
Novel Lactone-Based Insecticides and Drosophila suzukii Management: Synthesis, Potential Action Mechanisms and Selectivity for Non-Target Parasitoids
 , ,
, ,  , ,
, ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Synthesis Process
2.2. Chemical Solutions Preparation
2.3. Toxicities on D. suzukii
2.4. Toxicities on the Parasitoid T. anastrephae
2.5. In Silico Evaluation of the Potential Target Receptors of Lactone Derivates on D. suzukii and T. anastrephae
2.5.1. Prediction of Putative Targets of Lactone Derivates
2.5.2. Data Resources for the Selected Target Receptors of D. suzukii and T. anastrephae
2.5.3. Generation and Validation of 3D Structures of Target Receptors
2.5.4. Molecular Docking of Lactone Derivates against Target Receptors
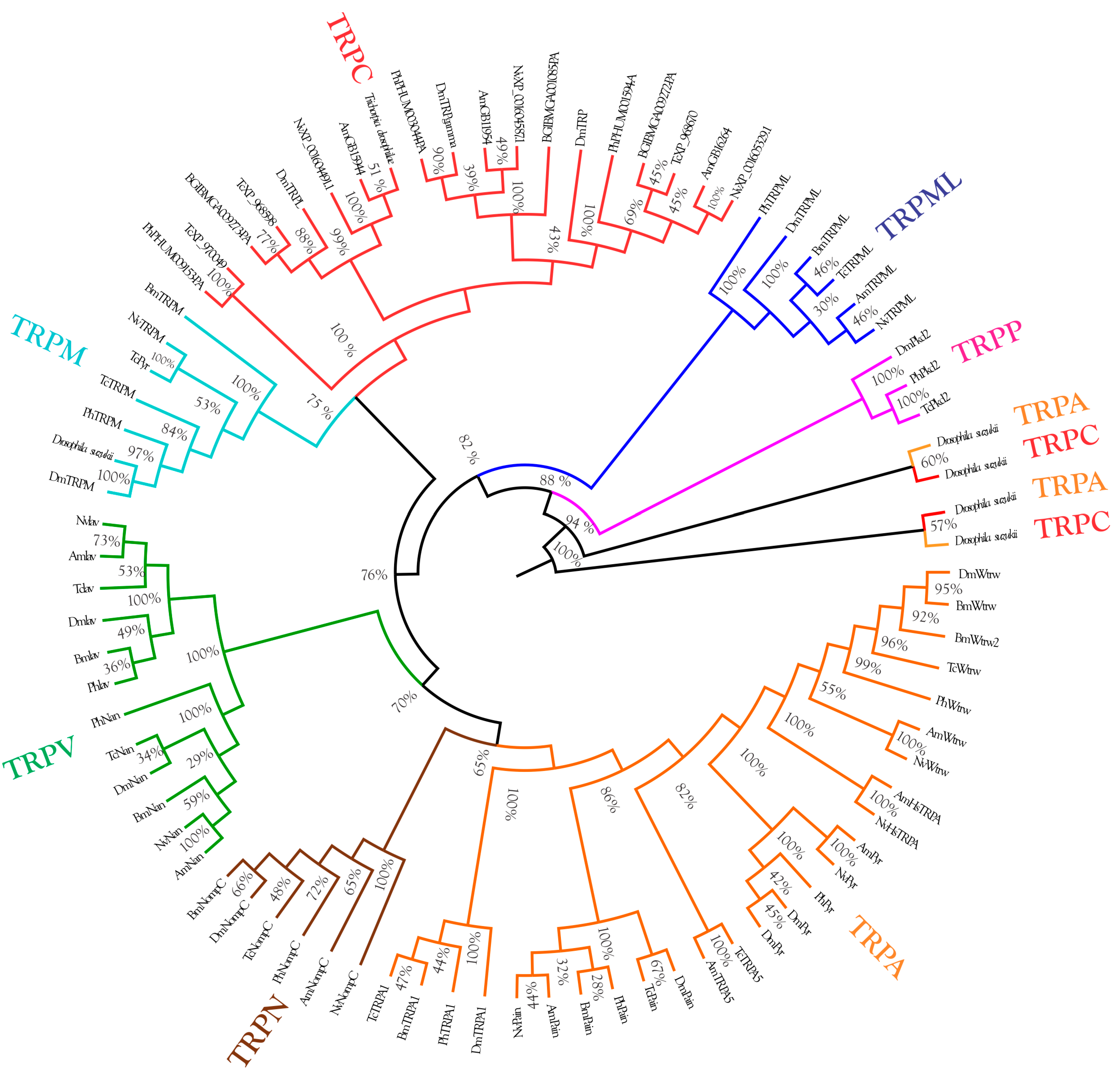
2.5.5. Phylogenetic Analysis of TRP Channels
3. Results
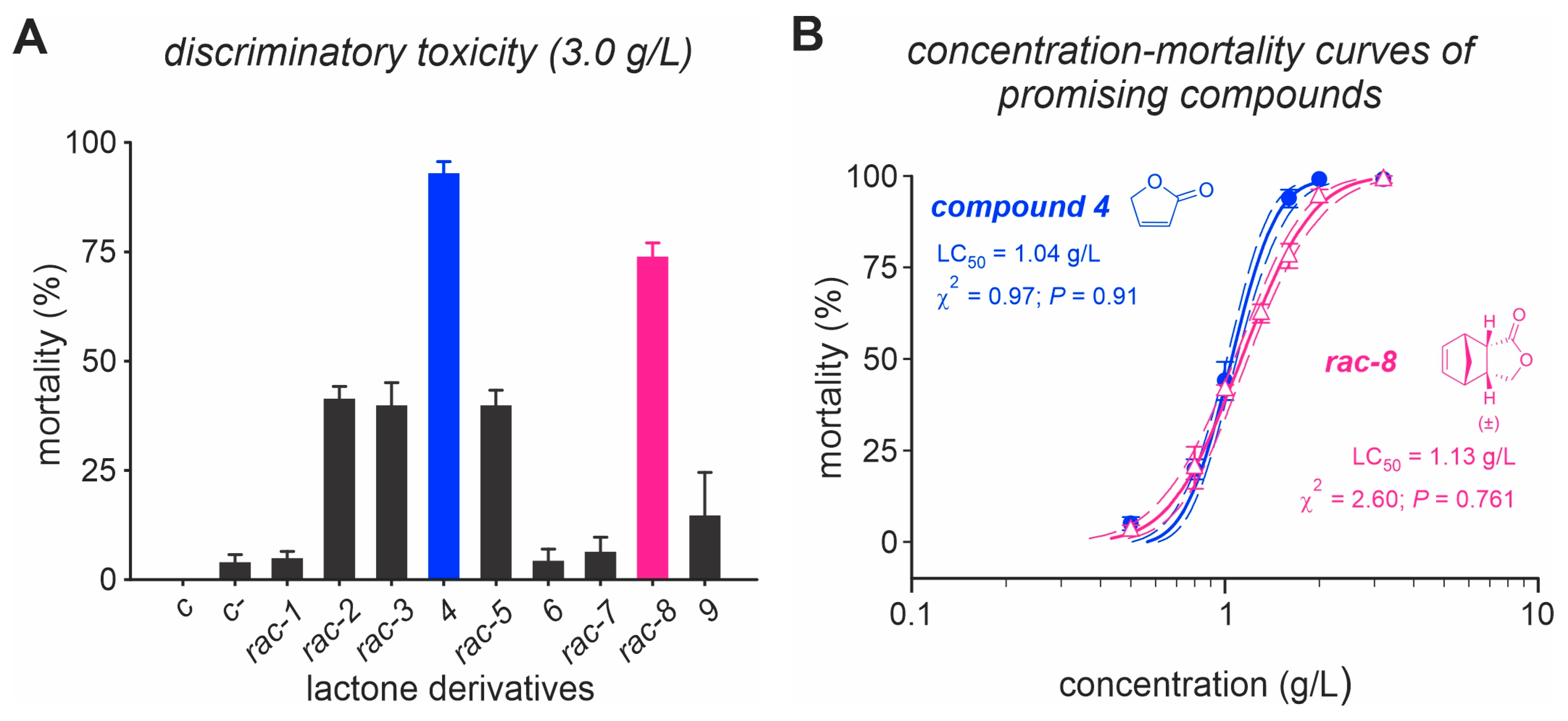
3.1. Insecticide Activity of Lactone Derivatives
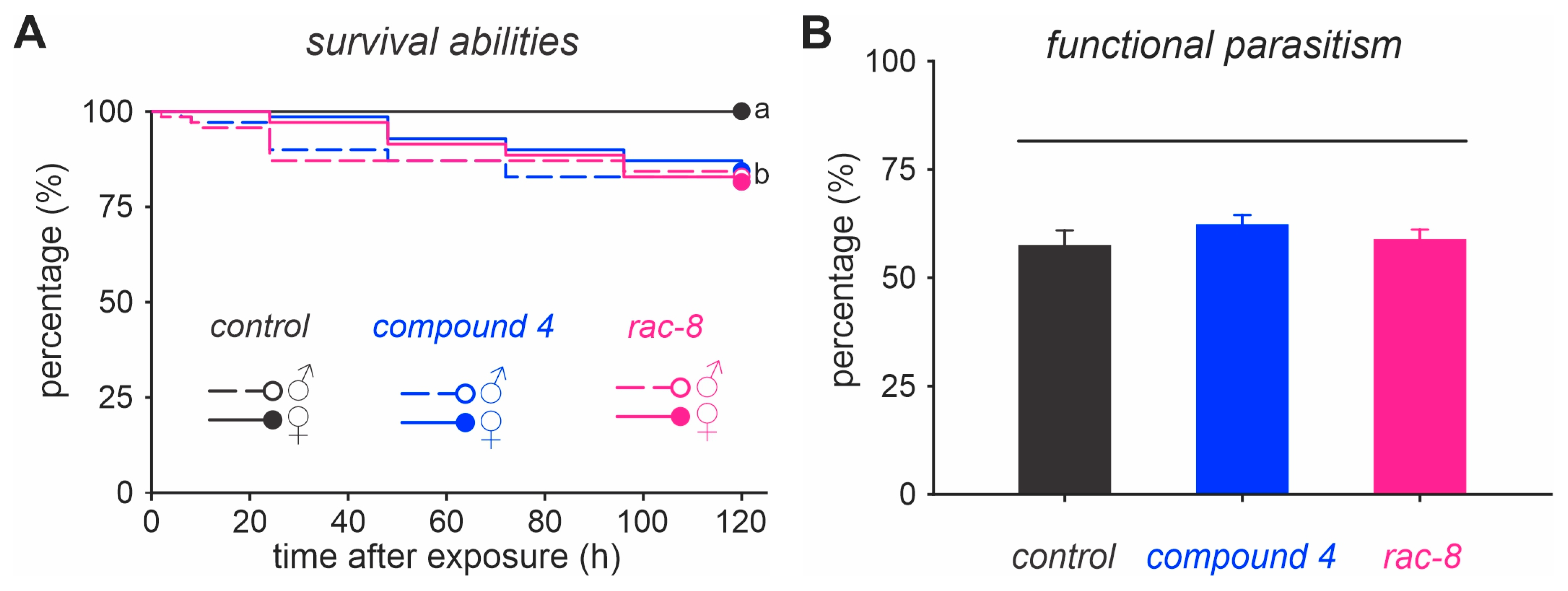
3.2. Functional Selectivity of Compound 4 and (rac)-8 Lactone Derivates to T. anastrephae Adults
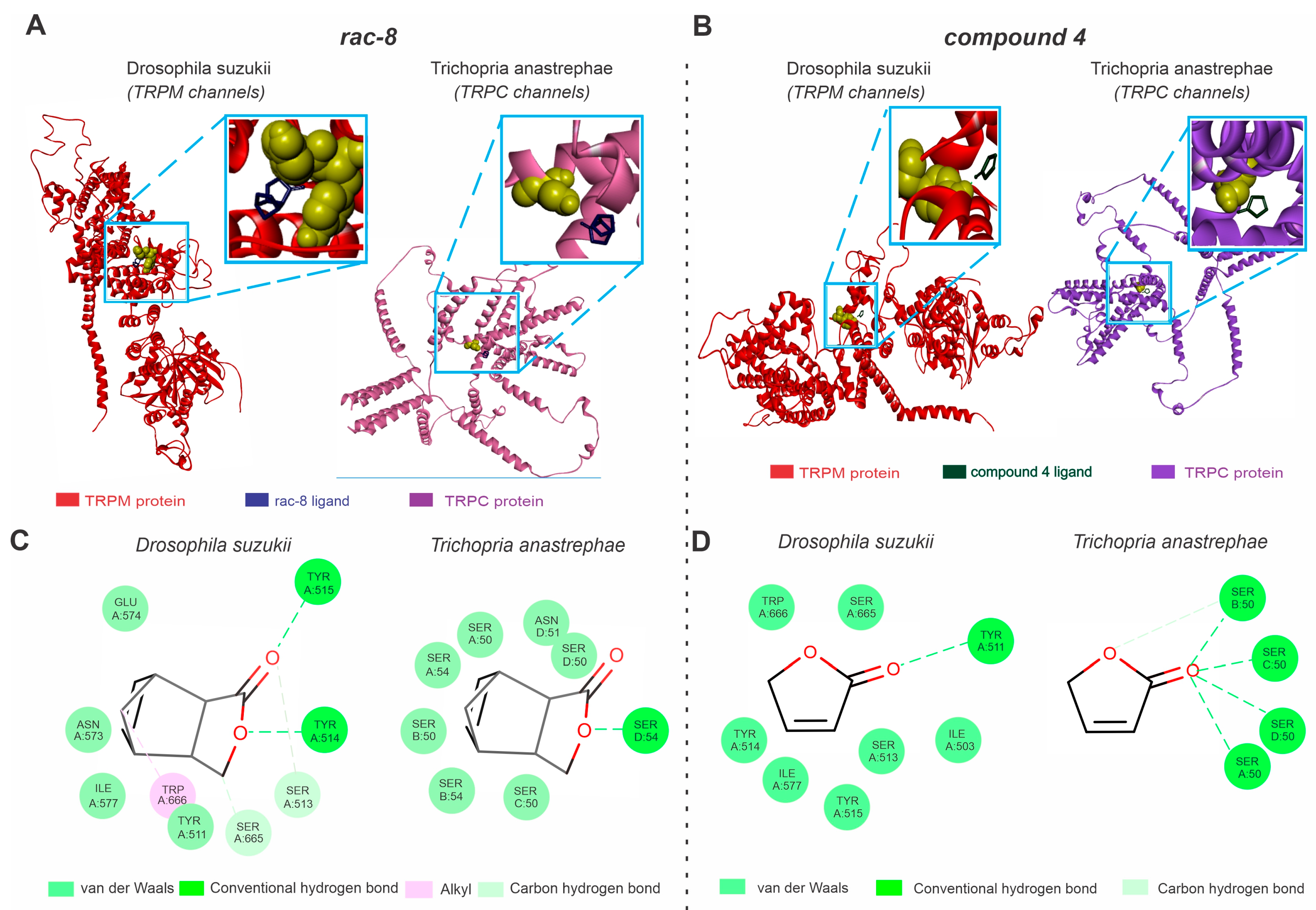
3.3. Molecular Docking Analysis of the TRP Channels with Lactone Derivatives
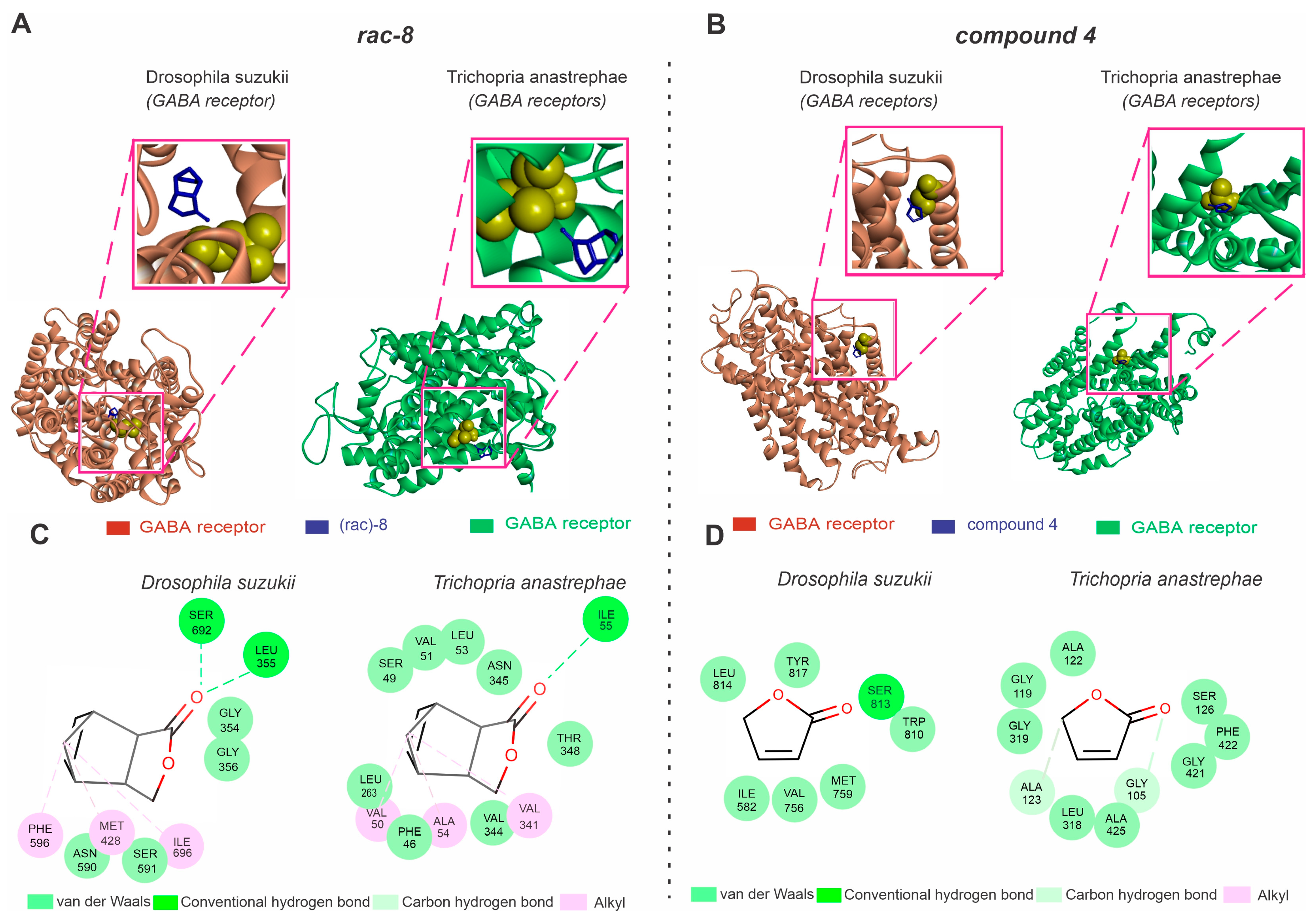
3.4. Molecular Docking Analysis of the GABA Receptors with Lactone Derivatives
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Andreazza, F.; Bernardi, D.; Dos Santos, R.; Garcia, F.; Oliveira, E.E.; Botton, M.; Nava, D.E. Drosophila Suzukii in Southern Neotropical Region: Current Status and Future Perspectives. Neotrop. Entomol. 2017, 46, 591–605. [Google Scholar] [CrossRef] [Green Version]
- Garcia, F.R.M.; Lasa, R.; Funes, C.F.; Buzzetti, K. Drosophila Suzukii Management in Latin America: Current Status and Perspectives. J. Econ. Entomol. 2022, 115, 1008–1023. [Google Scholar] [CrossRef] [PubMed]
- Bolda, M.P.; Goodhue, R.E.; Zalom, F.G. Spotted Wing Drosophila: Potential Economic Impact of a Newly Estab-lished Pest. Agric. Resour. Econ. Update 2010, 13, 5–8. [Google Scholar]
- Wollmann, J.; Schlesener, D.C.H.; Mendes, S.R.; Krüger, A.P.; Martins, L.N.; Bernardi, D.; Garcia, M.S.; Garcia, F.R.M. Infestation Index of Drosophila Suzukii (Diptera: Drosophilidae) in Small Fruit in Southern Brazil. Arq. Inst. Biol. 2020, 87, e0432018. [Google Scholar] [CrossRef]
- Diepenbrock, L.M.; Hardin, J.A.; Burrack, H.J. Season-Long Programs for Control of Drosophila Suzukii in South-eastern U.S. Blackberries. Crop Prot. 2017, 98, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Shaw, B.; Hemer, S.; Cannon, M.F.L.; Rogai, F.; Fountain, M.T. Insecticide Control of Drosophila Suzukii in Com-mercial Sweet Cherry Crops under Cladding. Insects 2019, 10, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlesener, D.C.H.; Wollmann, J.; Pazini, J.D.B.; Padilha, A.C.; Grützmacher, A.D.; Garcia, F.R.M. Insecticide Toxicity to Drosophila Suzukii (Diptera: Drosophilidae) Parasitoids: Trichopria Anastrephae (Hymenoptera: Diapriidae) and Pachycrepoideus Vindemmiae (Hymenoptera: Pteromalidae). J. Econ. Entomol. 2019, 112, 1197–1206. [Google Scholar] [CrossRef]
- Andreazza, F.; Bernardi, D.; Baronio, C.A.; Pasinato, J.; Nava, D.E.; Botton, M. Toxicities and Effects of Insecticidal Toxic Baits to Control Drosophila Suzukii and Zaprionus Indianus (Diptera: Drosophilidae). Pest. Manag. Sci. 2017, 73, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Noble, R.; Shaw, B.; Walker, A.; Whitfield, E.C.; Deakin, G.; Harris, A.; Dobrovin-Pennington, A.; Fountain, M.T. Control of Spotted Wing Drosophila (Drosophila Suzukii) in Sweet Cherry and Raspberry Using Bait Sprays. J. Pest Sci. 2023, 96, 623–633. [Google Scholar] [CrossRef]
- Babu, A.; Rodriguez-Saona, C.; Sial, A.A. Factors Influencing the Efficacy of Novel Attract-and-Kill (ACTTRA SWD) Formulations Against Drosophila Suzukii. J. Econ. Entomol. 2022, 115, 981–989. [Google Scholar] [CrossRef]
- Urbaneja-Bernat, P.; Holdcraft, R.; Hernández-Cumplido, J.; Rhodes, E.M.; Liburd, O.E.; Sial, A.A.; Mafra-Neto, A.; Rodriguez-Saona, C. Field, Semi-Field and Greenhouse Testing of HOOK SWD, a SPLAT-Based Attract-and-Kill Formulation to Manage Spotted-Wing Drosophila. J. Appl. Entomol. 2022, 146, 1230–1242. [Google Scholar] [CrossRef]
- Hogg, B.N.; Lee, J.C.; Rogers, M.A.; Worth, L.; Nieto, D.J.; Stahl, J.M.; Daane, K.M. Releases of the Parasitoid Pachycrepoideus Vindemmiae for Augmentative Biological Control of Spotted Wing Drosophila, Drosophila Suzukii. Biol. Control 2022, 168, 104865. [Google Scholar] [CrossRef]
- Seehausen, M.L.; Valenti, R.; Fontes, J.; Meier, M.; Marazzi, C.; Mazzi, D.; Kenis, M. Large-Arena Field Cage Releases of a Candidate Classical Biological Control Agent for Spotted Wing Drosophila Suggest Low Risk to Non-Target Species. J. Pest Sci. 2022, 95, 1057–1065. [Google Scholar] [CrossRef]
- Cruz, P.P.; Neutzling, A.S.; Garcia, F.R.M. Primeiro Registro de Trichopria Anastrephae, Parasitoide de Moscas-Das-Frutas, No Rio Grande Do Sul. Ciência Rural. 2011, 41, 1297–1299. [Google Scholar] [CrossRef] [Green Version]
- Garcia, F.R.M.; Ricalde, M.P. Augmentative Biological Control Using Parasitoids for Fruit Fly Management in Brazil. Insects 2013, 4, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Chabert, S.; Allemand, R.; Poyet, M.; Eslin, P.; Gibert, P. Ability of European Parasitoids (Hymenoptera) to Control a New Invasive Asiatic Pest, Drosophila Suzukii. Biol. Control 2012, 63, 40–47. [Google Scholar] [CrossRef]
- Lumaret, J.-P.; Errouissi, F.; Floate, K.; Rombke, J.; Wardhaugh, K. A Review on the Toxicity and Non-Target Effects of Macrocyclic Lactones in Terrestrial and Aquatic Environments. Curr. Pharm. Biotechnol. 2012, 13, 1004–1060. [Google Scholar] [CrossRef] [Green Version]
- Batiha, G.E.S.; Alqahtani, A.; Ilesanmi, O.B.; Saati, A.A.; El-Mleeh, A.; Hetta, H.F.; Beshbishy, A.M. Avermectin Derivatives, Pharmacokinetics, Therapeutic and Toxic Dosages, Mechanism of Action, and Their Biological Effects. Pharmaceuticals 2020, 13, 196. [Google Scholar] [CrossRef]
- Perrin, D.D.W.L.F. Armarego Purification of Laboratory Chemicals. In Recueil des Travaux Chimiques des Pays-Bas; John Wiley & Sons, Ltd.: Oxford, UK, 1988; Volume 107, p. 685. [Google Scholar]
- Näsman, J.H. 3-Methyl-2(5H)-Furanone. Org. Synth. 1990, 68, 162. [Google Scholar] [CrossRef]
- Nakao, T.; Banba, S.; Hirase, K. Comparison between the Modes of Action of Novel Meta-Diamide and Macrocyclic Lactone Insecticides on the RDL GABA Receptor. Pestic. Biochem. Physiol. 2015, 120, 101–108. [Google Scholar] [CrossRef]
- Hadiatullah, H.; Zhang, Y.; Samurkas, A.; Xie, Y.; Sundarraj, R.; Zuilhof, H.; Qiao, J.; Yuchi, Z. Recent Progress in the Structural Study of Ion Channels as Insecticide Targets. Insect Sci. 2022, 29, 1522–1551. [Google Scholar] [CrossRef]
- Chalivendra, S. Microbial Toxins in Insect and Nematode Pest Biocontrol. Int. J. Mol. Sci. 2021, 22, 7657. [Google Scholar] [CrossRef] [PubMed]
- Toledo, P.F.S.; Ferreira, T.P.; Bastos, I.M.A.S.; Rezende, S.M.; Viteri Jumbo, L.O.; Didonet, J.; Andrade, B.S.; Melo, T.S.; Smagghe, G.; Oliveira, E.E.; et al. Essential Oil from Negramina (Siparuna Guianensis) Plants Controls Aphids without Impairing Survival and Predatory Abilities of Non-Target Ladybeetles. Environ. Pollut. 2019, 255, 113153. [Google Scholar] [CrossRef] [PubMed]
- Toledo, P.F.S.; Viteri Jumbo, L.O.; Rezende, S.M.; Haddi, K.; Silva, B.A.; Mello, T.S.; Della Lucia, T.M.C.; Aguiar, R.W.S.; Smagghe, G.; Oliveira, E.E. Disentangling the Ecotoxicological Selectivity of Clove Essential Oil against Aphids and Non-Target Ladybeetles. Sci. Total Environ. 2020, 718, 137328. [Google Scholar] [CrossRef] [PubMed]
- Nesterov, A.; Spalthoff, C.; Kandasamy, R.; Katana, R.; Rankl, N.B.; Andrés, M.; Jähde, P.; Dorsch, J.A.; Stam, L.F.; Braun, F.J.; et al. TRP Channels in Insect Stretch Receptors as Insecticide Targets. Neuron 2015, 86, 665–671. [Google Scholar] [CrossRef] [Green Version]
- Salgado, V.L. Insect TRP Channels as Targets for Insecticides and Repellents. J. Pestic. Sci. 2017, 42, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.G.; Alvarenga, E.S.; Pimentel, M.F.; Picanço, M.C. Synthesis and Insecticidal Activity of Lactones Derived from Furan-2(5H)-One. J. Braz. Chem. Soc. 2015, 26, 2279–2289. [Google Scholar] [CrossRef]
- Andreazza, F.; Vacacela Ajila, H.E.; Haddi, K.; Colares, F.; Pallini, A.; Oliveira, E.E. Toxicity to and Egg-Laying Avoidance of Drosophila Suzukii (Diptera: Drosophilidae) Caused by an Old Alternative Inorganic Insecticide Preparation. Pest. Manag. Sci. 2018, 74, 861–867. [Google Scholar] [CrossRef]
- Bernardi, D.; Ribeiro, L.; Andreazza, F.; Neitzke, C.; Oliveira, E.E.; Botton, M.; Nava, D.E.; Vendramim, J.D. Potential Use of Annona by Products to Control Drosophila Suzukii and Toxicity to Its Parasitoid Trichopria Anastrephae. Ind. Crop. Prod. 2017, 110, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Keiser, M.J.; Roth, B.L.; Armbruster, B.N.; Ernsberger, P.; Irwin, J.J.; Shoichet, B.K. Relating Protein Pharmacology by Ligand Chemistry. Nat. Biotechnol. 2007, 25, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated Data and New Features for Efficient Prediction of Protein Targets of Small Molecules. Nucleic Acids Res. 2019, 47, W357–W3664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Ref-erence Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S. Blast2GO: A Comprehensive Suite for Functional Analysis in Plant Genomics. Int. J. Plant Genomics 2008, 2008, 619832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, G.N.; Sasisekharan, V. Conformation of Polypeptides and Proteins. Adv. Protein Chem. 1968, 23, 283–437. [Google Scholar] [CrossRef]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the Estimation of the Absolute Quality of Individual Protein Structure Models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A Programming Language for Software Integration and Development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar]
- Forli, S.; Huey, R.; Pique, M.E.; Sanner, M.F.; Goodsell, D.S.; Olson, A.J. Computational Protein-Ligand Docking and Virtual Drug Screening with the AutoDock Suite. Nat. Protoc. 2016, 11, 905–919. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization and Multithreading. J. Comput. Chem. 2010, 31, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrödinger, L.; DeLano, W. PyMOL | Pymol.Org. Available online: https://pymol.org/2/ (accessed on 26 April 2023).
- Biovia, D. Biovia Discovery Studio® 2016 Comprehensive Modeling and Simulations. Biovia Discov. Studio 2016, 1, 4. [Google Scholar]
- Matsuura, H.; Sokabe, T.; Kohno, K.; Tominaga, M.; Kadowaki, T. Evolutionary Conservation and Changes in Insect TRP Channels. BMC Evol. Biol. 2009, 9, 228. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Isman, M.B.; Rodriguez, E. Larval Growth Inhibitors from Species of Prthenium (Asteraceae). Phytochemistry 1983, 22, 2709–2713. [Google Scholar] [CrossRef]
- Szczepanik, M.; Gliszczyńska, A.; Hnatejko, M.; Zawitowska, B. Effects of Halolactones with Strong Feed-ing-Deterrent Activity on the Growth and Development of Larvae of the Lesser Mealworm, Alphitobius Diaperinus (Coleoptera: Tenebrionidae). Appl. Entomol. Zool. 2016, 51, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Sosa, A.; Diaz, M.; Salvatore, A.; Bardon, A.; Borkosky, S.; Vera, N. Insecticidal Effects of Vernonanthura Nebularum against Two Economically Important Pest Insects. Saudi J. Biol. Sci. 2019, 26, 881–889. [Google Scholar] [CrossRef]
- Andreazza, F.; Haddi, K.; Oliveira, E.E.; Ferreira, J.A.M. Drosophila Suzukii (Diptera: Drosophilidae) Arrives at Minas Gerais State, a Main Strawberry Production Region in Brazil. Fla. Entomol. 2016, 99, 796–798. [Google Scholar] [CrossRef] [Green Version]
- Mendonca, L.D.P.; Oliveira, E.E.; Andreazza, F.; Rezende, S.M.; Faroni, L.R.D.A.; Guedes, R.N.C.; Haddi, K. Host Potential and Adaptive Responses of Drosophila Suzukii (Diptera: Drosophilidae) to Barbados Cherries. J. Econ. Entomol. 2019, 112, 3002–3006. [Google Scholar] [CrossRef]
- Ferronato, P.; Woch, A.L.; Soares, P.L.; Bernardi, D.; Botton, M.; Andreazza, F.; Oliveira, E.E.; Corrêa, A.S. A Phy-logeographic Approach to the Drosophila Suzukii (Diptera: Drosophilidae) Invasion in Brazil. J. Econ. Entomol. 2019, 112, 425–433. [Google Scholar] [CrossRef]
- Zanuncio-Junior, J.S.; Fornazier, M.J.; Andreazza, F.; Culik, M.P.; Mendonça, L.D.P.; Oliveira, E.E.; Martins, D.D.S.; Fornazier, M.L.; Costa, H.; Ventura, J.A. Spread of Two Invasive Flies (Diptera: Drosophilidae) Infesting Commercial Fruits in Southeastern Brazil. Fla. Entomol. 2018, 101, 522–525. [Google Scholar] [CrossRef] [Green Version]
- De Souza, M.T.; de Souza, M.T.; Morais, M.C.; Oliveira, D.d.C.; de Melo, D.J.; Figueiredo, L.; Zarbin, P.H.G.; Zawadneak, M.A.C.; Bernardi, D. Essential Oils as a Source of Ecofriendly Insecticides for Drosophila Suzukii (Diptera: Drosophilidae) and Their Potential Non-Target Effects. Molecules 2022, 27, 6215. [Google Scholar] [CrossRef] [PubMed]
- Haddi, K.; Turchen, L.M.; Viteri Jumbo, L.O.; Guedes, R.N.C.; Pereira, E.J.G.; Aguiar, R.W.S.; Oliveira, E.E. Re-thinking Biorational Insecticides for Pest Management: Unintended Effects and Consequences. Pest. Manag. Sci. 2020, 76, 2286–2293. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.G.A.; Krüger, A.P.; Scheuneumann, T.; Morais, M.C.; Speriogin, H.J.; Garcia, F.R.M.; Nava, D.E.; Bernardi, D. Some Aspects of the Biology of Trichopria Anastrephae (Hymenoptera: Diapriidae), a Resident Parasitoid At-tacking Drosophila Suzukii (Diptera: Drosophilidae) in Brazil. J. Econ. Entomol. 2020, 113, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Woltering, S.B.; Romeis, J.; Collatz, J. Influence of the Rearing Host on Biological Parameters of Trichopria Dro-sophilae, a Potential Biological Control Agent of Drosophila Suzukii. Insects 2019, 10, 183. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, T.; Chubanov, V.; Chen, X.; Dietz, A.S.; Gudermann, T.; Montell, C. Drosophila TRPM Channel Is Es-sential for the Control of Extracellular Magnesium Levels. PLoS ONE 2010, 5, e10519. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, K.; Oikawa, H.; Hasobe, M.; Suzuki, N.; Yajima, S.; Tomizawa, M. Contact Repellency by L-Menthol Is Mediated by TRPM Channels in the Red Flour Beetle Tribolium Castaneum. Pest. Manag. Sci. 2021, 77, 1422–1427. [Google Scholar] [CrossRef]
- Turner, H.N.; Armengol, K.; Patel, A.A.; Himmel, N.J.; Sullivan, L.; Iyer, S.C.; Bhattacharya, S.; Iyer, E.P.R.; Landry, C.; Galko, M.J.; et al. The TRP Channels Pkd2, NompC, and Trpm Act in Cold-Sensing Neurons to Mediate Unique Aversive Behaviors to Noxious Cold in Drosophila. Curr. Biol. 2016, 26, 3116–3128. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantilla Afanador, J.G.; Araujo, S.H.C.; Teixeira, M.G.; Lopes, D.T.; Cerceau, C.I.; Andreazza, F.; Oliveira, D.C.; Bernardi, D.; Moura, W.S.; Aguiar, R.W.S.; et al. Novel Lactone-Based Insecticides and Drosophila suzukii Management: Synthesis, Potential Action Mechanisms and Selectivity for Non-Target Parasitoids. Insects 2023, 14, 697. https://doi.org/10.3390/insects14080697
Mantilla Afanador JG, Araujo SHC, Teixeira MG, Lopes DT, Cerceau CI, Andreazza F, Oliveira DC, Bernardi D, Moura WS, Aguiar RWS, et al. Novel Lactone-Based Insecticides and Drosophila suzukii Management: Synthesis, Potential Action Mechanisms and Selectivity for Non-Target Parasitoids. Insects. 2023; 14(8):697. https://doi.org/10.3390/insects14080697
Chicago/Turabian StyleMantilla Afanador, Javier G., Sabrina H. C. Araujo, Milena G. Teixeira, Dayane T. Lopes, Cristiane I. Cerceau, Felipe Andreazza, Daiana C. Oliveira, Daniel Bernardi, Wellington S. Moura, Raimundo W. S. Aguiar, and et al. 2023. "Novel Lactone-Based Insecticides and Drosophila suzukii Management: Synthesis, Potential Action Mechanisms and Selectivity for Non-Target Parasitoids" Insects 14, no. 8: 697. https://doi.org/10.3390/insects14080697
APA StyleMantilla Afanador, J. G., Araujo, S. H. C., Teixeira, M. G., Lopes, D. T., Cerceau, C. I., Andreazza, F., Oliveira, D. C., Bernardi, D., Moura, W. S., Aguiar, R. W. S., Oliveira, A. C. S. S., Santos, G. R., Alvarenga, E. S., & Oliveira, E. E. (2023). Novel Lactone-Based Insecticides and Drosophila suzukii Management: Synthesis, Potential Action Mechanisms and Selectivity for Non-Target Parasitoids. Insects, 14(8), 697. https://doi.org/10.3390/insects14080697