

Spent Coffee Grounds and Novaluron Are Toxic to Aedes aegypti (Diptera: Culicidae) Larvae
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquitoes
2.2. Spent Coffee Grounds
2.3. Novaluron
2.4. Bioassay
2.5. Combination of wSCGs and Novaluron
2.6. Statistical Analysis
3. Results
3.1. Characteristics and Quantification of wSCGs and dSCGs
3.2. Comparison of Larval Mortality and Adult Emergence Inhibition with wSCGs and dSCGs
3.3. Bioassay of Individual and Combination of wSCGs and Novaluron
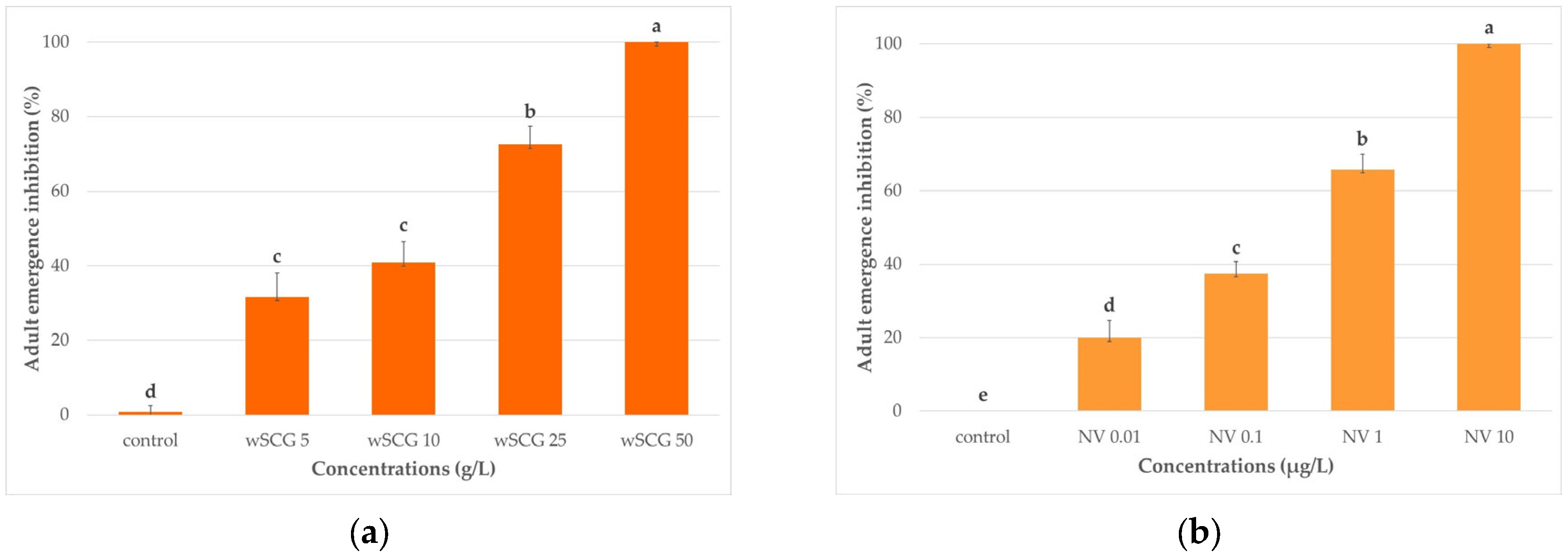
3.3.1. Individual Efficacies of wSCGs and Novaluron
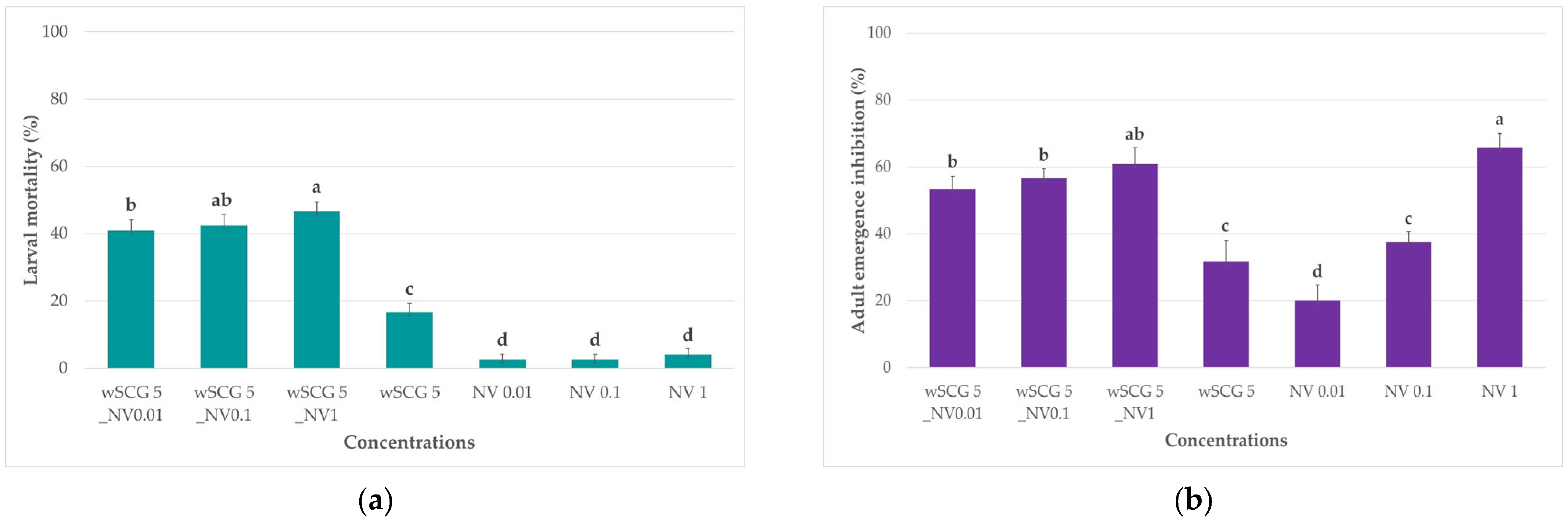
3.3.2. Synergistic Effects of Sublethal Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Vector-Borne Diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 2 March 2022).
- World Health Organization. Dengue and Severe Dengue. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 10 January 2022).
- World Health Organization, Regional Office for South-East Asia. Epidemiology of Dengue Fever and Dengue Haemorrhagic Fever. In Comprehensive Guideline for Prevention and Control of Dengue and Dengue Haemorrhagic Fever, Revised and Expanded ed.; World Health Organization, Regional Office for South-East Asia: New Delhi, India, 2011; pp. 9–15. Available online: https://apps.who.int/iris/handle/10665/204894 (accessed on 10 January 2022).
- Sornpeng, W.; Pimsamarn, S.; Akksilp, S. Resistance to Temephos of Aedes aegypti Linnaeus larvae (Diptera: Culicidae). KKU Res. J. 2009, 14, 650–654. [Google Scholar]
- Department of Disease Control. Mosquito Larvae Susceptibility Test Report. 2021. Available online: https://drive.google.com/drive/folders/1wYTCr6ftTZfAnEMN8a8OIoOufJlSgTTw (accessed on 26 May 2022).
- Department of Disease Control. Mosquito Larvae Susceptibility Test Report. 2022. Available online: https://drive.google.com/drive/folders/1keTNEWdfAv7pFiWK67QdLqo6BEaNl6x9 (accessed on 3 February 2023).
- Grisales, N.; Poupardin, R.; Gomez, S.; Fonseca-Gonzalez, I.; Ranson, H.; Lenhart, A. Temephos resistance in Aedes aegypti in Colombia compromises dengue vector control. PLoS Negl. Trop. Dis. 2013, 7, e2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomino, M.; Pinto, J.; Yañez, P.; Cornelio, A.; Dias, L.; Amorim, Q.; Lenhart, A.; Lima, J.B.P. First national-scale evaluation of temephos resistance in Aedes aegypti in Peru. Parasites Vectors 2022, 15, 254. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.; Ponce, P.; Cevallos, V.; Espinosa, P.; Vaca, D.; Quezada, W. Resistance status of Aedes aegypti to deltamethrin, malathion, and temephos in Ecuador. J. Am. Mosq. Control Assoc. 2019, 35, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamil, M.; Ramadan, K.M.; Awad, O.I.; Ibrahim, T.K.; Inayat, A.; Ma, X. Environmental impacts of biodiesel production from waste spent coffee grounds and its implementation in a compression ignition engine. Sci. Total Environ. 2019, 675, 13–30. [Google Scholar] [CrossRef]
- Martinez-Saez, N.; García, A.T.; Pérez, I.D.; Rebollo-Hernanz, M.; Mesías, M.; Morales, F.J.; Martín-Cabrejas, M.A.; Del Castillo, M.D. Use of spent coffee grounds as food ingredient in bakery products. Food Chem. 2017, 216, 114–122. [Google Scholar] [CrossRef]
- Vardon, D.R.; Moser, B.R.; Zheng, W.; Witkin, K.; Evangelista, R.L.; Strathmann, T.J.; Rajagopalan, K.; Sharma, B.K. Complete utilization of spent coffee grounds to produce biodiesel, bio-oil, and biochar. ACS Sustain. Chem. Eng. 2013, 1, 1286–1294. [Google Scholar] [CrossRef]
- Ronga, D.; Pane, C.; Zaccardelli, M.; Pecchioni, N. Use of spent coffee ground compost in peat-based growing media for the production of basil and tomato potting plants. Commun. Soil Sci. Plant Anal. 2016, 47, 356–368. [Google Scholar] [CrossRef]
- Guirado, M.M.; de Campos Bicudo, H.E.M. Effect of used coffee grounds on larval mortality of Aedes aegypti L. (Diptera: Culicidae): Suspension concentration and age versus efficacy. BioAssay 2007, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Laranja, A.T.; Manzatto, A.J.; de Campos Bicudo, H.E.M. Effects of caffeine and used coffee grounds on biological features of Aedes aegypti (Diptera, Culicidae) and their possible use in alternative control. Genet. Mol. Biol. 2003, 26, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Aditama, W.; Zulfikar, S.F.Y. The effectiveness of arabica coffee (Coffea arabica L.) grounds on mortality and growth of Aedes aegypti larva. Int. J. Mosq. Res. 2019, 6, 34–37. [Google Scholar]
- Cutler, G.C.; Scott-Dupree, C.D. Novaluron: Prospects and limitations in insect pest management. Pest Technol. 2007, 1, 38–46. [Google Scholar]
- Matsumura, F. Studies on the action mechanism of benzoylurea insecticides to inhibit the process of chitin synthesis in insects: A review on the status of research activities in the past, the present and the future prospects. Pestic. Biochem. Physiol. 2010, 97, 133–139. [Google Scholar] [CrossRef]
- Sun, R.; Liu, C.; Zhang, H.; Wang, Q. Benzoylurea chitin synthesis inhibitors. J. Agric. Food Chem. 2015, 63, 6847–6865. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAO Specifications and Evaluations for Plant Protection Products “Novaluron (±)-1-[3-Chloro-4-(1,1,2-trifluoro-2-trifluoromethoxyethoxy)phenyl]-3-(2,6-difluorobenzoyl)urea”. Available online: https://www.fao.org/fileadmin/templates/agphome/documents/Pests_Pesticides/Specs/novaluro.pdf (accessed on 2 March 2022).
- World Health Organization. Report of the Eighth WHOPES Working Group Meeting. Review of Novaluron 10% EC. Available online: http://whqlibdoc.who.int/hq/2005/WHO_CDS_WHOPES_2005.10.pdf (accessed on 2 March 2022).
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.-P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef]
- Malla, M.Y.; Sharma, M.; Saxena, R.; Mir, M.I.; Mir, A.H.; Bhat, S.H. Phytochemical screening and spectroscopic determination of total phenolic and flavonoid contents of Eclipta alba Linn. J. Nat. Prod. Plant Resour. 2013, 3, 86–91. [Google Scholar]
- Butsat, S.; Siriamornpun, S. Phenolic acids and antioxidant activities in husk of different Thai rice varieties. Food Sci. Technol. Int. 2010, 16, 329–336. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Drago, A.; Vettore, S.; Martini, S.; Dutto, M. Efficacy of used coffee grounds as larvicide against Aedes albopictus (Skuse, 1894) and Ae. aegypti Linné, 1762 (Diptera: Culicidae). J. Eur. Mosq. Control Assoc. 2021, 39, 27–30. [Google Scholar] [CrossRef]
- Derraik, J.G.; Slaney, D. The toxicity of used coffee grounds to the larvae of Ochlerotatus (Finlaya) notoscriptus (Skuse) (Diptera: Culicidae). Ann. Med. Entomol. 2005, 14, 14–24. [Google Scholar]
- Miranda, F.R.; Fernandes, K.M.; Bernardes, R.C.; Martins, G.F. Biological, histological and immunohistochemical studies on the toxicity of spent coffee grounds and caffeine on the larvae of Aedes aegypti (Diptera: Culicidae). Environ. Pollut. 2021, 271, 116307. [Google Scholar] [CrossRef] [PubMed]
- Tangtrakulwanich, K.; Suwannawong, B.; Nakrung, P. The Comparative study of arabica used coffee grounds and temephos in controlling the Aedes aegypti larvae. J. Food Sci. Agric. Technol. 2022, 6, 34–37. [Google Scholar]
- Hussein, H.; Abouamer, W.; Ali, H.; Elkhadragy, M.; Yehia, H.; Farouk, A. The valorization of spent coffee ground extract as a prospective insecticidal agent against some main key pests of Phaseolus vulgaris in the laboratory and field. Plants 2022, 11, 1124. [Google Scholar] [CrossRef]
- Socała, K.; Szopa, A.; Serefko, A.; Poleszak, E.; Wlaź, P. Neuroprotective effects of coffee bioactive compounds: A review. Int. J. Mol. Sci. 2020, 22, 107. [Google Scholar] [CrossRef]
- Campos-Vega, R.; Loarca-Piña, G.; Vergara-Castañeda, H.A.; Oomah, B.D. Spent coffee grounds: A review on current research and future prospects. Trends Food Sci. Technol. 2015, 45, 24–36. [Google Scholar] [CrossRef]
- Pratita, A.T.K. Skrining fitokimia dan analisis kromatografi lapis tipis senyawa alkaloid dari berbagai ekstrak kopi robusta (Coffea canephora). J. Kesehat. Bakti Tunas Husada J. Ilmu-Ilmu Keperawatan Anal. Kesehat. Farm. 2018, 17, 198–201. [Google Scholar] [CrossRef] [Green Version]
- Simmonds, M.S. Flavonoid–insect interactions: Recent advances in our knowledge. Phytochemistry 2003, 64, 21–30. [Google Scholar] [CrossRef]
- Summers, C.B.; Felton, G.W. Prooxidant effects of phenolic acids on the generalist herbivore Helicoverpa zea (Lepidoptera: Noctuidae): Potential mode of action for phenolic compounds in plant anti-herbivore chemistry. Insect Biochem. Mol. Biol. 1994, 24, 943–953. [Google Scholar] [CrossRef]
- Nakhaie Bahrami, M.; Mikani, A.; Moharramipour, S. Effect of caffeic acid on feeding, α-amylase and protease activities and allatostatin—A content of Egyptian cotton leafworm, Spodoptera littoralis (Lepidoptera: Noctuidae). J. Pestic. Sci. 2018, 43, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, R.S.; Wagh, T.P.; Sharma, N.; Mulani, F.A.; Sonavane, U.; Thulasiram, H.V.; Joshi, R.; Gupta, V.S.; Giri, A.P. Way toward “dietary pesticides”: Molecular investigation of insecticidal action of caffeic acid against Helicoverpa armigera. J. Agric. Food Chem. 2014, 62, 10847–10854. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Hussain, B.; Buhroo, A.A.; Ignacimuthu, S.; Sharma, H.C. Effect of plant secondary metabolites on legume pod borer, Helicoverpa armigera. J. Pest Sci. 2013, 86, 399–408. [Google Scholar] [CrossRef]
- Hisham, S.M.; Mohammad, A.M.; Mohammed, M.J. The effect of extracts and phenolic compounds isolation from Rosmarinus officinalis plant leaves on Tribolium castaneum mortality. Int. J. Drug Deliv. Technol. 2022, 12, 814–819. [Google Scholar] [CrossRef]
- Sambangi, P.; Rani, P.U. Physiological effects of resveratrol and coumaric acid on two major groundnut pests and their egg parasitoid behavior. Arch. Insect Biochem. Physiol. 2016, 91, 230–245. [Google Scholar] [CrossRef]
- Muñoz, I.J.; Schilman, P.E.; Barrozo, R.B. Impact of alkaloids in food consumption, metabolism and survival in a blood-sucking insect. Sci. Rep. 2020, 10, 9443. [Google Scholar] [CrossRef]
- Bhuvaneswari, K.; Raghavan, G.; Thangavel, K. Use of caffeine for the management of rice weevil Sitophilus oryzae (L.). Intern. J. Agric. Sci. 2007, 3, 269–272. [Google Scholar]
- Mulla, M.S.; Thavara, U.; Tawatsin, A.; Chompoosri, J.; Zaim, M.; Su, T. Laboratory and field evaluation of novaluron, a new acylurea insect growth regulator, against Aedes aegypti (Diptera: Culicidae). J. Vector Ecol. 2003, 28, 241–254. [Google Scholar]
- Arredondo-Jimenez, J.I.; Valdez-Delgado, K.M. Effect of novaluron (Rimon® 10 EC) on the mosquitoes Anopheles albimanus, Anopheles pseudopunctipennis, Aedes aegypti, Aedes albopictus and Culex quinquefasciatus from Chiapas, Mexico. Med. Vet. Entomol. 2006, 20, 377–387. [Google Scholar] [CrossRef]
- Fiaz, M.; Martínez, L.C.; Plata-Rueda, A.; Cossolin, J.F.S.; Serra, R.S.; Martins, G.F.; Serrão, J.E. Behavioral and ultrastructural effects of novaluron on Aedes aegypti larvae. Infect. Genet. Evol. 2021, 93, 104974. [Google Scholar] [CrossRef]
- NourElhouda, D.; Hamid, B.; Ali, B.; Noureddine, S. Biological effects of a benzoylphenylurea derivative (novaluron) on larvae of Culex pipiens (Diptera: Culicidae). Adv. Appl. Sci. Res. 2013, 4, 449–456. [Google Scholar]
- Tawatsin, A.; Thavara, U.; Bhakdeenuan, P.; Chompoosri, J.; Siriyasatien, P.; Asavadachanukorn, P.; Mulla, M.S. Field evaluation of novaluron, a chitin synthesis inhibitor larvicide, against mosquito larvae in polluted water in urban areas of Bangkok, Thailand. Southeast Asian J. Trop. Med. 2007, 38, 434–441. [Google Scholar]
- Rojrutha, S.; Klunreaungsang, R.; Adisaimontr, S.; Samun, Y.; Bunjun, B. Culex spp. mosquito larva control by appling novaluron in polluted water. Dis. Control J. 2012, 38, 138–146. [Google Scholar]
- Kalyanasundaram, M.; Das, P. Larvicidal and synergistic activity of plant extracts for mosquito control. Indian J. Med. Res. 1985, 82, 19–23. [Google Scholar] [PubMed]
- Chansang, A.; Champakaew, D.; Junkum, A.; Jitpakdi, A.; Amornlerdpison, D.; Aldred, A.K.; Riyong, D.; Wannasan, A.; Intirach, J.; Muangmoon, R. Synergy in the adulticidal efficacy of essential oils for the improvement of permethrin toxicity against Aedes aegypti L. (Diptera: Culicidae). Parasities Vectors 2018, 11, 417. [Google Scholar] [CrossRef] [Green Version]
- Rybak, M.E.; Sternberg, M.R.; Pao, C.I.; Ahluwalia, N.; Pfeiffer, C.M. Urine excretion of caffeine and select caffeine metabolites is common in the U.S. population and associated with caffeine intake. J. Nutr. 2015, 145, 766–774. [Google Scholar] [CrossRef] [Green Version]
- Petrovic, D.; Estoppey Younes, S.; Pruijm, M.; Ponte, B.; Ackermann, D.; Ehret, G.; Ansermot, N.; Mohaupt, M.; Paccaud, F.; Vogt, B. Relation of 24-hour urinary caffeine and caffeine metabolite excretions with self-reported consumption of coffee and other caffeinated beverages in the general population. Nutr. Metab. 2016, 13, 81. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.P. Caffeine as an environmental indicator for assessing urban aquatic ecosystems. Cad. Saude Publica 2005, 21, 1884–1892. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.D.S.; Maranho, L.A.; Cortez, F.S.; Pusceddu, F.H.; Santos, A.R.; Ribeiro, D.A.; Cesar, A.; Guimarães, L.L. Occurrence of pharmaceuticals and cocaine in a Brazilian coastal zone. Sci. Total Environ. 2016, 548, 148–154. [Google Scholar] [CrossRef]
- Zhou, H.; Wu, C.; Huang, X.; Gao, M.; Wen, X.; Tsuno, H.; Tanaka, H. Occurrence of selected pharmaceuticals and caffeine in sewage treatment plants and receiving rivers in Beijing, China. Water Environ. Res. 2010, 82, 2239–2248. [Google Scholar] [CrossRef]
- Komori, K.; Suzuki, Y.; Minamiyama, M.; Harada, A. Occurrence of selected pharmaceuticals in river water in Japan and assessment of their environmental risk. Environ. Monit. Assess. 2013, 185, 4529–4536. [Google Scholar] [CrossRef] [PubMed]
- Stamatis, N.K.; Konstantinou, I.K. Occurrence and removal of emerging pharmaceutical, personal care compounds and caffeine tracer in municipal sewage treatment plant in Western Greece. J. Environ. Sci. Health Part B 2013, 48, 800–813. [Google Scholar] [CrossRef]
- Fang, T.H.; Lin, C.W.; Kao, C.H. Occurrence and distribution of pharmaceutical compounds in the Danshuei River Estuary and the Northern Taiwan Strait. Mar. Pollut. Bull. 2019, 146, 509–520. [Google Scholar] [CrossRef]
- Biel-Maeso, M.; Baena-Nogueras, R.M.; Corada-Fernández, C.; Lara-Martín, P.A. Occurrence, distribution and environmental risk of pharmaceutically active compounds (PhACs) in coastal and ocean waters from the Gulf of Cadiz (SW Spain). Sci. Total Environ. 2018, 612, 649–659. [Google Scholar] [CrossRef] [PubMed]
- del Rey, Z.R.; Granek, E.F.; Sylvester, S. Occurrence and concentration of caffeine in Oregon coastal waters. Mar. Pollut. Bull. 2012, 64, 1417–1424. [Google Scholar] [CrossRef]
- Ojemaye, C.Y.; Petrik, L. Occurrences, levels and risk assessment studies of emerging pollutants (pharmaceuticals, perfluoroalkyl and endocrine disrupting compounds) in fish samples from Kalk Bay harbour, South Africa. Environ. Pollut. 2019, 252, 562–572. [Google Scholar] [CrossRef]
- Ali, A.M.; Rønning, H.T.; Sydnes, L.K.; Alarif, W.M.; Kallenborn, R.; Al-Lihaibi, S.S. Detection of PPCPs in marine organisms from contaminated coastal waters of the Saudi Red Sea. Sci. Total Environ. 2018, 621, 654–662. [Google Scholar] [CrossRef]
- Bruton, T.; Alboloushi, A.; De La Garza, B.; Kim, B.O.; Halden, R.U. Fate of caffeine in the environment and ecotoxicological considerations. In Contaminants of Emerging Concern in the Environment: Ecological and Human Health Considerations; Halden, R.U., Ed.; American Chemical Society: Washington, DC, USA, 2010; pp. 257–273. [Google Scholar]
- Lam, M.W.; Young, C.J.; Brain, R.A.; Johnson, D.J.; Hanson, M.A.; Wilson, C.J.; Richards, S.M.; Solomon, K.R.; Mabury, S.A. Aquatic persistence of eight pharmaceuticals in a microcosm study. Environ. Toxicol. Chem. 2004, 23, 1431–1440. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, S.; Shukor, M.Y.; Syed, M.A.; Ab Rahman, N.A.; Khalil, K.A.; Khalid, A.; Ahmad, S.A. Bacterial degradation of caffeine: A review. Am. J. Plant Biol. 2014, 2, 19–28. [Google Scholar] [CrossRef]
- Korekar, G.; Kumar, A.; Ugale, C. Occurrence, fate, persistence and remediation of caffeine: A review. Environ. Sci. Pollut. Res. 2020, 27, 34715–34733. [Google Scholar] [CrossRef]
- Rathnamma, V.V. Histopathological changes in the gill and liver of freshwater fish Labeo rohita (Hamilton) exposed to novaluron. Innoriginal Int. J. Sci. 2014, 1, 16–18. [Google Scholar]
- Berghiche, H.; Benradia, H.; Soltani, N. Impact of insect growth disruptor, novaluron, on biochemical composition of cuticle from the shrimp Palaemon adspersus. J. Entomol. Zool. 2016, 4, 147–151. [Google Scholar]
- Berghiche, H.; Benradia, H.; Soltani, N. Evaluation of the potential side-effects of novaluron on the shrimp Palaemon adspersus: Moulting hormone profile, cuticle secretion and chitin contents. Int. J. Environ. Monit. Anal. 2018, 6, 116–124. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Phytochemicals | Chemicals | Units | wSCGs | dSCGs |
|---|---|---|---|---|
| Alkaloid compound | Caffeine | %w/w ** | 0.2 | 0.4 |
| Flavonoid compounds | Total flavonoid compounds | mg/g | 7.3 ± 0.1 | 1.1 ± 0.1 |
| Phenolic compounds | Total phenolic compounds | mg/g | 50.7 ± 0.9 | 5.2 ± 0.1 |
| Caffeic acid | µg/g | 329.7 ± 0.4 | 8.6 ± 2.3 | |
| Coumaric acid | µg/g | 120.8 ± 3.1 | 5.8 ± 0.3 | |
| Protocatechuic acid | µg/g | 41.1 ± 30.9 | 9.1 ± 3.0 | |
| Vanillic acid | µg/g | 5597.0 ± 213.8 | 770.3 ± 22.5 | |
| Ferulic acid | µg/g | ND * | ND * | |
| Gallic acid | µg/g | ND * | ND * | |
| Sinapic acid | µg/g | ND * | ND * |
| Times (h) | Treatments | Larval Mortality Rates ( ± SD) | Treatments | Larval Mortality Rates ( ± SD) | U | p |
|---|---|---|---|---|---|---|
| 72 | control | 0.8 ± 1.7 | control | 0.8 ± 1.7 | 8.0 | 1.000 |
| wSCG 5 | 16.7 ± 2.7 | dSCG 5 | 8.3 ± 1.9 | 0.0 | 0.019 | |
| wSCG 10 | 24.2 ± 3.2 | dSCG 10 | 10.8 ± 3.2 | 0.0 | 0.019 | |
| wSCG 25 | 54.2 ± 3.2 | dSCG 25 | 14.2 ± 4.2 | 0.0 | 0.019 | |
| wSCG 50 | 100 | dSCG 50 | 14.2 ± 5.0 | 0.0 | 0.013 |
| Times (h) | Treatments | Adult Emergence Inhibition Rates ( ± SD) | Treatments | Adult Emergence Inhibition Rates ( ± SD) | U | p |
|---|---|---|---|---|---|---|
| 120 | control | 0.8 ± 1.7 | control | 0.8 ± 1.7 | 8.0 | 1.000 |
| wSCG 5 | 31.7 ± 6.4 | dSCG 5 | 14.2 ± 3.2 | 0.0 | 0.019 | |
| wSCG 10 | 40.8 ± 5.7 | dSCG 10 | 16.7 ± 6.7 | 0.0 | 0.018 | |
| wSCG 25 | 72.5 ± 5.0 | dSCG 25 | 24.2 ± 1.7 | 0.0 | 0.015 | |
| wSCG 50 | 100 | dSCG 50 | 25.0 ± 7.9 | 0.0 | 0.013 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thanasoponkul, W.; Changbunjong, T.; Sukkurd, R.; Saiwichai, T. Spent Coffee Grounds and Novaluron Are Toxic to Aedes aegypti (Diptera: Culicidae) Larvae. Insects 2023, 14, 564. https://doi.org/10.3390/insects14060564
Thanasoponkul W, Changbunjong T, Sukkurd R, Saiwichai T. Spent Coffee Grounds and Novaluron Are Toxic to Aedes aegypti (Diptera: Culicidae) Larvae. Insects. 2023; 14(6):564. https://doi.org/10.3390/insects14060564
Chicago/Turabian StyleThanasoponkul, Waralee, Tanasak Changbunjong, Rattanavadee Sukkurd, and Tawee Saiwichai. 2023. "Spent Coffee Grounds and Novaluron Are Toxic to Aedes aegypti (Diptera: Culicidae) Larvae" Insects 14, no. 6: 564. https://doi.org/10.3390/insects14060564
APA StyleThanasoponkul, W., Changbunjong, T., Sukkurd, R., & Saiwichai, T. (2023). Spent Coffee Grounds and Novaluron Are Toxic to Aedes aegypti (Diptera: Culicidae) Larvae. Insects, 14(6), 564. https://doi.org/10.3390/insects14060564