
First Records of Heartbeats via ECG in a Stingless Bee, Melipona flavolineata (Apidae, Meliponini), during Contention Stress Using Isoflurane as an Anesthetic

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Studied Species
2.2. Animal Preparation for the Experiment
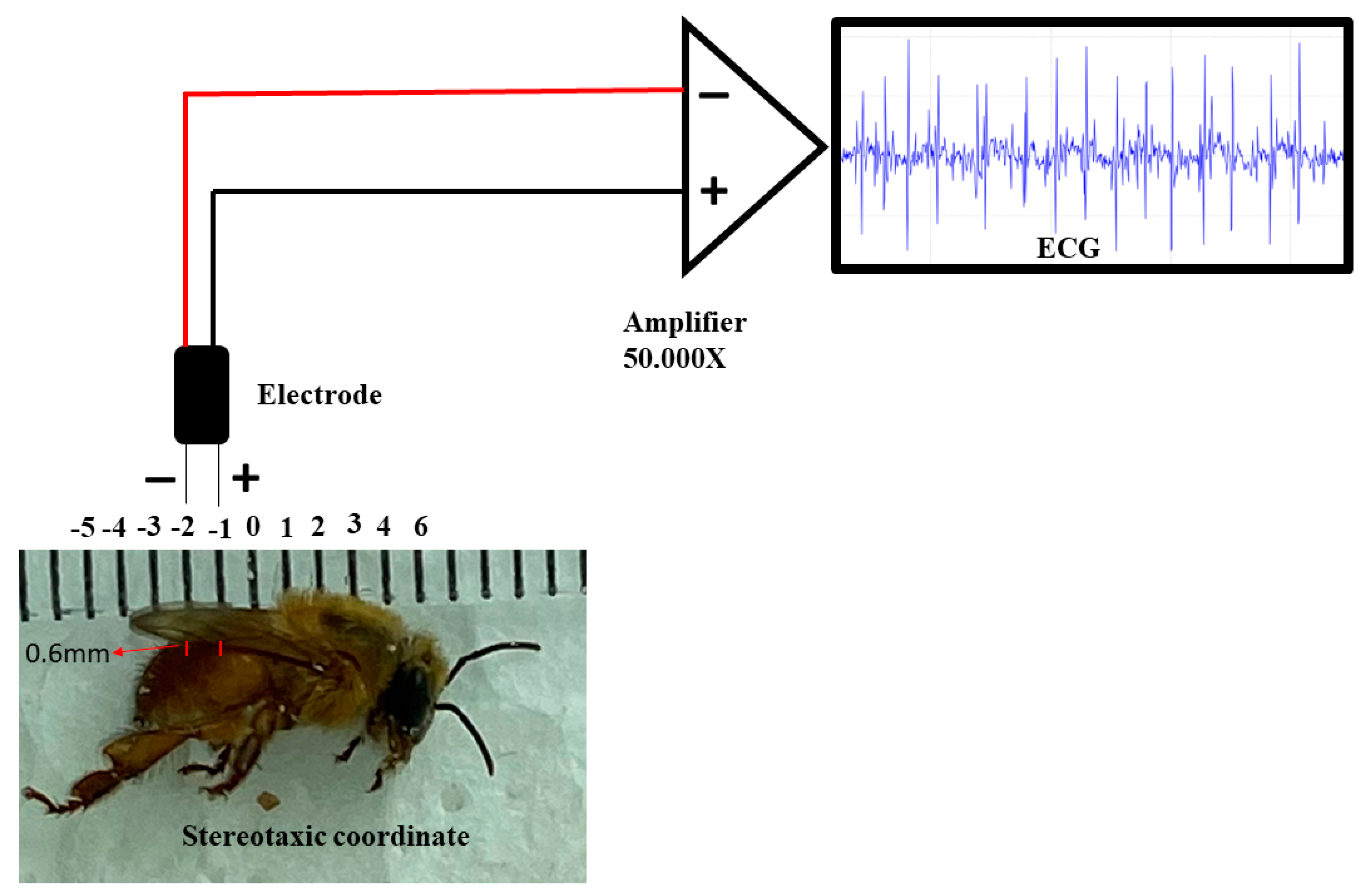
2.3. Manufacture of Electrodes, Implantation and Obtaining of Electrocardiographic (ECG) Recordings
2.4. Statistical Analyses
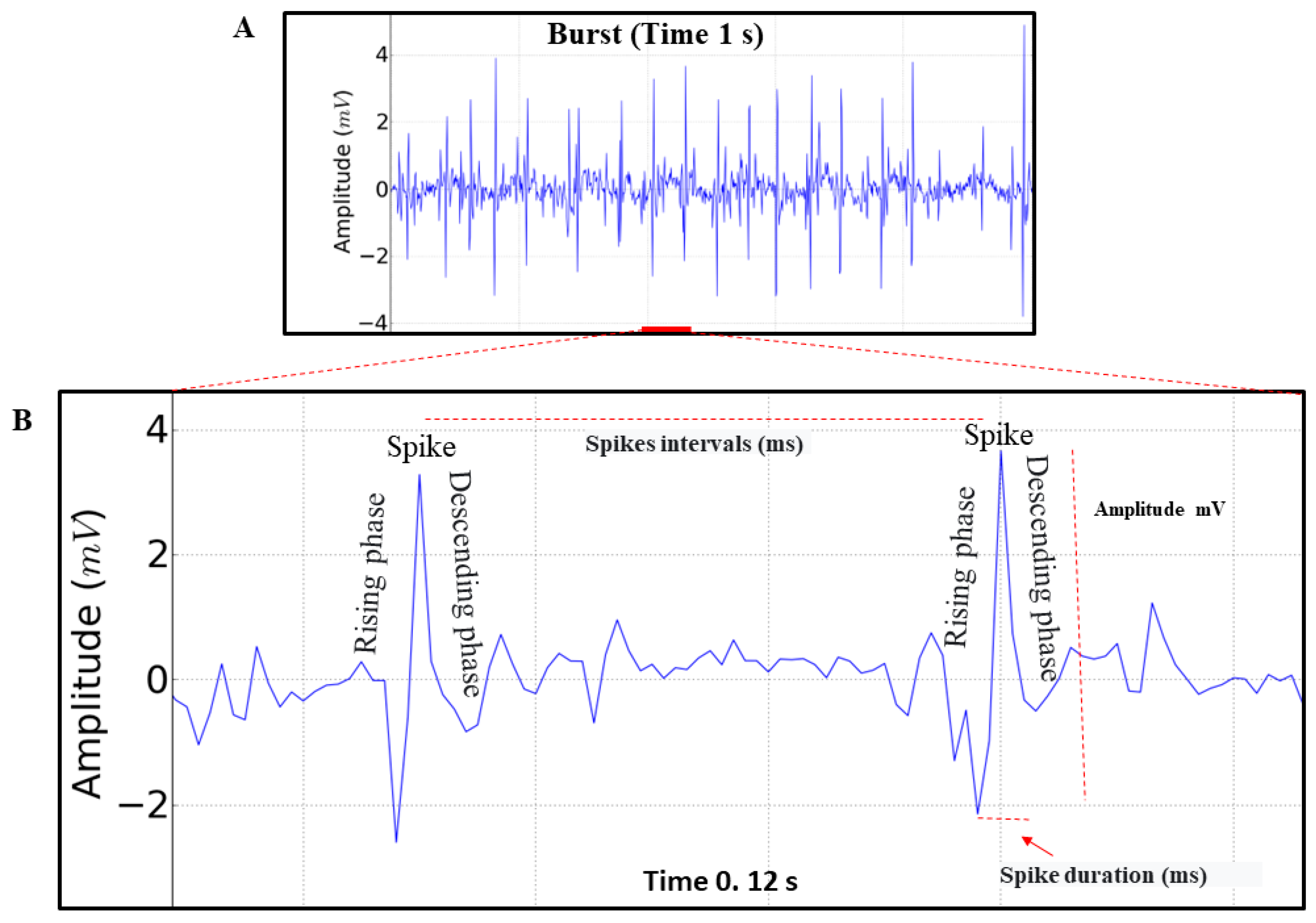
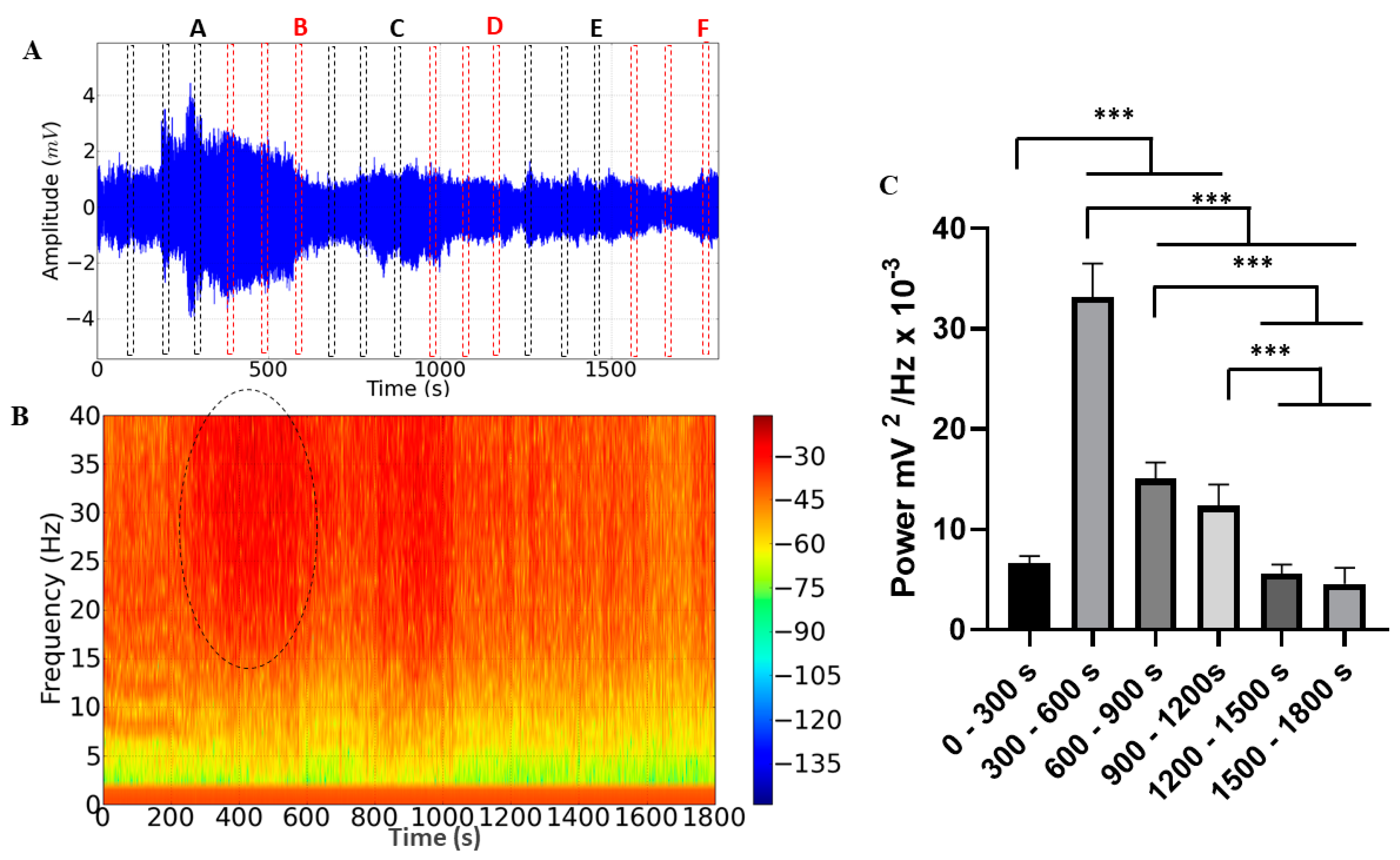
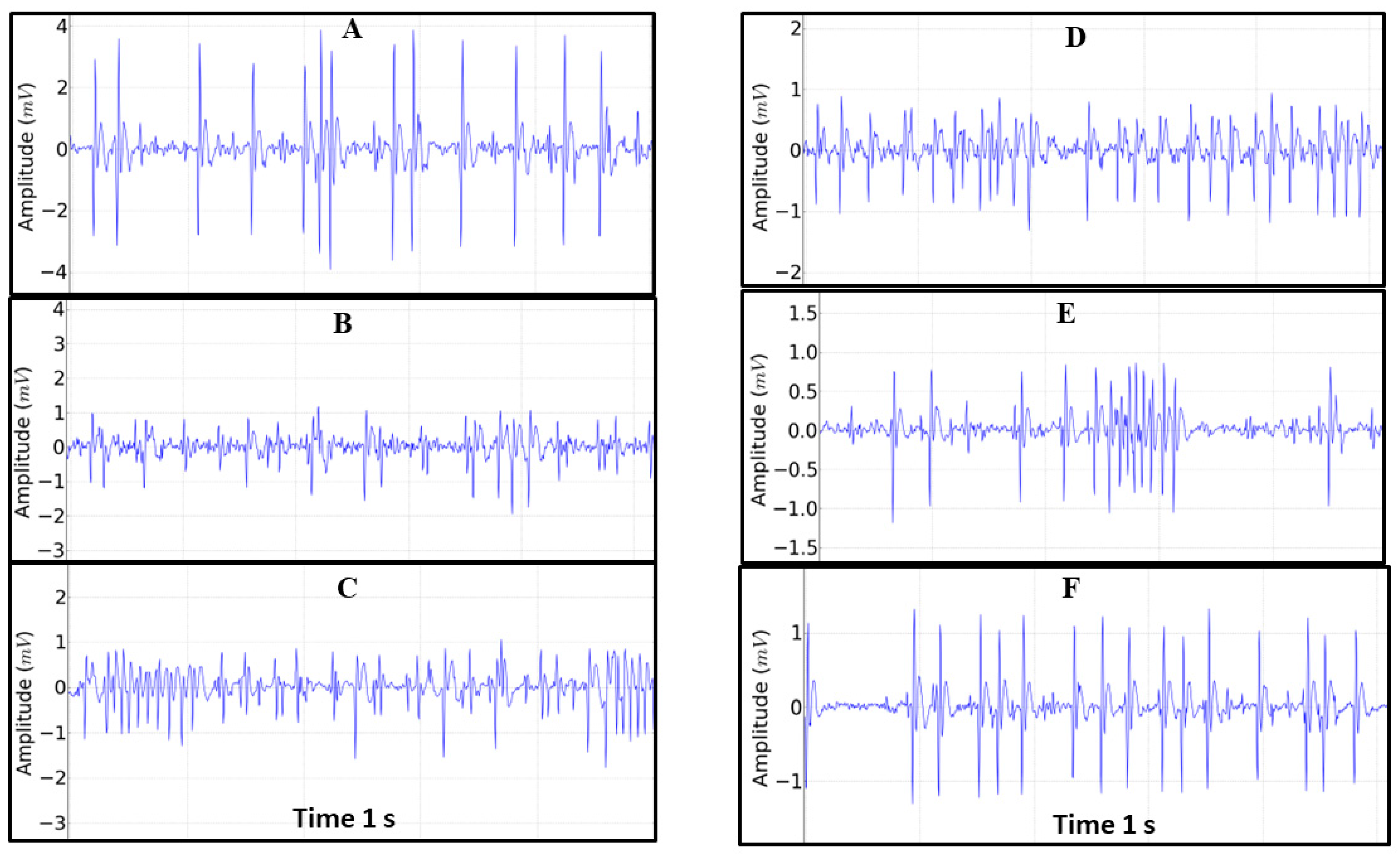
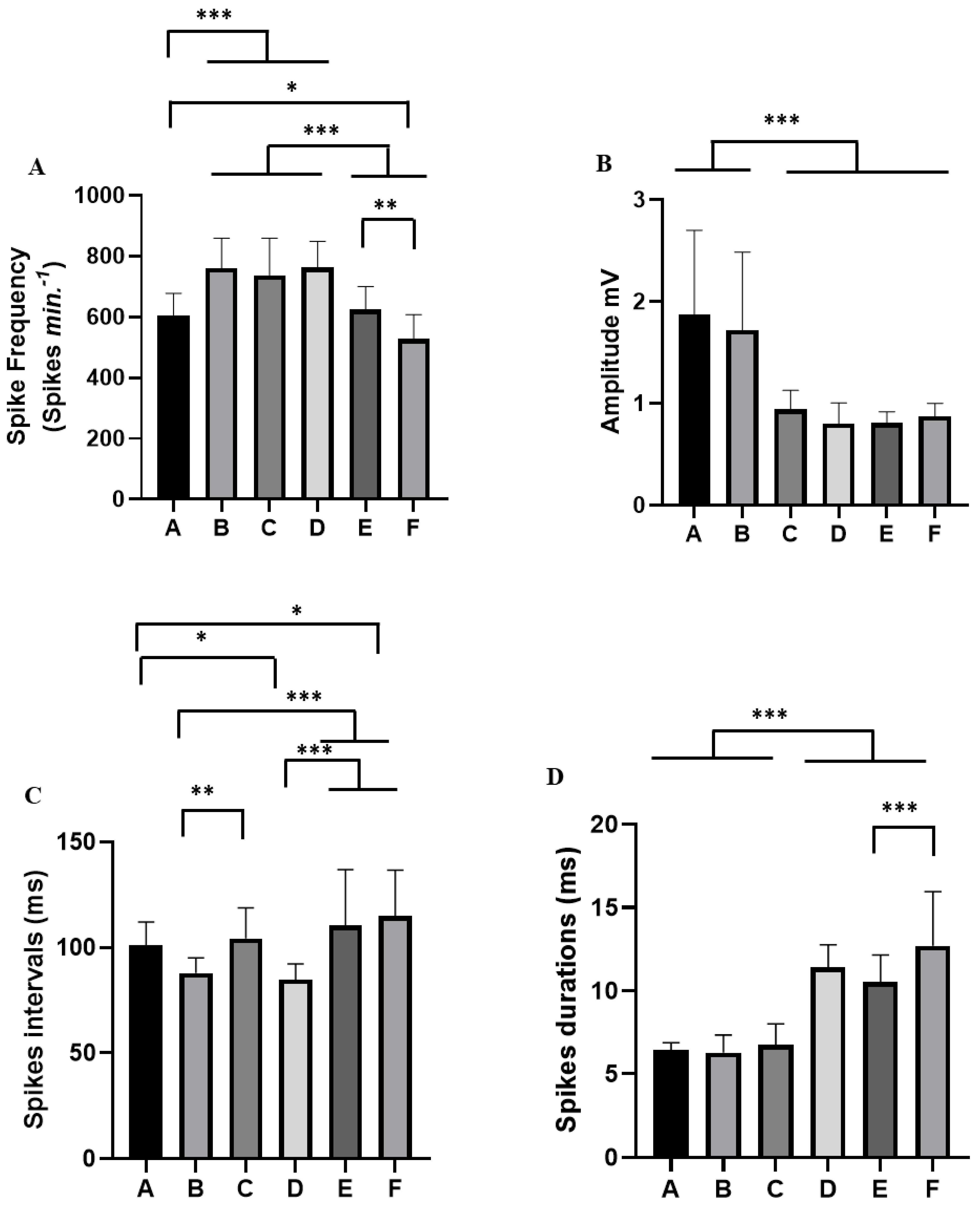
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bakker, R. Electrophysiology of fish intestine. In Transport Processes, Iono-and Osmoregulation. Proceedings in Life Sciences; Gilles, R., Gilles-Baillien, M., Eds.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 240–250. [Google Scholar] [CrossRef]
- Dubin, M.D. Interpretação Rápida do ECG, 3rd ed.; De Publicações Científicas: Rio de Janeiro, Brazil, 1996; 295p. [Google Scholar]
- Berkaya, S.K.; Uysal, A.K.; Gunal, E.S.; Ergin, S.; Gunal, S.; Gulmezoglu, M.B. A survey on ECG analysis. Biomed. Signal Process. Control 2018, 43, 216–235. [Google Scholar] [CrossRef]
- Hillyer, J.F. Insect heart rhythmicity is modulated by evolutionarily conserved neuropeptides and neurotransmitters. Curr. Opin. Insect Sci. 2018, 29, 41–48. [Google Scholar] [CrossRef]
- Minelli, A.; Boxshall, G.; Fusco, G. (Eds.) An Introduction to the Biology and Evolution of Arthropods. In Artrophod Biology and Evolution; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar] [CrossRef]
- Klowden, M.J. Physiological Systems in Insects; Academic Press: Cambridge, MA, USA, 2013; 682p. [Google Scholar]
- Borges, R.C.; Padovani, K.; Imperatriz-Fonseca, V.L.; Giannini, T.C. A Dataset of multi-functional ecological traits of Brazilian bees. Sci. Data 2020, 7, 120. [Google Scholar] [CrossRef]
- Ollerton, J. Pollinator Diversity: Distribution, Ecological Function, and Conservation. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 353–376. [Google Scholar] [CrossRef]
- Chapman, R.F. The Insects: Structure and Function, 4th ed.; Cambridge University Press: Cambridge, UK, 1998; 788p. [Google Scholar]
- Cruz-Landim, C. Abelhas—Morfologia e Função de Sistemas; UNESP: Sao Paulo, Brasil, 2009; 408p. [Google Scholar] [CrossRef]
- Carreck, N.L.; Andree, M.; Brent, C.S.; Cox-Foster, D.; Dade, H.A.; Ellis, J.D.; Hatjina, F.; Englesdorp, D.V. Standard methods for Apis mellifera anatomy and dissection. J. Apic. Res. 2013, 52, 1–40. [Google Scholar] [CrossRef]
- O’Neal, S.; Anderson, T. Dissection and Observation of Honey Bee Dorsal Vessel for Studies Cardiac Function. J. Vis. Exp. 2016, 118, 55029. [Google Scholar] [CrossRef]
- Cruz-Landim, C. Degenerative changes in the heart muscle from senescent honeybee workers (Apis mellifera adansonii). J. Invertebr. Pathol. 1976, 27, 1–5. [Google Scholar] [CrossRef]
- Kuusik, A.; Martin, A.-J.; Mänd, M.; Hiiesaar, K.; Metspalu, L.; Tartes, U. Interrelations of gas exchange cycles, body movements and heartbeats in the foragers of bumblebee Bombus terrestris (Hymenoptera: Apidae) al low temperatures. Eur. J. Entomol. 2002, 99, 209–214. [Google Scholar] [CrossRef]
- Mänd, M.; Kuusik, A.; Martin, A.-J.; Williams, I.H.; Luik, A.; Karise, R.; Metspalu, L.; Hiesaar, K. Regular periods of abdominal contractions recorded from larvae of the bumblebee, Bombus terrestris (Hymenoptera: Apidae). Eur. J. Entomol. 2006, 103, 319–322. [Google Scholar] [CrossRef]
- O’Neal, S.; Brewster, C.C.; Bloomquist, J.F.; Anderson, T. Amitraz and its metabolite modulate honey bee cardiac function and tolerance to viral infection. J. Invertebr. Pathol. 2017, 149, 119–126. [Google Scholar] [CrossRef]
- O’Neal, S.; Swale, D.R.; Bloomquist, J.F.; Anderson, T. ATP-sensitive inwardly rectifying potassium channel modulators alter cardiac function in honey bees. J. Insect Physiol. 2017, 99, 95–100. [Google Scholar] [CrossRef]
- Papaefthimiou, C.; Papachristoforou, A.; Theophilidis, G. Biphasic responses of the honeybee heart to nanomolar concentrations of amitraz. Pestic. Biochem. Physiol. 2013, 107, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Schwab, E.R.; Chilson, R.A.; Eddleman, C.D. Heartbeat rate modulation mediated by the ventral nerve cord in the honey bee, Apis mellifera. J. Comp. Physiol. B 1991, 161, 602–610. [Google Scholar] [CrossRef]
- Yelkovan, S.; Arıkan, H.; Çakıcı, Ö. Caste and age related changes in circulatory hemocytes of honey bee, Apis mellifera anatolica (Hymenoptera: Apidae). J. Apic. Res. 2021, 60, 512–521. [Google Scholar] [CrossRef]
- Michener, C.D. The Bees of the World, 2nd ed.; John Hopkins University Press: Baltimore, MD, USA, 2007; 953p. [Google Scholar]
- Roubik, D.W. Stingless Bee (Apidae: Apinae: Meliponini) Ecology. Annu. Rev. Entomol. 2023, 68, 231–256. [Google Scholar] [CrossRef]
- Gonzalez, V.H.; Cobos, M.E.; Jaramillo, J.; Ospina, R. Climate change will reduce the potential distribution ranges of Colombia’s most valuable pollinators. Perspect. Ecol. Evol. 2001, 19, 195–206. [Google Scholar] [CrossRef]
- Maia-Silva, C.; da Silva Pereira, J.; Freitas, B.M.; Hrncir, M. Don’t stay out too long! Thermal tolerance of the stingless bees Melipona subnitida decreases with increasing exposure time to elevated temperatures. Apidologie 2021, 52, 218–229. [Google Scholar] [CrossRef]
- Cham, K.O.; Nocelli, R.C.F.; Borges, L.O.; Viana-Silva, F.E.C.; Tonelli, C.A.M.; Malaspina, O.; Menezes, C.; Rosa-Fontana, A.S.; Blochtein, B.; Freitas, B.M.; et al. Pesticide Exposure Assessment Paradigm for Stingless Bees. Environ. Entomol. 2019, 48, 36–48. [Google Scholar] [CrossRef]
- Lourencetti, A.P.S.; Azevedo, P.; Miotelo, L.; Malaspina, O.; Nocelli, R.C.F. Surrogate species in pesticide risk assessments: Toxicological data of three stingless bees species. Environ. Pollut. 2023, 318, 120842. [Google Scholar] [CrossRef]
- Thompson, L.; Santos, G.; Cione, A. Letter to the editor regarding Lourencetti et al. Surrogate species in pesticide risk assessments: Toxicological data of three stingless bees species. Environ. Pollut. 2023, 319, 121011. [Google Scholar] [CrossRef]
- Pedro, S.R.M. The stingless bee fauna in Brazil (Hymenoptera: Apidae). Sociobiology 2014, 61, 348–354. [Google Scholar] [CrossRef]
- Pimentel, A.D.A.; de Abreu, V.H.R.; Krug, C.; Miranda, I.S. A review of pollen types foraged by Melipona in the Brazilian Amazon. Palynology 2023, 2175064. [Google Scholar] [CrossRef]
- Costa, L.; Venturieri, G.C. Diet impacts on Melipona flavolineata workers (Apidae, Meliponini). J. Apic. Res. 2009, 48, 38–45. [Google Scholar] [CrossRef]
- Teixeira, J.; Queiroz, A.C.; Veiga, J.; Leão, K.; Contrera, F.; Domingues, F.; Fontes, J.E.; Lopes, T.; Marsaioli, A.; Menezes, C. Soy extract as protein replacement to feed Melipona flavolineata Friese (Hymenoptera, Apidae, Meliponini). J. Apic. Res. 2020, 59, 104–114. [Google Scholar] [CrossRef]
- Veiga, J.C.; Leão, K.L.; Coelho, B.W.; Queiroz, A.C.M.; Menezes, C.; Contrera, F.A.L. The Life Histories of the “Uruçu Amarela” Males (Melipona flavolineata, Apidae, Meliponini). Sociobiology 2018, 65, 780–783. [Google Scholar] [CrossRef]
- Mcmillan, H.A.; Nørgard, M.; Maclean, H.J.; Overgaard, J.; Williams, C.J.A. A critical test of Drosophila anaesthetics: Isoflurane and sevoflurane are benign alternatives to cold and CO2. J. Insect Physiol. 2017, 101, 97–106. [Google Scholar] [CrossRef]
- Gooley, Z.C.; Gooley, A.C. Metabolic effects of anesthetics (cold, CO2, and isoflurane) and captivity conditions in isolated honey bee (Apis mellifera) foragers under different ambient temperatures. J. Apic. Res. 2021. [Google Scholar] [CrossRef]
- Ludin, N.M.; Cheeseman, J.F.; Merry, A.F.; Millar, C.D.; Warman, G.R. The effects of the general anaesthetic isoflurane on the honey bee (Apis mellifera) circadian clock. Chronobiol. Int. 2016, 33, 128–133. [Google Scholar] [CrossRef]
- Kaiser, W.; Weber, T.; Otto, D.; Miroschnikow, A. Oxygen supply of the heart and electrocardiogram potentials with reversed polarity in sleeping and resting honey bees. Apidologie 2014, 45, 73–87. [Google Scholar] [CrossRef]
- Lima, M.A.P.; Martins, G.F.; Oliveira, E.E.; Guedes, R.N.C. Agrochemical-induced stress in stingless bees: Peculiarities, underlying basis, and challenges. J. Comp. Physiol. A 2016, 202, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-M.; Fu, Y.; He, J.; Wang, J.-H. Effects of cold narcosis on memory acquisition, consolidation and retrieval in honeybees (Apis mellifera). Zool. Res. 2013, 35, 118–123. [Google Scholar] [CrossRef]
- Groening, J.; Venini, D.; Srinivasan, M.V. Effects of cold anaesthesia on the defensive behaviour of honeybees. Insectes Sociaux 2018, 65, 359–366. [Google Scholar] [CrossRef]
- Stec, D.; Kuszewska, K. CO2 narcosis influences the memory of honey bees. J. Apic. Res. 2020, 59, 663–668. [Google Scholar] [CrossRef]
- Tutun, H.; Sevin, S.; Çetintav, B. Effects of different chilling procedures on honey bees (Apis mellifera) for anestesia. Ank. Univ. Vet. Fak. Derg. 2020, 67, 289–294. [Google Scholar] [CrossRef]
- Gu, G.G.; Singh, S. Pharmacological analysis of heartbeat in Drosophila. J. Neurobiol. 1995, 28, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Wasserthal, L.T. Interaction of circulation and tracheal ventilation in holometabolous insects. Adv. Insect Physiol. 1996, 26, 297–351. [Google Scholar] [CrossRef]
- Hyllier, J.F.; Pass, G. The insect circulatory system: Structure, function, and evolution. Annu. Rev. Entomol. 2020, 65, 121–143. [Google Scholar] [CrossRef]
- Wasserthal, L.T. Drosophila flies combine periodic heartbeat reversal with a circulation in the anterior body mediated by a newly discovered anterior pair of ostial valves and ‘venous’ channels. J. Exp. Biol. 2007, 210, 3707–3719. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contrera, F.A.L.; Lopes, B.d.S.C.; Paz, C.A.d.; Hamoy, M.K.O.; Santos, M.F.d.; Barbosa, G.B.; Amaral, A.L.G.d.; Pinho, L.H.B.d.; Hamoy, M. First Records of Heartbeats via ECG in a Stingless Bee, Melipona flavolineata (Apidae, Meliponini), during Contention Stress Using Isoflurane as an Anesthetic. Insects 2023, 14, 696. https://doi.org/10.3390/insects14080696
Contrera FAL, Lopes BdSC, Paz CAd, Hamoy MKO, Santos MFd, Barbosa GB, Amaral ALGd, Pinho LHBd, Hamoy M. First Records of Heartbeats via ECG in a Stingless Bee, Melipona flavolineata (Apidae, Meliponini), during Contention Stress Using Isoflurane as an Anesthetic. Insects. 2023; 14(8):696. https://doi.org/10.3390/insects14080696
Chicago/Turabian StyleContrera, Felipe Andrés León, Bárbara dos Santos Conceição Lopes, Clarissa Araújo da Paz, Maria Klara Otake Hamoy, Murilo Farias dos Santos, Gabriela Brito Barbosa, Anthony Lucas Gurgel do Amaral, Luiz Henrique Barbosa de Pinho, and Moisés Hamoy. 2023. "First Records of Heartbeats via ECG in a Stingless Bee, Melipona flavolineata (Apidae, Meliponini), during Contention Stress Using Isoflurane as an Anesthetic" Insects 14, no. 8: 696. https://doi.org/10.3390/insects14080696
APA StyleContrera, F. A. L., Lopes, B. d. S. C., Paz, C. A. d., Hamoy, M. K. O., Santos, M. F. d., Barbosa, G. B., Amaral, A. L. G. d., Pinho, L. H. B. d., & Hamoy, M. (2023). First Records of Heartbeats via ECG in a Stingless Bee, Melipona flavolineata (Apidae, Meliponini), during Contention Stress Using Isoflurane as an Anesthetic. Insects, 14(8), 696. https://doi.org/10.3390/insects14080696