flower strips within simple vs. complex landscapes. J. Appl. Ecol. 2009, 46, 1106–1114. [Google Scholar] [CrossRef]
- Gillespie, M.; Wratten, S.; Sedcole, R.; Colfer, R. Manipulating floral resources dispersion for hoverflies (Diptera: Syrphidae) in a California lettuce agro-ecosystem. Biol. Control 2011, 59, 215–220. [Google Scholar] [CrossRef]
- Munoz, A.E.; Plantegenest, M.; Amouroux, P.; Zaviezo, T. Native flower strips increase visitation by non-bee insects to avocado flowers and promote yield. Basic Appl. Ecol. 2021, 56, 369–378. [Google Scholar] [CrossRef]
- Requier, F.; Leonhardt, S.D. Beyond flowers: Including non-floral resources in bee conservation schemes. J. Insect Conserv. 2020, 24, 5–16. [Google Scholar] [CrossRef]
- Davis, A.E.; Bickel, D.; Saunders, M.E.; Rader, R. Crop-pollinating Diptera have diverse diet and habitat needs in both larval and adult stages. Ecol. Appl. 2023, e2859. [Google Scholar] [CrossRef]
- Campoy, A.; Pérez-Bañón, C.; Aznar, D.; Rojo, S. Description of the preimaginal stages of the golden native dronefly from Australia, Eristalinus punctulatus (Macquart, 1847) (Diptera: Syrphidae). Austral Entomol. 2020, 59, 784–793. [Google Scholar] [CrossRef]
- Davis, A.E.; Deutsch, K.R.; Torres, A.M.; Loya, M.J.M.; Cody, L.V.; Harte, E.; Sossa, D.; Muniz, P.A.; Ng, W.H.; McArt, S.H. Eristalis flower flies can be mechanical vectors of the common trypanosome bee parasite, Crithidia bombi. Sci. Rep. 2021, 11, 15852. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Marshall, S.; Locke, M.; Moran, K.; Crins, W.; Skevington, J. Field Guide to the Flower Flies of Northeastern North America; Princeton University Press: Princeton, NJ, USA, 2019. [Google Scholar]
- Jauker, F.; Bondarenko, B.; Becker, H.C.; Steffan-Dewenter, I. Pollination efficiency of wild bees and hoverflies provided to oilseed rape. Agric. For. Entomol. 2012, 14, 81–87. [Google Scholar] [CrossRef]
- Jarlan, A.; De Oliveira, D.; Gingras, J. Effects of Eristalis tenax (Diptera: Syrphidae) pollination on characteristics of greenhouse sweet pepper fruits. J. Econ. Entomol. 1997, 90, 1650–1654. [Google Scholar] [CrossRef]
- Cook, D.; Voss, S.; Finch, J.; Rader, R.; Cook, J.; Spurr, C. The role of flies as pollinators of horticultural crops: An Australian case study with worldwide relevance. Insects 2020, 11, 341. [Google Scholar] [CrossRef]
- Howlett, B.G.; Gee, M. The potential management of the drone fly (Eristalis tenax) as a crop pollinator in New Zealand. N. Z. Plant Prot. 2019, 72, 221–230. [Google Scholar] [CrossRef]
- Gaffney, A.; Allen, G.R.; Brown, P.H. Insect visitation to flowering hybrid carrot seed crops. N. Z. J. Crop Hortic. Sci. 2011, 39, 79–93. [Google Scholar] [CrossRef]
- Gaffney, A.; Bohman, B.; Quarrell, S.R.; Brown, P.H.; Allen, G.R. Frequent insect visitors are not always pollen carriers in hybrid carrot pollination. Insects 2018, 9, 61. [Google Scholar] [CrossRef]
- Broussard, M.A.; Mas, F.; Howlett, B.; Pattemore, D.; Tylianakis, J.M. Possible mechanisms of pollination failure in hybrid carrot seed and implications for industry in a changing climate. PLoS ONE 2017, 12, e0180215. [Google Scholar] [CrossRef] [PubMed]
- Perez-Banon, C.; Petanidou, T.; Marcos-Garcia, M.A. Pollination in small islands by occasional visitors: The case of Daucus carota subsp commutatus (Apiaceae) in the Columbretes archipelago, Spain. Plant Ecol. 2007, 192, 133–151. [Google Scholar] [CrossRef]
- Reilly, J.; Artz, D.; Biddinger, D.; Bobiwash, K.; Boyle, N.; Brittain, C.; Brokaw, J.; Campbell, J.; Daniels, J.; Elle, E.; et al. Crop production in the USA is frequently limited by a lack of pollinators. Proc. R. Soc. B-Biol. Sci. 2020, 287, 20200922. [Google Scholar] [CrossRef] [PubMed]
- Dolley, W.L., Jr.; Hassett, C.C.; Bowen, W.J. Laboratory culture of the drone fly, Eristalis tenax. Science 1933, 78, 313–314. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, S.; Thyselius, M.; Holden, M.; Nordstrom, K. Rearing and long-Term maintenance of Eristalis tenax hoverflies for research studies. J. Vis. Exp. 2018, 135, 57711. [Google Scholar] [CrossRef]
- Campoy, A.; Sáez, L.; Pérez-Bañón, C.; Rojo, S. Demography and population parameters of two species of eristaline flower flies (Diptera, Syrphidae, Eristalini). J. Appl. Entomol. 2020, 144, 133–143. [Google Scholar] [CrossRef]
- Campoy, A.; Aracil, A.; Pérez-Bañón, C.; Rojo, S. An in-depth study of the larval head skeleton and the external feeding structures related with the ingestion of food particles by the eristaline flower flies Eristalis tenax and Eristalinus aeneus. Entomol. Exp. Appl. 2020, 168, 783–798. [Google Scholar] [CrossRef]
- Campoy, A.; Pérez-Bañón, C.; Rojo, S. Intra-puparial development in the hoverflies Eristalinus aeneus and Eristalis tenax (Diptera: Syrphidae). J. Morphol. 2020, 281, 1436–1445. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- O’Hara, R.B.; Kotze, D.J. Do not log-transform count data. Methods Ecol. Evol. 2010, 1, 118–122. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. 2022. Available online: https://cran.r-project.org/web/packages/DHARMa/vignettes/DHARMa.html (accessed on 10 October 2022).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Rader, R.; Bartomeus, I.; Garibaldi, L.; Garratt, M.; Howlett, B.; Winfree, R.; Cunningham, S.; Mayfield, M.; Arthur, A.; Andersson, G.; et al. Non-bee insects are important contributors to global crop pollination. Proc. Natl. Acad. Sci. USA 2016, 113, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.; Belliure, B.; Montserrat, M.; Gil, J.; Velásquez, Y. Pollination by the hoverfly Eristalinus aeneus (Diptera: Syrphidae) in two hybrid seed crops: Celery and fennel (Apiaceae). J. Agric. Sci. 2022, 160, 1–13. [Google Scholar] [CrossRef]
- Levot, G.W.; Brown, K.R.; Shipp, E. Larval growth of some calliphorid and sarcophagid Diptera. Bull. Entomol. Res. 1979, 69, 469–475. [Google Scholar] [CrossRef]
- Carey, J.R.; Liedo, P.; Muller, H.G.; Wang, J.L.; Chiou, J.M. Relationship of age patterns of fecundity to mortality, longevity, and lifetime reproduction in a large cohort of Mediterranean fruit fly females. J. Gerontol. Ser. A-Biol. Sci. Med. Sci. 1998, 53, B245–B251. [Google Scholar] [CrossRef]
- Vasconcelos, S.D.; Cruz, T.M.; Salgado, R.L.; Thyssen, P.J. Dipterans associated with a decomposing animal carcass in a rainforest fragment in Brazil: Notes on the early arrival and colonization by necrophagous species. J. Insect Sci. 2013, 13, 145. [Google Scholar] [CrossRef]
- Cox, M.E.; Johnstone, R.; Robinson, J. Relationships between perceived coastal waterway condition and social aspects of quality of life. Ecol. Soc. 2006, 11, 35. Available online: https://www.jstor.org/stable/26267792 (accessed on 15 October 2022). [CrossRef]
- Greene, G.L. Rearing techniques for Creophilus maxillosus (Coleoptera: Staphylinidae), a predator of fly larvae in cattle feedlots. J. Econ. Entomol. 1996, 89, 848–851. [Google Scholar] [CrossRef]
- Bornemissza, G.F. An analysis of arthropod succession in carrion and the effect of its decomposition on the soil fauna. Aust. J. Zool. 1957, 5, 1–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | Site | Days | Clutches | Eggs | Larvae | Species |
|---|---|---|---|---|---|---|
| 1 | Site 1 | 21 | 16 | 910 | 0 | E. tenax |
| 1 | Site 2 | 14 | 7 | 494 | 26 | E. tenax |
| 1 | Site 3 | 13 | 6 | 296 | 41 | E. tenax |
| 1 | Site 4 | 12 | 9 | 694 | 117 | E. tenax, E. punctulatus |
| 1 | Site 5 | 12 | 22 | 1355 | 107 | E. tenax |
| 1 | Site 6 | 19 | 9 | 382 | 9 | E. tenax |
| 1 | Site 7 | 19 | 4 | 113 | 3 | E. tenax |
| 2 | Site 1 | 21 | 9 | 476 | 0 | E. tenax |
| 2 | Site 2 | 14 | 2 | 233 | 16 | E. tenax |
| 2 | Site 3 | 13 | 0 | 0 | 41 | E. tenax |
| 2 | Site 4 | 12 | 4 | 258 | 201 | E. tenax, E. punctulatus |
| 2 | Site 5 | 12 | 21 | 1497 | 137 | E. tenax, E. punctulatus |
| 2 | Site 6 | 19 | 6 | 548 | 4 | E. tenax |
| 2 | Site 7 | 19 | 8 | 401 | 0 | E. tenax |
| Habitat | Site | 1st Instar | 2nd Instar | 3rd Instar | Dead |
|---|---|---|---|---|---|
| 1 | Site 1 | 0 | 0 | 0 | 0 |
| 1 | Site 2 | 17 | 4 | 3 | 2 |
| 1 | Site 3 | 35 | 1 | 0 | 5 |
| 1 | Site 4 | 54 | 50 | 12 | 1 |
| 1 | Site 5 | 100 | 0 | 0 | 7 |
| 1 | Site 6 | 1 | 4 | 4 | 0 |
| 1 | Site 7 | 3 | 0 | 0 | 0 |
| 2 | Site 1 | 0 | 0 | 0 | 0 |
| 2 | Site 2 | 2 | 4 | 4 | 6 |
| 2 | Site 3 | 5 | 19 | 13 | 4 |
| 2 | Site 4 | 79 | 90 | 32 | 0 |
| 2 | Site 5 | 135 | 2 | 0 | 0 |
| 2 | Site 6 | 0 | 0 | 0 | 4 |
| 2 | Site 7 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davis, A.E.; Schmidt, L.A.; Harrington, S.; Spurr, C.; Rader, R. Provisioning Australian Seed Carrot Agroecosystems with Non-Floral Habitat Provides Oviposition Sites for Crop-Pollinating Diptera. Insects 2023, 14, 439. https://doi.org/10.3390/insects14050439
Davis AE, Schmidt LA, Harrington S, Spurr C, Rader R. Provisioning Australian Seed Carrot Agroecosystems with Non-Floral Habitat Provides Oviposition Sites for Crop-Pollinating Diptera. Insects. 2023; 14(5):439. https://doi.org/10.3390/insects14050439
Chicago/Turabian StyleDavis, Abby E., Lena Alice Schmidt, Samantha Harrington, Cameron Spurr, and Romina Rader. 2023. "Provisioning Australian Seed Carrot Agroecosystems with Non-Floral Habitat Provides Oviposition Sites for Crop-Pollinating Diptera" Insects 14, no. 5: 439. https://doi.org/10.3390/insects14050439
APA StyleDavis, A. E., Schmidt, L. A., Harrington, S., Spurr, C., & Rader, R. (2023). Provisioning Australian Seed Carrot Agroecosystems with Non-Floral Habitat Provides Oviposition Sites for Crop-Pollinating Diptera. Insects, 14(5), 439. https://doi.org/10.3390/insects14050439