
Monarch Butterflies in Western North America: A Holistic Review of Population Trends, Ecology, Stressors, Resilience and Adaptation


Abstract
:Simple Summary
Abstract
1. Introduction
2. Monarch Population Dynamics in Western North America (1980s–2022)
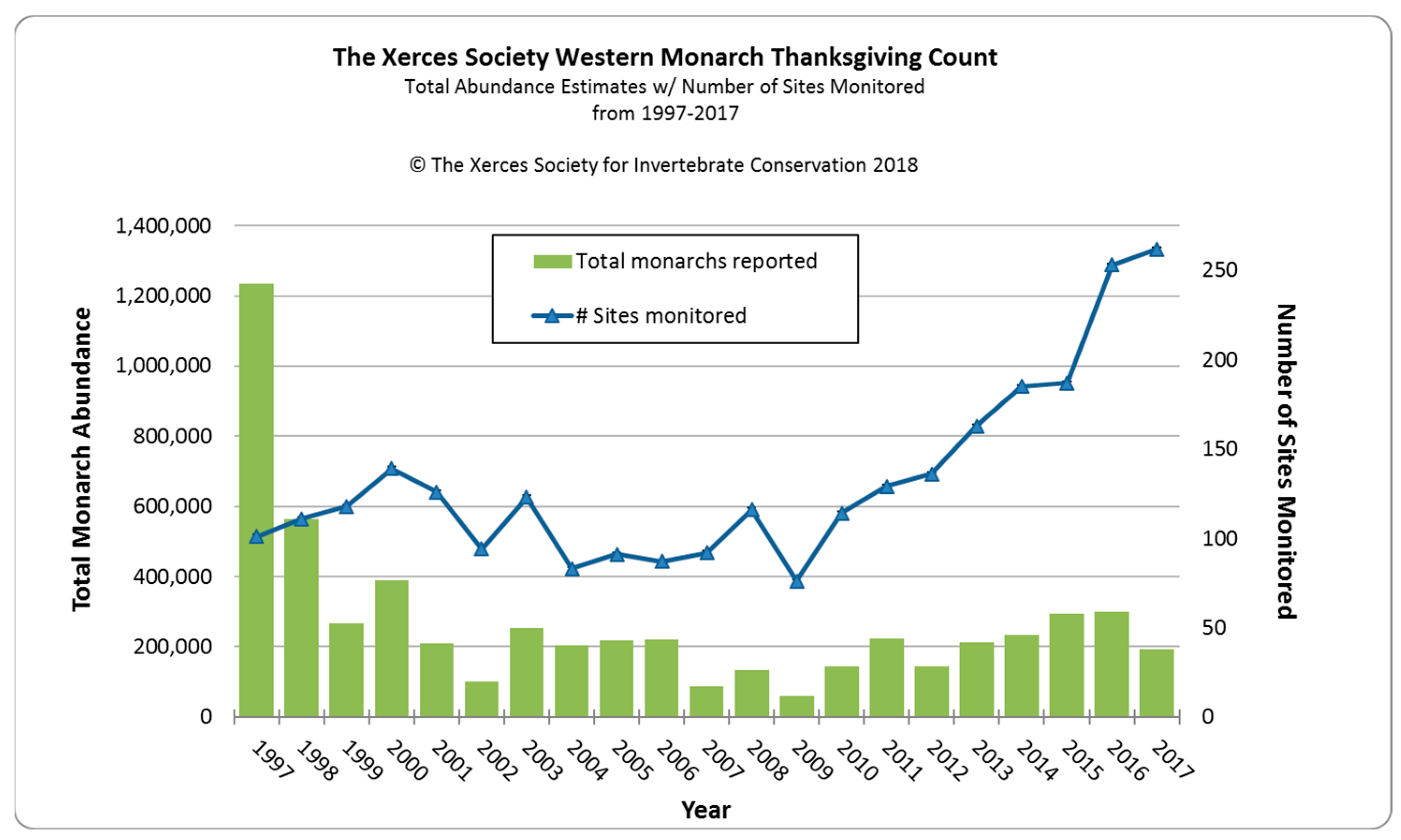
2.1. Monitoring
2.2. The Big Decline Post-1997
2.3. Neonicotinoids as the Potential Primary Driver of the Western Monarch Decline Post-1997
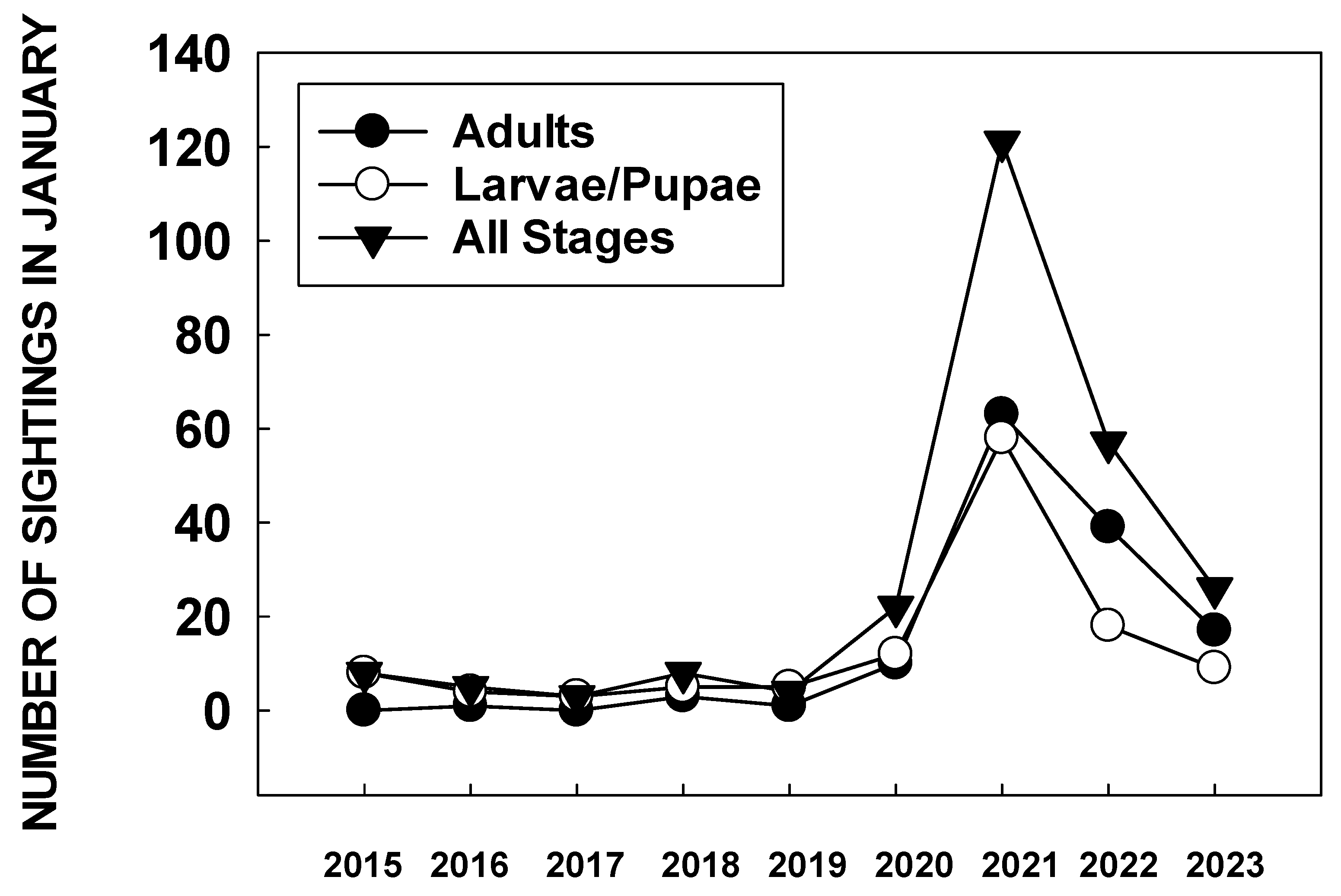
2.4. Sudden Decline 2018–2019
2.5. 2020: The Western Monarch Nadir
2.6. 2021: Monarch Resurrection
2.7. Western Monarchs: The Future
3. A Holistic View of Western Monarch Population Stressors
3.1. Pesticides
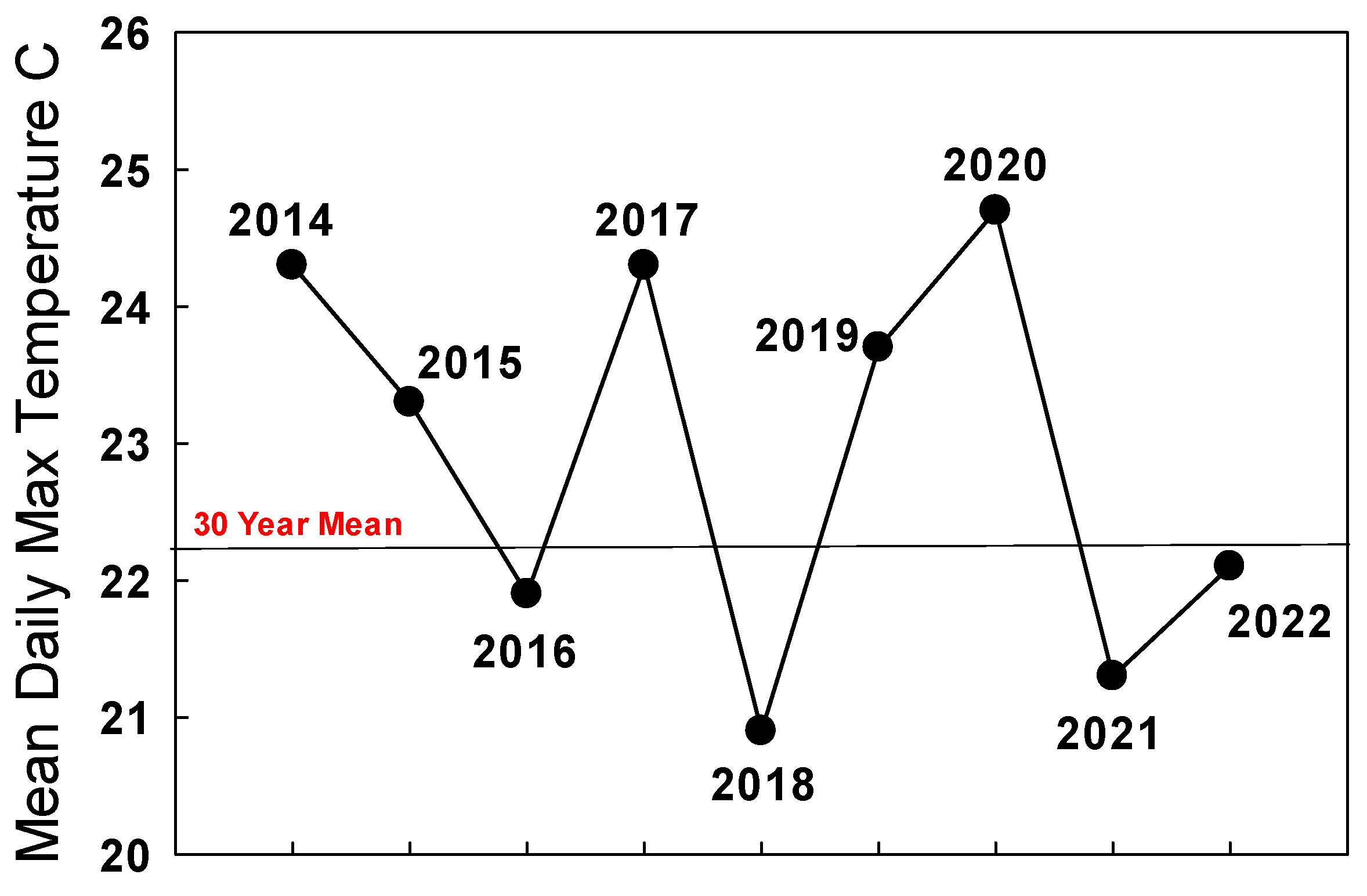
3.2. Climate
3.3. Habitat
3.3.1. Spring Habitat
3.3.2. Summer Habitat
3.3.3. Autumn Habitat
3.3.4. Winter Habitat
3.4. Natural Enemies
3.4.1. Predators
3.4.2. Parasitoids
3.4.3. Viral, Bacterial, and Unidentified Pathogens
3.4.4. Ophryocystis elektroscirrha (OE)
3.5. Non-Native Milkweeds
3.6. Human Interference
The Value of Rearing Monarchs
4. Conclusions and Perspectives
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schultz, C.B.; Brown, L.M.; Pelton, E.; Crone, E.E. Citizen science monitoring demonstrates dramatic declines of monarch butterflies in western North America. Biol. Conserv. 2017, 214, 343–346. [Google Scholar] [CrossRef]
- Pelton, E.M.; Schultz, C.B.; Jepsen, S.J.; Hoffman Black, S.; Crone, E.E. Western monarch population plummets: Status, probable causes and recommended conservation actions. Front. Ecol. Evol. 2019, 7, 258. [Google Scholar] [CrossRef]
- Crone, E.E.; Schultz, C.B. Resilience or catastrophe? A possible state change for monarch butterflies in western North America. Ecol. Lett. 2021, 24, 1533–1538. [Google Scholar] [CrossRef]
- Brower, L.P. Understanding and misunderstanding the migration of the monarch butterfly (Nymphalidae) in North America. J. Lepid. Soc. 1995, 49, 304–385. [Google Scholar]
- Thogmartin, W.E.; Widerholt, R.; Oberhauser, K.; Drum, R.G.; Diffendorfer, J.E.; Altizer, S.; Taylor, O.R.; Pleasants, J.; Semmens, D.; Semmens, B.; et al. Monarch butterfly decline: Identifying the threatening processes. Roy. Soc. Open Sci. 2017, 4, 170760. [Google Scholar] [CrossRef]
- Malcolm, S.B. Anthropogenic impacts on mortality and population viability of the monarch butterfly. Ann. Rev. Entomol. 2018, 63, 277–302. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, A.A.E.; Flockhart, D.T.T.; Newman, A.A.M.; Norris, D.R. An evaluation of studies on the potential threats contributing to the decline of eastern migratory North American monarch butterflies (Danaus plexippus). Front. Ecol. Evol. 2019, 7, 99. [Google Scholar] [CrossRef]
- Leong, K.L.H.; Sakai, W.H.; Bremer, W.; Feurstein, D.; Yoshimura, G. Analysis of the pattern of distribution and abundance of overwintering sites along the California coastline. In The Monarch Butterfly: Biology and Conservation; Oberhauser, K.S., Solensky, M.J., Eds.; Cornell University Press: Ithaca, NY, USA, 2004; pp. 177–185. [Google Scholar]
- Fisher, A.; Saniee, K.; van der Heide, C.; Griffiths, J.; Meade, D.; Villablanca, F. Climatic niche model for overwintering monarch butterflies in a topographically complex region of California. Insects 2018, 9, 167. [Google Scholar] [CrossRef]
- Tuskes, P.M.; Brower, L.P. Overwintering ecology of the monarch butterfly, Danaus plexippus L., in California. Ecol. Entomol. 1978, 3, 141–153. [Google Scholar] [CrossRef]
- Chaplin, S.B.; Wells, P.H. Energy reserves and metabolic expenditures of monarch butterflies overwintering in southern California. Ecol. Entomol. 1982, 7, 249–256. [Google Scholar] [CrossRef]
- Oberhauser, K.; Ries, L.; Altizer, S.; Batalden, R.V.; Kudell-Ekstrum, J.; Garland, M.; Howard, E.; Jepsen, S.; Lovett, J.; Monroe, M.; et al. Contributions to monarch biology and conservation through citizen science: Seventy years and counting. In Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly; Oberhauser, K.S., Nail, K.R., Altizer, S., Eds.; Cornell University Press: Ithaca, NY, USA, 2015; pp. 13–29. [Google Scholar]
- Williams, E.H.; Brower, L.P. A conservation concern: How many monarchs are there? News Lepid. Soc. 2016, 58, 90–93. [Google Scholar]
- Brower, L.P.; Taylor, O.R.; Williams, E.H.; Slayback, D.A.; Zubieta, R.R.; Ramirez, I. Decline of monarch butterflies overwintering in Mexico: Is the migratory phenomenon at risk? Insect Cons. Divers. 2012, 5, 95–100. [Google Scholar] [CrossRef]
- Pleasants, J.M.; Oberhauser, K. Milkweed loss in agricultural fields because of herbicide use: Effect on the monarch butterfly population. Insect Cons. Divers. 2013, 6, 135–144. [Google Scholar] [CrossRef]
- Yu, Z.; Lu, C. Historical cropland expansion and abandonment in the continental US during 1850 to 2016. Glob. Ecol. Biogeogr. 2017, 27, 322–333. [Google Scholar] [CrossRef]
- Flockhart, D.T.T.; Pichancourt, J.; Norris, D.R.; Martin, T.G. Unravelling the annual cycle in a migratory animal: Breeding season habitat loss drives population declines of monarch butterflies. J. Anim. Ecol. 2015, 84, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P.; Nauen, R. Neonicotinoids-from zero to hero in insecticide chemistry. Pest Sci. Man. 2008, 64, 1084–1098. [Google Scholar] [CrossRef] [PubMed]
- Douglas, M.R.; Tooker, J.F. Large-scale deployment of seed treatments has driven rapid increase in use of neonicotinoid insecticides and preemptive pest management in US field crops. Environ. Sci. Technol. 2015, 49, 5088–5097. [Google Scholar] [CrossRef]
- Sparks, T.C.; Crossthwaite, A.J.; Nauen, R.; Banba, S.; Codova, D.; Earley, F.; Ebbinghaus-Kintscher, U.; Fujioka, S.; Hirao, A.; Karmon, D.; et al. Insecticides, biologics and nematicides: Updates to IRAC’s mode of action classification-a tool for resistance management. Pest. Biochem. Phys. 2020, 167, 104587. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Blacquiere, T.; Field, L.M.; Hails, R.S.; Petrokofsky, G.; Potts, S.G.; Raine, N.E.; Vanbergen, A.J.; McLean, A.R. A restatement of the natural science evidence base concerning neonicotinoid insecticides and insect pollinators. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140558. [Google Scholar] [CrossRef]
- Lu, C.; Hung, Y.T.; Cheng, Q. A review of sub-lethal neonicotinoid insecticides exposure and effects on pollinators. Curr. Pollut. Rep. 2020, 6, 137–151. [Google Scholar] [CrossRef]
- Singla, A.; Barmota, H.; Sahoo, K.J.; Kang, B.K. Influence of neonicotinoids on pollinators: A review. J. Apic. Res. 2021, 60, 19–32. [Google Scholar] [CrossRef]
- Gilburn, A.S.; Bunnefield, N.; Wilson, J.M.; Botham, M.S.; Brereton, T.M.; Fox, R.; Goulson, D. Are neonicotinoid insecticides driving declines of widespread butterflies? Peer J. 2015, 3, e1402. [Google Scholar] [CrossRef] [PubMed]
- Forister, M.L.; Cousens, B.; Harrison, J.G.; Anderson, K.; Thorne, J.H.; Waetjen, D.; Nice, C.C.; Parsia, M.D.; Hladik, M.L.; Meese, R.; et al. Increasing neonicotinoid use and the declining butterfly fauna of lowland California. Biol. Lett. 2016, 12, 20160475. [Google Scholar] [CrossRef]
- Alkassab, A.T.; Kirchner, W.H. Sublethal exposure to neonicotinoids and related side effects on insect pollinators: Honeybees, bumblebees and solitary bees. J. Plant Dis. Prot. 2017, 124, 1–30. [Google Scholar] [CrossRef]
- Siviter, H.; Richman, S.K.; Muth, F. Field-realistic neonicotinoid exposure has sub-lethal effects on non-Apis bees: A meta-analysis. Ecol. Lett. 2021, 24, 2586–2597. [Google Scholar] [CrossRef] [PubMed]
- Braak, N.; Neve, R.; Jones, A.K.; Gibbs, M.; Breuker, C.J. The effects of insecticides on butterflies: A review. Environ. Pollut. 2018, 242, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Pecenka, J.R.; Lundgren, J.G. Non-target effects of clothianidin on monarch butterflies. Sci. Nat. 2015, 102, 19. [Google Scholar] [CrossRef]
- Krischik, V.; Rogers, M.; Gupta, G.; Varshney, A. Soil-applied imidacloprid translocates to ornamental flowers and reduces survival of adult Coleomegilla maculata, Harmonia axyridis and Hippodamia convergens Lady beetles and larval Danaus plexippus and Vanessa cardui butterflies. PLoS ONE 2015, 10, 1371. [Google Scholar] [CrossRef]
- Krishnan, N.; Zhang, Y.; Aust, M.E.; Hellmich, R.L.; Coats, J.R.; Bradbury, S.P. Monarch butterfly (Danaus plexippus) life-stage risks from foliar and seed treatment insecticides. Environ. Tox. Chem. 2021, 40, 1761–1777. [Google Scholar] [CrossRef]
- Krishnan, N.; Jurenka, R.A.; Bradbury, S.P. Neonicotinoids can cause arrested pupal ecdysis in lepidoptera. Sci. Rep. 2021, 11, 15787. [Google Scholar] [CrossRef]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef] [PubMed]
- David, A.; Botias, C.; Abdul-Sada, A.; Nicholls, E.; Rotheray, E.L.; Hill, E.M.; Goulson, D. Widespread contamination of wildflower and bee-collected pollen with complex mixtures of neonicotinoids and fungicides commonly applied to crops. Environ. Int. 2016, 88, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Douglas, M.; Rohr, J.R.; Tooker, J.F. Neonicotinoid insecticide travels through a soil food chain, disrupting biological control of non-target pests and decreasing soya bean yield. J. Appl. Ecol. 2014, 52, 250–260. [Google Scholar] [CrossRef]
- Sanchez-Bayo, F.; Hyne, R.V. Detection and analysis of neonicotinoids in river waters-development of a passive sampler for three commonly used insecticides. Chemosphere 2014, 99, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Bottias, C.; David, A.; Hill, E.M.; Goulson, D. Contamination of wild plants near neonicotinoid seed-treated crops and implications for non-target insects. Sci. Total Environ. 2016, 566, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Hladik, M.L.; Kolpin, D.W.; Kuivila, K.M. Widespread occurrence of neonicotinoid insecticides in streams in a high corn and soybean producing region, USA. Environ. Pollut. 2014, 193, 189–196. [Google Scholar] [CrossRef]
- Morrissey, C.A.; Mineau, P.; Devries, J.H.; Sanchez-Bayo, F.; Liess, M.; Cavallaro, M.C.; Liber, K. Neonicotinoid contamination of global surface waters and associated risk to aquatic invertebrates: A review. Environ. Int. 2015, 74, 291–303. [Google Scholar] [CrossRef]
- Schmidt, T.S.; Miller, J.L.; Mahler, B.J.; Van Metre, P.C.; Nowell, L.H.; Sandstrom, M.W.; Carlisle, D.M.; Moran, P.W.; Bradley, P.M. Ecological consequences of neonicotinoid mixtures in streams. Sci. Adv. 2022, 8, eabj8182. [Google Scholar] [CrossRef]
- Main, A.R.; Michel, N.L.; Cavallaro, M.C.; Headley, J.V.; Peru, K.M.; Morrissey, C.A. Snowmelt transport of neonicotinoid insecticides to Canadian prairie wetlands. Agric. Ecosys. Environ. 2016, 215, 76–84. [Google Scholar] [CrossRef]
- Keim, B. Backyard Pesticide May Fuel Bee Die-Offs. 2012. Available online: www.wired.com/2012/04/neonicotinoids-gardens/ (accessed on 10 June 2023).
- Morrison, B.A.; Xia, K.; Stewart, R.D. Evaluating neonicotinoid insecticide uptake by plants used as buffers and cover crops. Chemosphere 2023, 322, 138154. [Google Scholar] [CrossRef]
- James, D.G. A neonicotinoid insecticide at a rate found in nectar reduces longevity but not oogenesis in monarch butterflies, Danaus plexippus (L.) (Lepidoptera: Nymphalidae). Insects 2019, 10, 276. [Google Scholar] [CrossRef] [PubMed]
- Prouty, C.; Bartlett, L.J.; Krischik, V.; Altizer, S. Adult monarch butterflies show high tolerance to neonicotinoid insecticides. Ecol. Entomol. 2023, 48, 531–543. [Google Scholar] [CrossRef]
- Mullin, C.A. Effects of ‘inactive’ ingredients on bees. Curr. Opin. Insect Sci. 2015, 10, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Mesnage, R.; Antoniou, M.N. Ignoring adjuvant toxicity falsifies the safety profile of commercial pesticides. Front. Public Health 2018, 5, 361. [Google Scholar] [CrossRef] [PubMed]
- Nagy, K.; Duca, R.C.; Lovas, S.; Creta, M.; Scheepers, P.; Godderis, L.; Adam, B. Systematic review of comparative studies assessing the toxicity of pesticide active ingredients and their product formulations. Environ. Res. 2020, 181, 108926. [Google Scholar] [CrossRef]
- Krupke, C.H.; Holland, J.D.; Long, E.Y.; Eltzer, B.D. Planting of neonicotinoid-treated maize poses risks for honey bees and other non-target organisms over a wide area without consistent crop yield benefit. J. Appl. Ecol. 2017, 54, 1449–1458. [Google Scholar] [CrossRef]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzwizer, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.D.; et al. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2014, 22, 35–67. [Google Scholar] [CrossRef]
- Thompson, D.A.; Lehmler, H.J.; Kolpin, D.W.; Hladik, M.L.; Vargo, J.D.; Schilling, K.E.; LeFevre, G.H.; Peeples, T.L.; Poch, M.C.; LaDuca, L.E.; et al. A critical review on the potential impacts of neonicotinoid insecticide use: Current knowledge of environmental fate, toxicity and implications for human health. Environ. Sci. Process. Impacts 2020, 22, 1315–1346. [Google Scholar] [CrossRef]
- Thompson, D.A.; Kolpin, D.W.; Hladik, M.L.; Lehmler, H.J.; Meppelink, S.M.; Poch, M.C.; Vargo, J.D.; Soupene, V.A.; Irfan, N.M.; Robinson, M.; et al. Prevalence of neonicotinoid insecticides in paired private-well tap water and human urine samples in a region of intense agriculture overlying vulnerable aquifers in eastern Iowa. Chemosphere 2023, 319, 137904. [Google Scholar] [CrossRef]
- Zhou, Y.; Guo, J.; Wang, Z.; Sun, Z.; Yun, X.; Zhang, J. Levels and inhalation risk of neonicotinoid insecticides in fine particulate matter in urban and rural areas of China. Environ. Int. 2020, 142, 105822. [Google Scholar] [CrossRef]
- Zhang, D.; Lu, S. Human exposure to neonicotinoids and the associated health risks. Environ. Int. 2022, 163, 107201. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yu, W.; Zhang, L.; Cao, L.; Ling, J.; Liao, K.; Shen, G.; Du, W.; Chen, K.; Zhao, M.; et al. First evidence of neonicotinoid insecticides in human bile and associated hepatotoxicity risk. J. Hazard. Mater. 2023, 446, 130715. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.D.; Tooker, J.F. Neonicotinoids pose undocumented threats to food webs. Proc. Natl. Acad. Sci. USA 2020, 117, 22609–22613. [Google Scholar] [CrossRef] [PubMed]
- Kremer, A.N.; King, B.H. A neonicotinoid affects the mating behavior of Spalangia endius (Hymenoptera: Pteromalidae), a biological control agent of filth flies. Environ. Entomol. 2019, 48, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Muth, F.; Leonard, A.S. A neonicotinoid pesticide impairs foraging, but not learning, in free-flying bumblebees. Sci. Rep. 2019, 9, 4764. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.J.; Raine, N.E. Chronic impairment of bumblebee natural foraging behavior induced by sublethal pesticide exposure. Funct. Ecol. 2014, 28, 1459–1471. [Google Scholar] [CrossRef]
- Eng, M.L.; Stutchbury, B.J.M.; Morrissey, C.A. Imidacloprid and chlorpyrifos insecticides impair migratory ability in a seed-eating songbird. Sci. Rep. 2017, 7, 15176. [Google Scholar] [CrossRef]
- Eng, M.L.; Stutchbury, B.J.M.; Morrissey, C.A. A neonicotinoid reduces fueling and delays migration in songbirds. Science 2019, 365, 1177–1180. [Google Scholar] [CrossRef]
- English, S.G.; Sandoval-Herrera, N.I.; Bishop, C.A.; Bishop, M.; Maisonneuve, F.; Elliot, J.E.; Welch, K.C., Jr. Neonicotinoid pesticides exert metabolic effects on avian pollinators. Sci. Rep. 2021, 11, 2914. [Google Scholar] [CrossRef]
- Hallman, C.A.; Foppen, R.B.D.; Van Turnhout, C.A.M.; De Kroon, H.; Jongejans, E. Declines in insectivorous birds are associated with high neonicotinoid concentrations. Nature 2014, 511, 341–343. [Google Scholar] [CrossRef]
- Erti, H.M.H.; Mora, M.A.; Brightsmith, D.J.; Navarro-Alberto, J.A. Potential impact of neonicotinoid use on Northern Bobwhite (Colinus virginianus) in Texas: A historical analysis. PLoS ONE. 2018, 13, e0191100. [Google Scholar]
- Brower, L.P.; Williams, E.H.; Jaramillo-Lopez, P.; Kust, D.R.; Slayback, D.A.; Ramirez, M.I. Butterfly mortality and salvage logging from the March 2016 storm in the monarch butterfly biosphere reserve in Mexico. Am. Entomol. 2017, 63, 151–164. [Google Scholar] [CrossRef]
- James, D.G.; Kappen, L. Further insights on the migration biology of monarch butterflies, Danaus plexippus (Lepidoptera: Nymphalidae) from the Pacific Northwest. Insects 2021, 12, 161. [Google Scholar] [CrossRef]
- James, D.G. Do some fall migrants from the Pacific Northwest augment winter breeding populations of monarch butterflies in southern California? J. Lepid. Soc. 2018, 72, 244–246. [Google Scholar] [CrossRef]
- James, D.G. Western North American monarchs: Spiraling into oblivion or adapting to a changing environment? Anim. Migr. 2021, 8, 19–26. [Google Scholar] [CrossRef]
- James, D.G.; Schaefer, M.C.; Easton, K.K.; Carl, A. First population study on winter breeding monarch butterflies, Danaus plexippus (Lepidoptera: Nymphalidae) in the urban south bay of San Francisco. Insects 2021, 12, 946. [Google Scholar] [CrossRef] [PubMed]
- Smither, C.N. A note on overwintering in Danaus plexippus (Linnaeus) (Lepidoptera: Nymphalidae in Australia. Aust. Zool. 1965, 13, 135–136. [Google Scholar]
- James, D.G. Studies on a winter breeding population of Danaus plexippus (L.) (Lepidoptera: Nymphalidae) at Spencer, New South Wales. Gen. Appl. Ent. 1981, 13, 47–53. [Google Scholar]
- James, D.G. Population and general biology of non-reproductive colonies of the monarch butterfly, Danaus plexippus (L.) (Lepidoptera: Nymphalidae) in New South Wales. Aust. J. Zool. 1984, 32, 663–670. [Google Scholar] [CrossRef]
- James, D.G. Migration biology of the monarch butterfly in Australia. In Biology and Conservation of the Monarch Butterfly; Malcolm, S.B., Zalucki, M.P., Eds.; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1993; pp. 189–200. [Google Scholar]
- Goehring, L.; Oberhauser, K.S. Effects of photoperiod, temperature and host plant age on induction of reproductive diapause and development time in Danaus plexippus. Ecol. Entomol. 2002, 27, 674–685. [Google Scholar] [CrossRef]
- Mansingh, A. Physiological classification of dormancies in insects. Can. Entomol. 1971, 103, 983–1009. [Google Scholar] [CrossRef]
- Howard, E.; Aschen, H.; Davis, A.K. Citizen science observations of monarch butterfly overwintering in the southern United States. Psyche 2010, 2010, 689301. [Google Scholar] [CrossRef]
- Satterfield, D.A.; Maerz, J.C.; Hunter, M.D.; Flockhart, D.T.T.; Hobson, K.A.; Norris, D.R.; Streit, A.; De Roode, J.C.; Altizer, S. Migartory monarchs that encounter resident monarchs show life history differences and higher rates of parasite infection. Ecol. Lett. 2018, 21, 1670–1680. [Google Scholar] [CrossRef] [PubMed]
- James, D.G. Ovarian dormancy in Danaus plexippus (L.) (Lepidoptera: Nymphalidae)-Oligopause not diapause. J. Aust. Ent. Soc. 1982, 21, 31–35. [Google Scholar] [CrossRef]
- Talla, V.; Pierce, A.A.; Adams, K.L.; De Man, T.J.B.; Nallu, S.; Villablanca, F.X.; Kronforst, M.R.; De Roode, J.C. Genomic evidence for gene flow between monarchs with divergent migratory phenotypes and flight performance. Mol. Ecol. 2020, 29, 2567–2582. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.F.; Herman, W.S. Effect of photoperiod and temperature on reproduction of the monarch butterfly. J. Insect Physiol. 1976, 12, 1565–1568. [Google Scholar] [CrossRef]
- James, D.G. Effect of temperature upon energy reserves of the monarch butterfly, Danaus plexippus (L.) (Lepidoptera: Danaidae). Aust. J. Zool. 1986, 34, 373–379. [Google Scholar] [CrossRef]
- Brower, L.P.; Pyle, R.M. The interchange of migratory monarchs between Mexico and the western United States and the importance of floral corridors to the fall and spring migrations. In Conserving Migratory Pollinators and Nectar Corridors in the Western United States; Nabhan, G., Ed.; University of Arizona Press: Tucson, AZ, USA, 2004; pp. 144–166. [Google Scholar]
- Vane-Wright, R.I. The Columbus hypothesis: An explanation for the dramatic 19th Century range expansion of the monarch butterfly. In Biology and Conservation of the Monarch Butterfly; Malcolm, S.B., Zalucki, M.P., Eds.; Natural History Museum of Los Ange-les County: Los Angeles, CA, USA, 1993; pp. 179–187. [Google Scholar]
- Zalucki, M.P.; Clarke, A.R. Monarchs across the Pacific: The Columbus hypothesis revisited. Biol. J. Linn. Soc. 2004, 82, 111–121. [Google Scholar] [CrossRef]
- James, D.G. Overwintering Biology of the Monarch Butterfly, Danaus plexippus, in New South Wales. Ph.D. Thesis, Macquarie University, Sydney, NSW, Australia, 1984; p. 199. [Google Scholar]
- Garcia-Berro, A.; Talla, V.; Vila, R.; Kar Wai, H.; Shipilina, D.; Chan, K.G.; Pierce, N.E.; Backstrom, N.; Talavera, G. Migratory behavior is positively associated with genetic diversity in butterflies. Mol. Ecol. 2023, 32, 560–574. [Google Scholar] [CrossRef]
- Mann, M.E.; Gleick, P.H. Climate change and California drought in the 21st century. Proc. Natl. Acad. Sci. USA 2015, 112, 3858–3859. [Google Scholar] [CrossRef]
- Diffenbaugh, N.S.; Swain, D.L.; Touma, D. Anthropogenic warming has increased drought risk in California. Proc. Natl. Acad. Sci. USA 2015, 112, 3931–3936. [Google Scholar] [CrossRef]
- Urquhart, F.A.; Urquhart, N.R.; Munger, F. A study of a continuously breeding population of Danaus plexippus in southern California compared to a migratory population and its significance in the study of insect movement. J. Res. Lep. 1970, 7, 169–181. [Google Scholar]
- Neelin, J.D.; Langenbrunner, B.; Meyerson, J.; Hall, A.; Berg, N. California winter precipitation change under global warming in the coupled model intercomparison project phase 5 ensemble. J. Clim. 2013, 26, 6238–6256. [Google Scholar] [CrossRef]
- Oberhauser, K.S.; Brinda, S.J.; Weaver, S.; Moon, R.D.; Manweiler, S.A.; Read, N. Growth and survival of monarch butterflies (Lepidoptera: Danaidae) after exposure to permethrin barrier treatments. Environ. Entomol. 2006, 6, 1626–1634. [Google Scholar] [CrossRef]
- Krueger, A.J.; Hanford, K.; Weissling, T.J.; Velez, A.M.; Anderson, T.D. Pyrethroid exposure reduces growth and development of monarch butterfly (Lepidoptera: Nymphalidae) caterpillars. J. Insect Sci. 2021, 21, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, N.; Zhang, Y.; Bidne, K.G.; Hellmich, R.L.; Coats, J.R. Assessing field-scale risks of foliar insecticide applications to monarch butterfly (Danaus plexippus) larvae. Environ. Toxicol. Chem. 2020, 39, 923–941. [Google Scholar] [CrossRef] [PubMed]
- Runquist, E.; Nordmeyer, C.; Stapleton, S. Widespread annual occurrence of pesticides within designated critical habitats for endangered prairie butterflies. J. Insect Cons. 2024. [Google Scholar] [CrossRef]
- Whalon, M.E.; Wingerd, B.A. Bt: Mode of action and use. Arch. Insect Biochem. Physiol. 2003, 54, 200–211. [Google Scholar] [CrossRef]
- Peacock, J.W.; Schweitzer, D.F.; Carter, J.L.; Dubois, N.R. Laboratory assessment of the effects of Bacillus thuringiensis on native Lepidoptera. Environ. Entomol. 1998, 27, 450–457. [Google Scholar] [CrossRef]
- Redmond, C.T.; Wallis, L.; Geis, M.; Williamson, R.C.; Potter, D.A. Strengths and limitations of Bacillus thuringiensis galleriae for managing Japanese beetle (Popillia japonica) adults and grubs with caveats for cross-order activity to monarch butterfly (Danaus plexippus) larvae. Pest Manag. Sci. 2019, 76, 472–479. [Google Scholar] [CrossRef]
- Sparks, T.C.; Wessels, F.J.; Lorsbach, B.A.; Nugent, B.M.; Watson, G.B. The new age of insecticide discovery-the crop protection industry and the impact of natural products. Pestic. Biochem. Physiol 2019, 161, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, S.; Gotyal, B.S.; Babu, V.R. Role of novel insecticides in crop protection and their selectivity to natural enemies: A review. J. Environ. Biol. 2020, 41, 149–160. [Google Scholar] [CrossRef]
- Olaya-Arenas, P.; Hauri, K.; Scharf, M.E.; Kaplan, I. Larval pesticide exposure impacts monarch butterfly performance. Sci. Rep. 2020, 10, 14490. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.D.; Chen, X.D.; Johnson, C.S. Effects of herbicides on Behr’s Metalmark butterfly, a surrogate for the endangered Lange’s Metalmark. Environ. Pollut. 2012, 164, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Stevens, S.R.; Frey, D.F. Host plant pattern and variation in climate predict the location of natal grounds for migratory monarch butterflies in western North America. J. Insect Cons. 2010, 14, 731–744. [Google Scholar] [CrossRef]
- Spaeth, K.E.; Barbour, P.J.; Moranz, R.; Dinsmore, S.J.; Williams, C.J. Asclepias dynamics on US rangelands: Implications for conservation of monarch butterflies and other insects. Ecosphere 2022, 13, e03816. [Google Scholar] [CrossRef]
- Halsch, C.A.; Code, A.; Hoyle, S.M.; Fordyce, J.A.; Baert, N.; Forister, M.L. Pesticide contamination of milkweeds across the agricultural, urban and open spaces of low-elevation northern California. Front. Ecol. Evol. 2020, 8, 162. [Google Scholar] [CrossRef]
- Main, A.R.; Hladik, M.L.; Webb, E.B.; Goyne, K.W.; Mengel, D. Beyond neonicotinoids-Wild pollinators are exposed to a range of pesticides while foraging in agroecosystems. Sci. Tot. Environ. 2020, 742, 140436. [Google Scholar] [CrossRef]
- Halsch, C.A.; Hoyle, S.M.; Code, A.; Fordyce, J.A.; Forister, M.L. Milkweed plants bought at nurseries may expose monarch caterpillars to harmful pesticide residues. Biol. Cons. 2022, 273, 109699. [Google Scholar] [CrossRef]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western US forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef]
- Bonfils, C.; Santer, B.D.; Pierce, D.W.; Hildago, H.G.; Bala, G.; Das, T.; Barnett, T.P.; Cayan, D.R.; Doutriaux, C.; Wood, A.W.; et al. Detection and attribution of temperature changes in the mountainous western United States. J. Clim. 2008, 21, 6404–6424. [Google Scholar] [CrossRef]
- Hamlet, A.F.; Lettenmaier, D.P. Effects of 20th century warming and climate variability on flood risk in the western US. Water Resour. Res. 2007, 43, WO6427. [Google Scholar] [CrossRef]
- Zalucki, M.P. Temperature and rate of development in Danaus plexippus L. and D. chrysippus L (Lepidoptera: Nymphalidae). J. Aust. Ent. Soc. 1982, 21, 241–246. [Google Scholar] [CrossRef]
- Nail, K.R.; Batalden, R.V.; Oberhauser, K.S. What’s too hot and what’s too cold? Lethal and sub-lethal effects of extreme temperatures on developing monarchs. In Monarchs in a Changing World; Oberhauser, K.S., Nail, K.R., Altizer, S., Eds.; Cornell University Press: Ithaca, NY, USA, 2015; pp. 99–108. [Google Scholar]
- Shi, H.; Garcia-Reyes, M.; Jacox, M.G.; Rykaczewski, R.R.; Black, B.A.; Bograd, S.J.; Sydeman, W.J. Co-occurrence of California drought and northeast Pacific marine heatwaves under climate change. Geophys. Res. Lett. 2021, 48, e2021GL092765. [Google Scholar] [CrossRef]
- James, D.G. Population biology of monarch butterflies, Danaus plexippus L (Lepidoptera: Nymphalidae) at a milkweed-rich summer breeding site in central Washington. J. Lepid. Soc. 2016, 70, 182–193. [Google Scholar] [CrossRef]
- Leone, J.B.; Larson, D.L.; Larson, J.L.; Pennarola, N.; Oberhauser, K. Adult monarch (Danaus plexippus) abundance is higher in burned sites than grazed sites. Front. Ecol. Evol. 2019, 7, 435. [Google Scholar] [CrossRef]
- Xie, Y.; Lin, M.; Decharme, B.; Seferian, R. Tripling of western US particulate pollution from wildfires in a warming climate. Proc. Natl. Acad. Sci. USA 2022, 119, e2111372119. [Google Scholar] [CrossRef]
- Zipkin, E.F.; Ries, L.; Reeves, R.; Regetz, J.; Oberhauser, K. Tracking climate impacts on the migratory monarch butterfly. Glob. Chang. Biol. 2012, 18, 3039–3049. [Google Scholar] [CrossRef]
- Swain, D.L.; Langenbrunner, B.; Neelin, D.J.; Hall, A. Increasing precipitation volatility in twenty-first century California. Nat. Clim. Chang. 2018, 8, 427–433. [Google Scholar] [CrossRef]
- Leong, K.L.H. Evaluation and management of California monarch winter sites. West. Wildl. 2016, 3, 19–26. [Google Scholar]
- Leong, K.L.H. Microenvironmental factors associated with the winter habitat of the monarch butterfly (Lepidoptera: Danaidae) in central California. Ann. Entomol. Soc. Am. 1990, 83, 906–910. [Google Scholar] [CrossRef]
- Espeset, A.E.; Harrison, J.G.; Shapiro, A.M.; Nice, C.C.; Thorne, J.H.; Waetjen, D.P.; Fordyce, J.A.; Forister, M.L. Understanding a migratory species in a changing world: Climatic effects and demographic declines in the western monarch revealed by four decades of intensive monitoring. Oecologia 2016, 181, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Crone, E.E.; Pelton, E.M.; Brown, L.M.; Thomas, C.C.; Schultz, C.B. Why are monarch butterflies declining in the west? Understanding the importance of multiple correlated drivers. Ecol. Appl. 2019, 29, e01975. [Google Scholar] [CrossRef] [PubMed]
- Dilts, T.E.; Steele, M.O.; Engler, J.D.; Pelton, E.M.; Jepsen, S.J.; McKnight, S.J.; Taylor, A.R.; Fallon, C.E.; Black, S.H.; Cruz, E.E.; et al. Host plants and climate structure habitat associations of the western monarch butterfly. Front. Eco. Evol. 2019, 7, 188. [Google Scholar] [CrossRef]
- Luna, T.; Dumroese, R.K. Monarchs (Danaus plexippus) and milkweeds (Asclepias spp.) the current situation and methods for propagating milkweeds. Nativ. Plants J. 2013, 14, 5–16. [Google Scholar] [CrossRef]
- Waterbury, B.; Potter, A.; Svancara, L.K. Monarch butterfly distribution and breeding ecology in Idaho and Washington. Front. Ecol. Evol. 2019, 7, 172. [Google Scholar] [CrossRef]
- Morris, G.M.; Kline, C.; Morris, S.M. Status of Danaus plexippus population in Arizona. J. Lepid. Soc. 2015, 69, 91–107. [Google Scholar]
- Svancara, L.K.; Abatzoglou, J.T.; Waterbury, B. Modeling current and future potential distributions of milkweed and the monarch butterfly in Idaho. Front. Ecol. Evol. 2019, 7, 168. [Google Scholar] [CrossRef]
- Hartzler, R.G. Reduction in common milkweed (Asclepias syriaca) occurrence in Iowa cropland from 1999 to 2009. Crop Protect. 2010, 29, 1542–1544. [Google Scholar] [CrossRef]
- Flockhart, D.T.T.; Martin, T.G.; Norris, D.R. Experimental examination of intraspecific density-dependent competition during the breeding period in monarch butterflies (Danaus plexippus). PLoS ONE. 2012, 7, e45080. [Google Scholar] [CrossRef]
- Pleasants, J. Milkweed restoration in the mid-west for monarch butterfly recovery: Estimates of milkweed lost, milkweeds remaining and milkweeds that must be added to increase the monarch population. Insect Conserv. Divers. 2017, 10, 42–53. [Google Scholar] [CrossRef]
- Boyle, J.H.; Dalgleish, H.J.; Puzey, J.R. Monarch butterfly and milkweed declines substantially predate the use of genetically modified crops. Proc. Natl. Acad. Sci. USA 2019, 116, 3006–3011. [Google Scholar] [CrossRef] [PubMed]
- Zaviezo, T.; Munoz, A.E. Conservation biological control of arthropod pests using native plants. Curr. Opin. Insect Sci. 2023, 56, 101022. [Google Scholar] [CrossRef] [PubMed]
- James, D.G.; Price, T.S. Field-testing of methyl salicylate for recruitment and retention of beneficial insects and grapes in grapes and hops. J. Chem. Ecol. 2004, 30, 1613–1618. [Google Scholar] [CrossRef]
- Woods, J.L.; James, D.G.; Lee, J.C.; Gent, D.H. Validation of airborne methyl salicylate as a means to improve conservation biological control of spider mites in Oregon hop yards. Exp. Appl. Acarol. 2011, 55, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.; Gurr, G.M.; Simmons, A.T.; Wratten, S.D.; James, D.G.; Leeson, G.; Nicol, H.; Orre, G.U.S. Field evaluation of the ‘attract and reward’ approach in vineyards. Ann. Appl. Biol. 2011, 159, 69–78. [Google Scholar] [CrossRef]
- James, D.G.; Seymour, L.; Lauby, G.; Buckley, K. Beauty with benefits: Butterfly conservation in Washington State, USA, wine grape vineyards. J. Insect Cons. 2015, 19, 341–348. [Google Scholar] [CrossRef]
- James, D.G.; Seymour, L.; Lauby, G.; Buckley, K. Beneficial Insect Attraction to Milkweeds (Asclepias speciosa, Asclepias fascicularis) in Washington State, USA. Insects 2016, 7, 30. [Google Scholar] [CrossRef]
- Thakur, T.; Hurley, T. Do farmers need to be paid to grow milkweed or will they volunteer if it is easy enough? Appl. Econ. Perspect. Policy 2022, 45, 1008–1024. [Google Scholar] [CrossRef]
- Kasten, K.; Stenoien, C.; Caldwell, W.; Oberhauser, K.S. Can roadside habitat lead monarchs on a route to recovery? J. Insect Cons. 2016, 20, 1047–1057. [Google Scholar] [CrossRef]
- Cariveau, A.B.; Anderson, E.; Baum, K.A.; Hopwood, J.; Lonsdorf, E.; Nootenboom, C.; Tuerk, K.; Oberhauser, K.; Snell-Rood, E. Rapid assessment of roadsides as potential habitat for monarchs and other pollinators. Front. Ecol. Evol. 2019, 7, 386. [Google Scholar] [CrossRef]
- Lalonde, S.; McCune, J.L.; Rivest, S.A.; Kharouba, H.M. Decline in common milkweed along roadsides around Ottawa, Canada. Ecoscience 2022, 29, 25–37. [Google Scholar] [CrossRef]
- Hopwood, J.; Black, S.; Fleury, S. Roadside Best Management Practices that Benefit Pollinators; US Department of Transportation: Washington, DC, USA, 2015; p. FHWA-HEP-16-059. Available online: https://rosap.ntl.bts.gov/view/dot/55913 (accessed on 2 January 2024).
- Baum, K.A.; Sharber, W.V. Fire creates host plant patches for monarch butterflies. Biol. Lett. 2012, 8, 968–971. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.J. Enhancing monarch butterfly reproduction by mowing fields of common milkweed. Am. Mid. Nat. 2015, 173, 229–240. [Google Scholar] [CrossRef]
- Alcock, J.; Brower, L.P.; Williams, E.H. Monarch butterflies use regenerating milkweeds for reproduction in mowed hayfields in northern Virginia. J. Lepid. Soc. 2016, 70, 177–181. [Google Scholar] [CrossRef]
- Knight, S.M.; Norris, D.R.; Derbyshire, R.; Flockhart, D.T.T. Strategic mowing of roadside milkweeds increases monarch butterfly oviposition. Glob. Ecol. Cons. 2019, 19, e00678. [Google Scholar] [CrossRef]
- Haan, N.L.; Landis, D.A. Grassland disturbance increases monarch butterfly oviposition and decreases arthropod predator abundance. Biol. Cons. 2020, 233, 185–192. [Google Scholar] [CrossRef]
- Brown, J.J.; Chippendale, G.M. Migration of the monarch butterfly, Danaus plexippus: Energy sources. J. Insect Physiol. 1974, 20, 1117–1130. [Google Scholar] [CrossRef]
- Pyle, R.M. Chasing Monarchs: Migrating with the Butterflies of Passage; Houghton Mifflin Company: New York, NY, USA, 1999; p. 307. [Google Scholar]
- Hobson, K.A.; Garcia-Rubio, O.R.; Carrera-Trevino, R.; Anparasan, L.; Kardynal, K.J.; McNeil, J.N.; Garcia-Serrano, E.; Mora-Alvarez, B.X. Isotopic analysis of stored lipids in migratory and overwintering monarch butterflies (Danaus plexippus): Evidence for southern critical late-stage nectaring sites? Behav. Evol. Ecol. 2020, 8, 572140. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Inamine, H. Mechanisms behind the monarch’s decline. Science 2018, 360, 1294–1296. [Google Scholar] [CrossRef]
- Taylor, O.R.; Pleasants, J.M.; Grundel, R.; Pecoraro, S.D.; Lovett, J.P.; Ryan, A. Evaluating the migration mortality hypothesis using monarch tagging data. Front. Ecol. Evol. 2020, 8, 264. [Google Scholar] [CrossRef]
- Shepardson, L. The Butterfly Trees; The James Barry Company: San Francisco, CA, USA, 1914; p. 32. [Google Scholar]
- Williams, C.B. The Migration of Butterflies; Oliver: Edinburgh, Scotland; Boyd: London, UK, 1930; p. 473. [Google Scholar]
- Urquhart, F.A.; Beard, P.; Brownlee, R. A population study of the monarch butterfly (D. plexippus) in northern California. J Res. Lep. 1965, 4, 221–226. [Google Scholar]
- Herman, W.S. The endocrine basis of reproductive inactivity in monarch butterflies overwintering in central California. J. Insect Physiol. 1973, 19, 1883–1887. [Google Scholar] [CrossRef]
- Hill, H.F., Jr.; Wenner, A.M.; Wells, P.H. Reproductive behavior in an overwintering aggregation of monarch butterflies. Am. Mid. Nat. 1976, 95, 10–19. [Google Scholar] [CrossRef]
- Leong, K.L.H.; Frey, D.; Brenner, G.; Baker, S.; Fox, D. Use of multivariate analyses to characterize the monarch butterfly (Lepidoptera: Danaidae) winter habitat. Ann. Entomol. Soc. Am. 1971, 84, 263–267. [Google Scholar] [CrossRef]
- Frey, D.; Leong, K.L.H.; Fredericks, D.; Raskowitz, S. Clustering pattern of monarch butterfly (Lepidoptera: Danaidae) at two California central coast overwintering sites. Ann. Entomol. Soc. Am. 1992, 83, 907–910. [Google Scholar] [CrossRef]
- Borkin, S.S. Notes on shifting distribution patterns and survival of immature Danaus plexippus (Lepidoptera: Danaidae) on the food plant Asclepias syriaca. Great Lakes Entomol. 1982, 15, 199–205. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Kitching, R.L. Temporal and spatial variation of mortality in field populations of Danaus plexippus L. and D. chrysippus L. (Lepidoptera: Nymphalidae). Oecologia 1982, 53, 201–207. [Google Scholar] [CrossRef]
- De Anda, A.; Oberhauser, K.S. Invertebrate natural enemies and stage-specific mortality rates of monarch eggs and larvae. In Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly; Oberhauser, K.S., Nail, K.R., Altizer, S., Eds.; Cornell University Press: Ithaca, NY, USA, 2015; pp. 60–70. [Google Scholar]
- Grant, T.J.; Flockhart, D.T.T.; Blader, T.R.; Hellmich, R.L.; Pitman, G.M.; Tyner, S.; Norris, D.R.; Bradbury, S.P. Estimating arthropod survival probability from field counts: A case study with monarch butterflies. Ecosphere 2020, 11, e03082. [Google Scholar] [CrossRef]
- Myers, A.T.; Haan, N.L.; Landis, D.A. Video surveillance reveals a community of largely nocturnal Danaus plexippus (L.) egg predators. J. Insect Cons. 2020, 24, 731–737. [Google Scholar] [CrossRef]
- Hermann, S.L.; Blackledge, C.; Haan, N.L.; Myers, A.T.; Landis, D.A. Predators of monarch butterfly eggs and neonates are more diverse than previously recognized. Sci. Rep. 2019, 9, 14304. [Google Scholar] [CrossRef] [PubMed]
- McCoshum, S.M.; Andreoli, S.L.; Stenoien, C.M.; Oberhauser, K.S.; Baum, K.A. Species distribution models for natural enemies of the monarch butterfly (Danaus plexippus) larvae and pupae: Distribution patterns and implications for conservation. J. Insect Cons. 2016, 20, 223–227. [Google Scholar] [CrossRef]
- Stevenson, M.; Hudman, K.L.; Scott, A.; Contreras, K.; Kopachena, J.G. High survivorship of first-generation monarch butterfly eggs to third instar associated with a diverse arthropod community. Insects 2021, 12, 567. [Google Scholar] [CrossRef] [PubMed]
- Oberhauser, K.S.; Wiederholt, R.; Diffendorfer, J.E.; Semmens, D.; Ries, L.; Thogmartin, W.E.; Lopez-Hoffman, L.; Semmens, B. A trans-national monarch butterfly population model and implications for regional conservation priorities. Ecol. Entomol. 2017, 42, 51–60. [Google Scholar] [CrossRef]
- Brower, L.P.; Glazier, S.C. Localization of heart poisons in the monarch butterfly. Science 1975, 188, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.M.; Potter, D.A. Invasive paper wasp turns urban pollinator gardens into ecological traps for monarch butterfly larvae. Sci. Rep. 2020, 10, 9553. [Google Scholar] [CrossRef]
- McGruddy, R.A.; Howse, M.W.F.; Haywood, J.; Ward, C.J.I.; Staufer, T.B.; Hayek-Williams, M.; Tift, R.J.; Lester, P.J. Invasive paper wasps have strong cascading effects on the host plant of monarch butterflies. Ecol. Entomol. 2020, 46, 459–469. [Google Scholar] [CrossRef]
- Oberhauser, K.S.; Elmquist, D.; Perilla-Lopez, J.M.; Gebhard, I.; Lukens, L.; Stireman, J. Tachinid fly (Diptera: Tachinidae) parasitoids of Danaus plexippus (Lepidoptera: Nymphalidae). Ann. Entomol. Soc. Am. 2017, 110, 536–543. [Google Scholar] [CrossRef]
- Schaefer, M.C.; James, D.G. Viability of monarch butterfly (Danaus plexippus) (L.) (Lepidoptera: Nymphalidae) pupae during winter in the South Bay area of San Francisco, California. J. Lepid. Soc. 2024, in press. [Google Scholar]
- Stenoien, C.; McCoshum, S.; Caldwell, W.; De Anda, A.; Oberhauser, K.S. New reports that monarch butterflies (Lepidoptera: Nymphalidae, Danaus plexippus Linnaeus) are hosts for a pupal parasitoid (Hymenoptera: Chalcidoidae, Pteromalus cassotis Walker). J. Kans. Entomol. Soc. 2015, 88, 16–26. [Google Scholar] [CrossRef]
- Myers, J.H.; Corey, J.S. Ecology and evolution of pathogens in natural populations of Lepidoptera. Evol. Appl. 2016, 9, 231–247. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, M.E. Viruses as costs to gregarious feeding behavior in the Lepidoptera. Oikos 1991, 61, 291–296. [Google Scholar] [CrossRef]
- Arnott, H.J.; Smith, K.M.; Fullilove, S.L. Ultrastructure of a cytoplasmic polyhedrosis virus affecting the monarch butterfly, Danaus plexippus. I. Development of virus and normal polyhedra in the larva. J. Ultrastruct. Res. 1968, 24, 479–507. [Google Scholar] [CrossRef] [PubMed]
- Altizer, S.M.; Oberhauser, K.S. Effects of the protozoan parasite, Ophryocystis elektroscirrha on the fitness of monarch butterflies (Danaus plexippus). J. Invert. Pathol. 1999, 74, 76–88. [Google Scholar] [CrossRef] [PubMed]
- De Roode, J.C.; Yates, A.J.; Altizer, S.M. Virulence-transmission trade-offs and population divergence in virulence in a naturally occurring butterfly parasite. Proc. Natl. Acad. Sci. USA 2008, 105, 7489–7494. [Google Scholar] [CrossRef] [PubMed]
- Bradley, C.A.; Altizer, S.M. Parasites hinder monarch butterfly flight: Implications for disease spread in migratory hosts. Ecol. Lett. 2005, 8, 290–300. [Google Scholar] [CrossRef]
- Leong, K.L.H.; Kaya, H.K.; Yoshimura, M.A.; Frey, D.F. The occurrence and effect of a protozoan parasite, Ophryocystis elektroscirrha (Neogregarinida: Ophryocystidae) on overwintering monarch butterflies, Danaus plexippus (Lepidoptera: Danaidae) from two California winter sites. Ecol. Entomol. 1992, 17, 338–342. [Google Scholar] [CrossRef]
- Altizer, S.M.; Oberhauser, K.S.; Brower, L.P. Associations between host migration and the prevalence of a protozoan parasite in natural populations of adult monarch butterflies. Ecol. Entomol. 2000, 25, 125–139. [Google Scholar] [CrossRef]
- Satterfield, D.A.; Villablanca, F.X.; Maerz, J.C.; Altizer, S. Migratory monarchs wintering in California experience low infection risk compared to monarchs breeding year-round on native milkweed. Integr. Comp. Biol. 2016, 56, 343–352. [Google Scholar] [CrossRef]
- De Roode, J.C.; Altizer, S. Host-parasite genetic interactions and virulence-transmission relationships in natural populations of monarch butterflies. Evolution 2010, 64, 502–514. [Google Scholar] [CrossRef]
- Majewska, A.A.; Davis, A.K.; Altizer, S.; De Roode, J.C. Parasite dynamics in North American monarchs predicted by host density and seasonal migratory culling. J. Anim. Ecol. 2022, 91, 780–793. [Google Scholar] [CrossRef] [PubMed]
- Majewska, A.A.; Satterfield, D.A.; Harrison, R.B.; Altizer, S.; Hepinstall-Cymerman, J. Urbanization predicts infection risk by a protozoan parasite in non-migratory populations of monarch butterflies from the southern coastal US and Hawaii. Landsc. Ecol. 2019, 34, 649–661. [Google Scholar] [CrossRef]
- Majewska, A.A.; Altizer, S.M. Exposure to non-native tropical milkweed promotes reproductive development in migratory monarch butterflies. Insects 2019, 10, 253. [Google Scholar] [CrossRef] [PubMed]
- James, D.G. Induction of reproductive dormancy in Australian monarch butterflies, Danaus plexippus. Aust. J. Zool. 1983, 31, 497–504. [Google Scholar] [CrossRef]
- James, D.G.; James, T.A. Migration and overwintering in Australian monarch butterflies, Danaus plexippus (L.) (Lepidoptera: Nymphalidae): A review with new observations and research needs. J. Lepid. Soc. 2019, 73, 177–190. [Google Scholar] [CrossRef]
- Pasternak, C. How to Raise Monarch Butterflies: A Step-by-Step Guide for Kids (How It Works); Firefly Books: Richmond Hill, ON, Canada, 2012; p. 48. [Google Scholar]
- McGovern, S.F. Raising Monarchs; Sunbury Press: Boiling Springs, PA, USA, 2013; p. 52. [Google Scholar]
- Gustafsson, K.M.; Agrawal, A.A.; Lewenstein, B.V.; Wolf, S.A. The monarch butterfly through time and space: The social construction of an icon. BioScience 2015, 65, 612–622. [Google Scholar] [CrossRef]
- Zhang, W.; Goodale, E.; Chen, J. How contact with nature affects children’s biophilia, biophobia and conservation attitude in China. Biol. Cons. 2014, 177, 109–116. [Google Scholar] [CrossRef]
- Rosa, C.D.; Profice, C.C.; Collado, S. Nature experiences and adult’s self-reported pro-environmental behaviors: The role of connectedness to nature and childhood nature experiences. Front. Psychol. 2018, 9, 1055. [Google Scholar] [CrossRef]
- Whitburn, J.; Linklater, W.L.; Milfont, T.L. Exposure to urban nature and tree-planting are related to pro-environmental behavior via connection to nature, the use of nature for psychological restoration and environmental attitudes. Environ. Behav. 2019, 51, 787–810. [Google Scholar] [CrossRef]
- Hayward, M.W.; Meyer, N.F.V.; Balkenhol, N.; Beranek, C.T.; Bugir, C.K.; Bushell, K.V.; Callen, A.; Dickman, A.J.; Griffin, A.S.; Haswell, P.M.; et al. Intergenerational inequity: Stealing the joy and benefits of nature from our children. Front. Ecol. Evol. 2022, 10, 830830. [Google Scholar] [CrossRef]
- Tenger-Trolander, A.; Lu, W.; Noyes, M.; Kronforst, M.R. Contemporary loss of migration in monarch butterflies. Proc. Natl. Acad. Sci. USA 2019, 10, 1073. [Google Scholar] [CrossRef]
- Tenger-Trolander, A.; Kronforst, M.R. Migration behaviour of commercial monarchs reared outdoors and wild-derived monarchs reared indoors. Proc. R. Soc. B 2020, 287, 20201326. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.K.; Smith, F.M.; Ballew, A.M. A poor substitute for the real thing: Captive-reared monarch butterflies are weaker, paler and have less-elongated wings than wild migrants. Biol. Lett. 2020, 16, 20190922. [Google Scholar] [CrossRef]
- Oberhauser, K.S. Captivity concerns for monarch butterflies. Nature 2019, 573, 501–502. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, A.A.E.; Newman, A.E.M.; Raine, N.E.; Mitchell, G.W.; Norris, D.R. Captive-reared migratory monarch butterflies show natural orientation when released in the wild. Cons. Physiol. 2021, 9, coab32. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.K. Captive-reared migratory monarchs fly in the wrong direction: A critique of Wilcox et al. Cons. Physiol. 2021, 9, coab063. [Google Scholar] [CrossRef]
- Steffy, G. Trends observed in fall migrant monarch butterflies (Lepidoptera: Nymphalidae) east of the Appalachian Mountains at an inland stopover in southern Pennsylvania over an eighteen-year period. Ann. Entomol. Soc. Am. 2015, 108, 718–728. [Google Scholar] [CrossRef]
- James, D.G.; Kappen, L.; Russell, T.; Harryman, B.; Bly, C.; James, T.S.; Seymour, L. Citizen scientist tagging reveals destinations and routes of migrating Monarch butterflies (Danaus plexippus (L.)) from the Pacific Northwest. J. Lepid. Soc. 2018, 72, 127–144. [Google Scholar]
- Wensler, R.J. The ultrastructure of the indirect flight muscles of the monarch butterfly, Danaus plexippus (L.) with implications for fuel utilization. Acta Zool. 1977, 58, 157–167. [Google Scholar] [CrossRef]
- Pelling, A.E.; Wilkinson, P.R. Dynamic mechanical oscillations during metamorphosis of the monarch butterfly. J. R. Soc. Interface 2009, 6, 29–37. [Google Scholar] [CrossRef]
- Baril, T.; Hayward, A. Migrators within migrators: Exploring transposable element dynamics in the monarch butterfly, Danaus plexippus. Mob. DNA 2022, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Pocius, V.M.; Majewska, A.A.; Freedman, M.G. The role of experiments in monarch butterfly conservation: A review of recent studies and approaches. Ann. Entomol. Soc. Am. 2022, 115, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Freedman, M.G.; de Roode, J.C.; Forister, M.L.; Kronforst, M.R.; Pierce, A.A.; Schultz, C.B.; Taylor, O.R.; Crone, E.E. Are eastern and western monarch butterflies distinct populations? A review of evidence for ecological, phenotypic and genetic differentiation and implications for conservation. Cons. Sci. Prac. 2021, 3, e432. [Google Scholar] [CrossRef]
- Cooper, C. Citizen Science: How Ordinary People Are Changing the Face of Discovery; The Overlook Press: New York, NY, USA, 2016; p. 294. [Google Scholar]
- James, D.G. Murderers touched by the magic of Monarchs. News Lepid. Soc. 2016, 58, 127–129. [Google Scholar]
- Soga, M.; Gaston, K.J. Extinction of experience: The loss of human-nature interactions. Front. Ecol. Environ. 2016, 14, 94–101. [Google Scholar] [CrossRef]
- Pyle, R.M. The Extinction of Experience. In Through a Green Lens; Pyle, R.M., Ed.; Oregon State University Press: Corvallis, OR, USA, 2016; pp. 53–58. [Google Scholar]
- Gaston, K.J.; Soga, M. Extinction of experience: The need to be more specific. People Nat. 2020, 2, 575–581. [Google Scholar] [CrossRef]
- Wagner, D.L. Insect declines in the anthropocene. Ann. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef]
- Yang, L.H. Complexity, humility and action: A current perspective on monarchs in western North America. Curr. Opin. Insect Sci. 2023, 59, 101078. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Year | Total Monarchs Reported | Number of Sites |
|---|---|---|
| 2018 | 27,721 | 213 |
| 2019 | 29,436 | 242 |
| 2020 | 1899 | 249 |
| 2021 | 247,246 | 284 |
| 2022 | 335,479 | 272 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, D.G. Monarch Butterflies in Western North America: A Holistic Review of Population Trends, Ecology, Stressors, Resilience and Adaptation. Insects 2024, 15, 40. https://doi.org/10.3390/insects15010040
James DG. Monarch Butterflies in Western North America: A Holistic Review of Population Trends, Ecology, Stressors, Resilience and Adaptation. Insects. 2024; 15(1):40. https://doi.org/10.3390/insects15010040
Chicago/Turabian StyleJames, David G. 2024. "Monarch Butterflies in Western North America: A Holistic Review of Population Trends, Ecology, Stressors, Resilience and Adaptation" Insects 15, no. 1: 40. https://doi.org/10.3390/insects15010040
APA StyleJames, D. G. (2024). Monarch Butterflies in Western North America: A Holistic Review of Population Trends, Ecology, Stressors, Resilience and Adaptation. Insects, 15(1), 40. https://doi.org/10.3390/insects15010040