
Digestive Characteristics of Hyphantria cunea Larvae on Different Host Plants

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Host Plants
2.2. Experimental Insects
2.3. Treatment with Insect Digestive Enzyme Inhibitors
2.4. Body Weight and Food Utilization of H. cunea Larvae
2.5. Determination of Nutrient Contents in H. cunea Larvae
2.6. Determination of Digestive Enzyme Activity in the Midgut of H. cunea Larvae
2.7. Statistical Analysis
3. Results
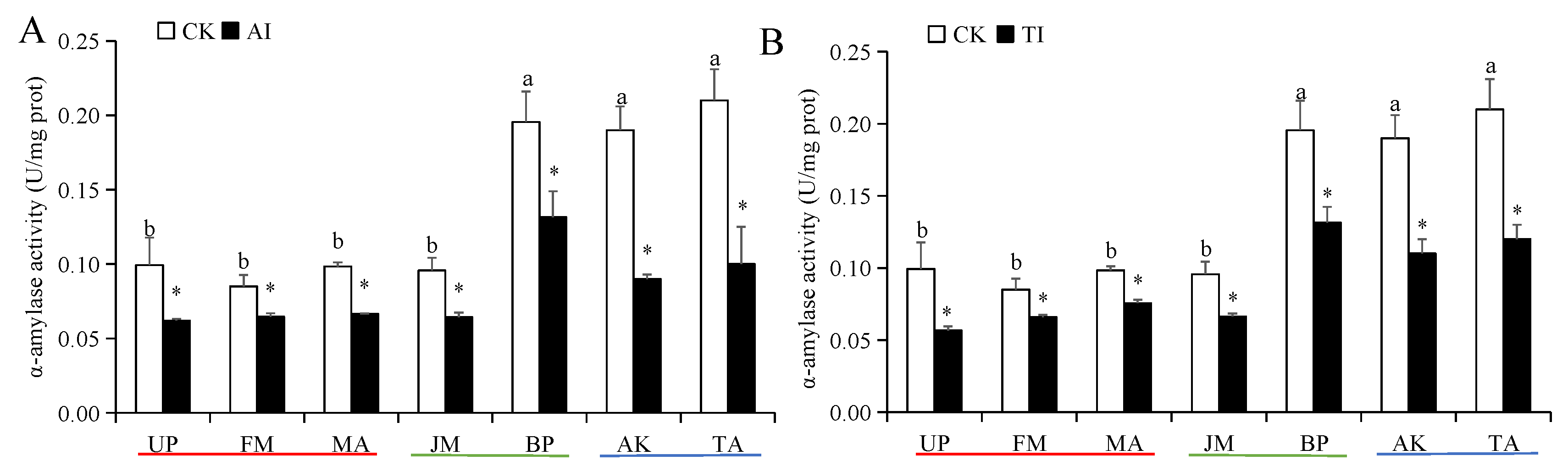
3.1. Digestive Enzyme Activity of H. cunea Larvae
3.2. Growth status of H. cunea Larvae
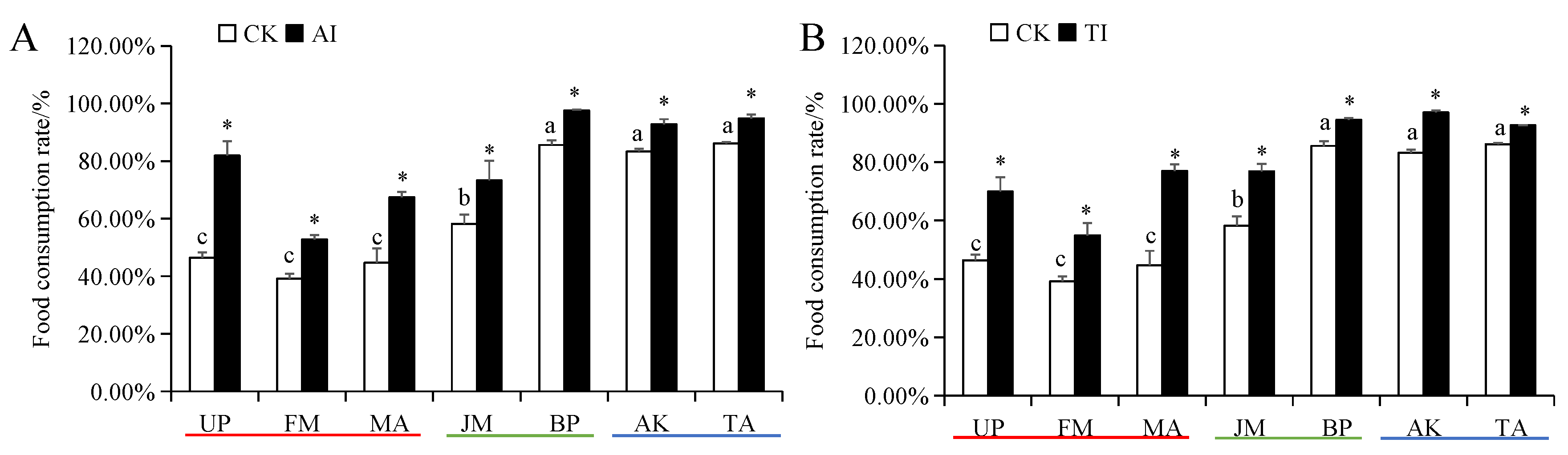
3.3. Food Utilization of H. cunea Larvae
3.4. Nutrient Contents of H. cunea Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; Fan, Z.Z.; Zhang, R.; Kong, X.B.; Liu, F.; Fang, J.X.; Zhang, S.F.; Zhang, Z. Bacteria-mediated RNAi for managing fall webworm, Hyphantria cunea: Screening target genes and analyzing lethal effect. Pest Manag. Sci. 2023, 79, 1261–1628. [Google Scholar] [CrossRef] [PubMed]
- Trajkovic, A.; Zikic, V. Stuck in the caterpillars’ web: A half-century of biocontrol research and application on gregarious Lepidopteran pests in Europe. Sustainability 2023, 15, 2881. [Google Scholar] [CrossRef]
- Sullivan, G.T.; Karaca, I.; Ozman-Sullivan, S.K.; Kara, K. Tachinid (Diptera:Tachinidae) parasitoids of overwintered Hyphantria cunea (Drury) (Lepidoptera: Arctiidae) pupae in hazelnut plantations in Samsun province, Turkey. J. Entomol. Res. Soc. 2012, 14, 21–30. [Google Scholar]
- Ge, X.Z.; He, S.Y.; Zhu, C.Y.; Wang, T.; Xu, Z.C.; Zong, S.X. Projecting the current and future potential global distribution of Hyphantria cunea (Lepidoptera: Arctiidae) using CLIMEX. Pest. Manag. Sci. 2019, 75, 160–169. [Google Scholar] [CrossRef]
- Zhao, X.D.; Geng, Y.S.; Hu, T.Y.; Li, W.X.; Liang, Y.Y.; Hao, D.J. Comparing the performance of Hyphantria cunea (Lepidoptera: Arctiidae) on artificial and natural diets: Feasibility of mass-rearing on artificial diets. J. Econ. Entomol. 2023, 116, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Bai, P.H.; Wang, H.M.; Liu, B.S.; Li, M.; Liu, B.M.; Gu, X.S.; Tang, R. Botanical volatiles selection in mediating electrophysiological responses and reproductive behaviors for the fall webworm moth Hyphantria cunea. Front. Physiol. 2020, 11, 486. [Google Scholar] [CrossRef] [PubMed]
- Schowalter, T.D.; Ring, D.R. Biology and management of the fall webworm, Hyphantria cunea (Lepidoptera: Erebidae). J. Integr. Pest Manag. 2017, 8, 7. [Google Scholar] [CrossRef]
- Tang, X.G.; Yuan, Y.D.; Liu, X.F.; Zhang, J.C. Potential range expansion and niche shift of the invasive Hyphantria cunea between native and invasive countries. Ecol. Entomol. 2021, 46, 910–925. [Google Scholar] [CrossRef]
- Cao, L.J.; Wei, S.J.; Hoffmann, A.A.; Wen, J.B.; Chen, M. Rapid genetic structuring of populations of the invasive fall webworm in relation to spatial expansion and control campaigns. Divers. Distrib. 2016, 22, 1276–1287. [Google Scholar] [CrossRef]
- Kim, D.E.; Kil, J.H. A report on the occurrence of and crop damage caused by Hyphantria cunea (Drury) with in Korea. Korean J. Appl. Entomol. 2012, 53, 285–293. [Google Scholar] [CrossRef]
- Chen, Q.; Zhao, H.; Wen, M.; Li, J.; Zhou, H.; Wang, J.; Zhou, Y.; Liu, Y.; Du, L.; Kang, H.; et al. Genome of the webworm Hyphantria cunea unveils genetic adaptations supporting its rapid invasion and spread. BMC Genom. 2020, 21, 242. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.I.; Park, Y.S. Dispersal patterns of exotic forest pests in South Korea. Insect Sci. 2012, 19, 535–548. [Google Scholar] [CrossRef]
- Wang, W.X.; Zhou, L.F.; Dong, G.P.; Chen, F.M. Isolation and identification of entomopathogenic fungi and an evaluation of their actions against the larvae of the fall webworm, Hyphantria cunea (Drury) (Lepidoptera: Arctiidae). BioControl 2020, 65, 101–111. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Zhang, S.S.; Niu, B.L.; Ji, D.F.; Liu, X.J.; Li, M.W.; Bai, H.; Palli, S.R.; Wang, C.Z.; Tan, A.J. A determining factor for insect feeding preference in the silkworm, Bombyx mori. PLoS Biol. 2019, 17, e3000162. [Google Scholar] [CrossRef]
- Jang, T.; Rho, M.S.; Koh, S.H.; Lee, K.P. Host-plant quality alters herbivore responses to temperature: A case study using the generalist Hyphantria cunea. Entomol. Exp. Appl. 2015, 154, 120–130. [Google Scholar] [CrossRef]
- Lazarevic, J.; Jankovic-Tomanic, M.; Savkovic, U.; Dordevic, M.; Milanovic, S.; Stojkovic, B. Host-associated divergence in the activity of digestive enzymes in two populations of the gypsy moth Lymantria dispar (Lepidoptera: Erebidae). J. Entomol. Sci. 2017, 20, 189–194. [Google Scholar] [CrossRef]
- Sarate, P.J.; Tamhane, V.A.; Kotkar, H.M.; Ratnakaran, N.; Susan, N.; Gupta, V.S.; Giri, A.P. Developmental and digestive flexibilities in the midgut of a polyphagous pest, the cotton bollworm, Helicoverpa armigera. J. Insect Sci. 2012, 12, 238–240. [Google Scholar] [CrossRef]
- Wu, N.N.; Zhang, S.F.; Li, X.W.; Cao, Y.H.; Liu, X.J.; Wang, Q.H.; Liu, Q.; Liu, H.H. Fall webworm genomes yield insights into rapid adaptation of invasive species. Nat. Ecol. Evol. 2019, 3, 105–115. [Google Scholar] [CrossRef]
- Li, L.S.; Yuan, Y.F.; Wu, L.; Chen, M. Effects of host plants on the feeding behavior and detoxification enzyme activities in Hyphantria cunea (Lepidoptera: Arctiidae) larvae. Acta Ecol. Sin. 2018, 61, 232–239. [Google Scholar]
- Mason, P.A.; Wilkes, S.R.; Lill, J.T.; Singer, M.S. Abundance trumps quality: Bi-trophic performance and parasitism risk fail to explain host use in the fall webworm. Oikos 2011, 120, 1509–1518. [Google Scholar] [CrossRef]
- Holtof, M.; Lenaerts, C.; Cullen, D.; Vanden Broeck, J. Extracellular nutrient digestion and absorption in the insect gut. Cell Tissue Res. 2019, 377, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.M.; Fang, J.; Zhao, J.Q.; Wang, K.; Ren, Y.J.; Meng, Z.J.; Yan, S.C. Effects of 8 host young trees on growth, development and nutritional efficiency of Hyphantria cunea larvae. For. Res. 2023, 35, 124–131. [Google Scholar]
- Yan, S.C.; Tan, M.T.; Zheng, L.; Wu, H.F.; Wang, K.; Chai, R.S.; Jiang, D. Defense response of Fraxinus mandshurica seedlings to Hyphantria cunea larvae under Cd stress: A contradiction between attraction and resistance. Sci. Total Environ. 2023, 859, 160390. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.L.; Pan, Z.Y.; Meng, X. The Effect of quercetin on the growth, development, nutrition utilization, and detoxifification enzymes in Hyphantria cunea Drury (Lepidoptera: Arctiidae). Forests 2022, 13, 1945. [Google Scholar] [CrossRef]
- Jiang, D.; Dong, X.W.; Yan, S.C. Heavy metal accumulation/excretion in and food utilization of Lymantria dispar larvae fed with Zn- or Pb-stressed Populus alba berolinensis leaves. Environ. Entomol. 2018, 47, 1329–1336. [Google Scholar] [CrossRef]
- Terra, W.R.; Barroso, I.G.; Dias, R.O.; Ferreira, C. Molecular physiology of insect midgut. Adv. Insect Physiol. 2019, 56, 366–418. [Google Scholar]
- Chen, S.X.; Yang, F.; Fang, M.; Yao, L.; Zheng, R.W.; Tang, Q.F. Effect of host plants on the biology and digestive physiology of Spodoptera frugiperda. Interciencia 2022, 47, 328–334. [Google Scholar]
- Teimouri, N.; Sendi, J.J.; Zibaee, A.; Khosravi, R. Feeding indices and enzymatic activities of carob moth Ectomyelois ceratoniae (Zeller) (Lepidoptera: Pyrallidae) on two commercial pistachio cultivars and an artificial diet. J. Saudi Soc. Agric. Sci. 2015, 14, 76–82. [Google Scholar] [CrossRef]
- Chai, Y.P.; Zhang, J.Y.; Wang, Y.X.; Jia, D.; Ma, R.Y.; Guo, Y.Q. Effects of starvation and different plant feedings on the digestive enzyme activities in theadults of alligator weed flea beetle Agasicles hygrophila. J. Integr. Plant Biol. 2016, 43, 138–144. [Google Scholar]
- Shen, H.Y.; Liang, Y.; Chen, Z.H.; Li, C.L.; Zhang, Z.H.; Weng, H.F.; Huang, B.; Zhang, F.P.; Liang, G.H. Effects of different host plants on the growth, reproduction and physiological enzyme activity of Dendrolimus houi Lajonquiere (Lepidoptera:Lasiocampidae). For. Res. 2022, 35, 63–70. [Google Scholar]
- Ashouri, S.; Pourabad, R.F. Regulation of gene expression encoding the digestive alpha-amylase in the larvae of Colorado potato beetle, Leptinotarsa decemlineata (Say) in response to plant protein extracts. Gene 2021, 766, 145159. [Google Scholar] [CrossRef] [PubMed]
- Da Lage, J.-L. The Amylases of insects. Int. J. Insect Sci. 2018, 10, 1179543318804783. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.J.; Song, Y.; Jiang, X.M.; He, L.; Wei, L.Y.; Zhao, Z.W. Synergism of feeding and digestion regulated by the neuropeptide F system in Ostrinia furnacalis Larvae. Cells 2023, 12, 194. [Google Scholar] [CrossRef] [PubMed]
- Lazarevic, J.; Jankovic-Tomanic, M. Dietary and phylogenetic correlates of digestive trypsin activity in insect pests. Entomol. Exp. Appl. 2015, 157, 123–151. [Google Scholar] [CrossRef]
- Tan, M.T.; Wu, H.F.; Yan, S.C.; Jiang, D. Evaluating the toxic effects of tannic acid treatment on Hyphantria cunea larvae. Insects 2022, 13, 872. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Tan, M.T.; Yan, S.C.; Jiang, D. Cadmium exposure-triggered growth retardation in Hyphantria cunea larvae involves disturbances in food utilization and energy metabolism. Ecotoxicol. Environ. Saf. 2023, 256, 114886. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.S.; Zheng, L.; Tan, M.T.; Wu, H.F.; Yan, S.C.; Jiang, D. The susceptibility of Hyphantria cunea larvae to microbial pesticides Bacillus thuringiensis and Mamestra brassicae nuclear polyhedrosis virus under Cd stress. Pestic. Biochem. Physiol. 2023, 191, 105383. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.L.; Zhou, L.F.; Wang, Q.Y.; Ding, J.Q. Plant chemistry determines host preference and performance of an invasive insect. Front. Plant Sci. 2020, 11, 594663. [Google Scholar] [CrossRef]
- Vengateswari, G.; Arunthirumeni, M.; Shivaswamy, M.S.; Shivakumar, M.S. Effect of host plants nutrients, antioxidants, and phytochemicals on growth, development, and fecundity of Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae). Int. J. Trop. Insect Sci. 2022, 42, 3161–3173. [Google Scholar] [CrossRef]
- Gou, Y.P.; Quandahor, P.; Zhang, Y.X.; Coulter, J.A.; Liu, C.Z. Host plant nutrient contents influence nutrient contents in Bradysia cellarum and Bradysia impatiens. PLoS ONE 2020, 15, e0226471. [Google Scholar] [CrossRef]
- Hafeez, M.; Li, X.W.; Zhang, J.M.; Zhang, Z.J.; Huang, J.; Wang, L.K.; Khan, M.M.; Shah, S.K.; Fernandez-Grandon, G.M.; Lu, Y.B. Role of digestive protease enzymes and related genes in host plant adaptation of a polyphagous pest, Spodoptera frugiperda. Insect Sci. 2021, 28, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Babamir-Satehi, A.; Habibpour, B.; Aghdam, H.R.; Hemmati, S.A. Interaction between feeding efficiency and digestive physiology of the pink stem borer, Sesamia cretica Lederer (Lepidoptera: Noctuidae), and biochemical compounds of different sugarcane cultivars. Arthropod-Plant Interact. 2022, 16, 309–316. [Google Scholar] [CrossRef]
- Barzin, S.; Naseri, B.; Fathi, S.A.A.; Razmjou, J.; Aeinehchi, P. Feeding efficiency and digestive physiology of Trogoderma granarium Everts (Coleoptera: Dermestidae) on different rice cultivars. J. Stored Prod. Res. 2019, 84, 101511. [Google Scholar] [CrossRef]
- Mendonca, E.G.; Barros, R.D.; Cordeiro, G.; Silva, C.R.; Campos, W.G.; de Oliveira, J.A.; Oliveira, M.G.D. Larval development and proteolytic activity of Anticarsia gemmatalis Hubner (Lepidoptera: Noctuidae) exposed to different soybean protease inhibitors. Arch. Insect Biochem. Physiol. 2020, 103, e21637. [Google Scholar] [CrossRef] [PubMed]
- Juma, G.; Le, R.B.; Calatayud, P.A. Assortments of digestive enzymes induced in first instar larvae of Busseola fusca feeding on different plants. Int. J. Insect Sci. 2019, 11, 1179543319843521. [Google Scholar] [CrossRef]
- Bezerra, C.; Oliveira, C.T.; Macedo, M. Inga vera trypsin inhibitor interferes in the proteolytic activity and nutritional physiology of Ephestia kuehniella larvae. Entomol. Exp. Appl. 2017, 165, 109–119. [Google Scholar] [CrossRef]
- Kuwar, S.S.; Pauchet, Y.; Heckel, D.G. Effects of class-specific, synthetic, and natural proteinase inhibitors on life-history traits of the cotton bollworm Helicoverpa armigera. Arch. Insect Biochem. Physiol. 2020, 103, e21647. [Google Scholar] [CrossRef]
- Macedo, M.L.R.; de Oliveira, C.F.R.; Costa, P.M.; Castelhano, E.C.; Silva-Filho, M.C. Adaptive mechanisms of insect pests against plant protease inhibitors and future prospects related to crop protection: A review. Protein Pept. Lett. 2015, 22, 149–163. [Google Scholar] [CrossRef]
- Zhu-Salzman, K.; Zeng, R. Insect Responses to Plant Defensive Protease Inhibitors. Annu. Rev. Entomol. 2015, 60, 233–252. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient Content | UP | FM | MA | JM | BP | AK | TA | |
|---|---|---|---|---|---|---|---|---|
| Total protein (mg/g) | CK | 10.53 ± 0.65 ab | 11.55 ± 0.41 ab | 10.02 ± 0.25 b | 12.11 ± 0.41 a | 7.37 ± 0.63 c | 7.66 ± 0.66 c | 7.08 ± 0.88 c |
| AI | 8.19 ± 0.18 * | 9.60 ± 0.47 * | 7.33 ± 0.61 * | 14.83 ± 0.20 * | 13.77 ± 1.24 * | 10.73 ± 0.27 * | 8.18 ± 0.53 | |
| TI | 7.70 ± 0.75 * | 9.12 ± 0.54 * | 8.08 ± 0.60 * | 17.05 ± 0.66 * | 14.02 ± 1.11 * | 11.32 ± 1.14 * | 8.63 ± 0.30 | |
| Total amino acids (μmol/mg prot) | CK | 1.92 ± 0.19 a | 1.90 ± 0.12 a | 1.80 ± 0.05 ab | 1.83 ± 0.13 ab | 1.52 ± 0.19 abc | 1.41 ± 0.13 c | 1.18 ± 0.05 c |
| AI | 2.31 ± 0.20 * | 1.20 ± 0.07 * | 2.00 ± 0.01 * | 1.09 ± 0.14 * | 0.85 ± 0.11 * | 0.96 ± 0.01 * | 1.02 ± 0.01 * | |
| TI | 1.94 ± 0.19 | 1.26 ± 0.14 * | 2.30 ± 0.04 * | 1.07 ± 0.02 * | 0.96 ± 0.07 * | 0.84 ± 0.10 * | 0.86 ± 0.03 * | |
| Glucose (μmol/g prot) | CK | 110.30 ± 3.41 a | 120.90 ± 11.36 a | 110.90 ± 3.31 a | 82.20 ± 15.37 b | 55.20 ± 1.56 b | 62.00 ± 6.61 b | 67.60 ± 3.80 b |
| AI | 40.70 ± 1.97 * | 85.50 ± 1.22 * | 88.90 ± 4.83 * | 48.10 ± 3.68 * | 27.80 ± 1.74 * | 41.70 ± 7.66 * | 37.70 ± 1.50 * | |
| TI | 62.40 ± 3.06 * | 76.80 ± 7.96 * | 81.95 ± 4.97 * | 43.50 ± 6.60 * | 36.90 ± 6.09 * | 52.60 ± 1.57 * | 53.90 ± 1.80 * | |
| Trehalose (mg/g tissue) | CK | 8.69 ± 0.30 a | 9.26 ± 0.29 b | 7.23 ± 0.46 c | 5.75 ± 0.13 d | 2.50 ± 0.21 e | 1.24 ± 0.11 f | 3.82 ± 0.41 g |
| AI | 2.43 ± 0.12 * | 4.50 ± 0.52 * | 1.81 ± 0.16 * | 3.60 ± 0.44 * | 0.74 ± 0.35 * | 0.53 ± 0.15 * | 0.60 ± 0.18 * | |
| TI | 2.48 ± 0.24 * | 4.21 ± 0.37 * | 3.50 ± 0.28 * | 3.34 ± 0.58 * | 0.85 ± 0.57 * | 0.38 ± 0.04 * | 0.78 ± 0.23 * | |
| Free fatty acids (μmol/g prot) | CK | 114.60 ± 11.28 a | 55.00 ± 3.40 b | 105.60 ± 7.98 a | 49.00 ± 11.30 b | 6.30 ± 1.28 c | 11.20 ± 1.60 c | 9.40 ± 2.00 c |
| AI | 152.90 ± 6.38 * | 79.40 ± 11.24 * | 223.90 ± 6.15 * | 80.20 ± 5.09 * | 1.70 ± 0.21 * | 2.60 ± 0.35 * | 3.10 ± 0.54 * | |
| TI | 157.20 ± 13.98 * | 79.20 ± 7.09 * | 269.80 ± 9.98 * | 84.10 ± 8.50 * | 2.80 ± 0.91 * | 2.80 ± 1.94 * | 3.10 ± 1.58 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, A.; Li, T.; Yuan, L.; Tan, M.; Jiang, D.; Yan, S. Digestive Characteristics of Hyphantria cunea Larvae on Different Host Plants. Insects 2023, 14, 463. https://doi.org/10.3390/insects14050463
Zhang A, Li T, Yuan L, Tan M, Jiang D, Yan S. Digestive Characteristics of Hyphantria cunea Larvae on Different Host Plants. Insects. 2023; 14(5):463. https://doi.org/10.3390/insects14050463
Chicago/Turabian StyleZhang, Aoying, Tao Li, Lisha Yuan, Mingtao Tan, Dun Jiang, and Shanchun Yan. 2023. "Digestive Characteristics of Hyphantria cunea Larvae on Different Host Plants" Insects 14, no. 5: 463. https://doi.org/10.3390/insects14050463
APA StyleZhang, A., Li, T., Yuan, L., Tan, M., Jiang, D., & Yan, S. (2023). Digestive Characteristics of Hyphantria cunea Larvae on Different Host Plants. Insects, 14(5), 463. https://doi.org/10.3390/insects14050463