
Winter Rains Support Butterfly Diversity, but Summer Monsoon Rainfall Drives Post-Monsoon Butterfly Abundance in the Arid Southwest of the US

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Sites
2.2. Butterfly Surveys
2.3. Environmental Data
2.4. Variable Selection
2.5. Modeling
3. Results
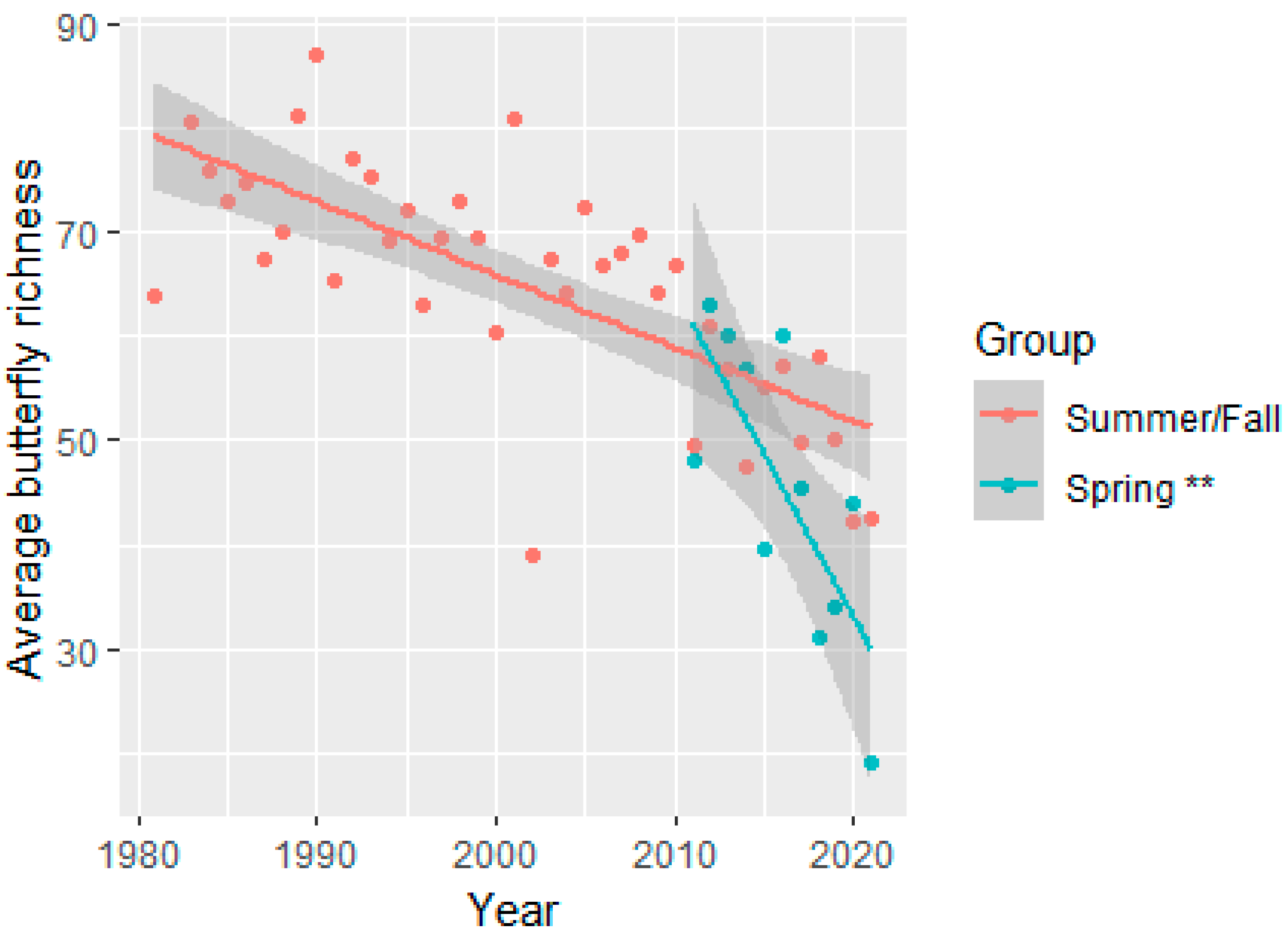
3.1. Butterfly Abundance and Richness Declines over Time and Observer Effort
3.2. Effect of Climate Patterns on Butterfly Abundance
3.3. Effect of Climate Patterns on Butterfly Richness
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated Modern Human-Induced Species Losses: Entering the Sixth Mass Extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide Decline of the Entomofauna: A Review of Its Drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic Valuation of the Vulnerability of World Agriculture Confronted with Pollinator Decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Losey, J.E.; Vaughan, M. The Economic Value of Ecological Services Provided by Insects. BioScience 2006, 56, 311–323. [Google Scholar] [CrossRef]
- Warren, M.S.; Maes, D.; van Swaay, C.A.M.; Goffart, P.; van Dyck, H.; Bourn, N.A.D.; Wynhoff, I.; Hoare, D.; Ellis, S. The Decline of Butterflies in Europe: Problems, Significance, and Possible Solutions. Proc. Natl. Acad. Sci. USA 2021, 118, e2002551117. [Google Scholar] [CrossRef]
- Wagner, D.L. Insect Declines in the Anthropocene. Annu. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef]
- Kucherov, N.B.; Minor, E.S.; Johnson, P.P.; Taron, D.; Matteson, K.C. Butterfly Declines in Protected Areas of Illinois: Assessing the Influence of Two Decades of Climate and Landscape Change. PLoS ONE 2021, 16, e0257889. [Google Scholar] [CrossRef]
- Duffy, K.; Gouhier, T.C.; Ganguly, A.R. Climate-Mediated Shifts in Temperature Fluctuations Promote Extinction Risk. Nat. Clim. Chang. 2022, 12, 1037–1044. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Diamond, S.E.; Buckley, L.B. Heat Stress and the Fitness Consequences of Climate Change for Terrestrial Ectotherms. Funct. Ecol. 2013, 27, 1415–1423. [Google Scholar] [CrossRef]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in Fluctuating Thermal Environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef]
- Bale, J.S.; Gerday, C.; Parker, A.; Marahiel, M.A.; Shanks, I.A.; Davies, P.L.; Warren, G. Insects and Low Temperatures: From Molecular Biology to Distributions and Abundance. Philos. Trans. R. Soc. B Biol. Sci. 2002, 357, 849–862. [Google Scholar] [CrossRef]
- Turnock, W.J.; Fields, P.G. Winter Climates and Coldhardiness in Terrestrial Insects. Eur. J. Entomol. 2005, 102, 561–576. [Google Scholar] [CrossRef]
- Marshall, K.E.; Sinclair, B.J. The Impacts of Repeated Cold Exposure on Insects. J. Exp. Biol. 2012, 215, 1607–1613. [Google Scholar] [CrossRef]
- Forister, M.L.; Halsch, C.A.; Nice, C.C.; Fordyce, J.A.; Dilts, T.E.; Oliver, J.C.; Prudic, K.L.; Shapiro, A.M.; Wilson, J.K.; Glassberg, J. Fewer Butterflies Seen by Community Scientists across the Warming and Drying Landscapes of the American West. Science 2021, 371, 1042–1045. [Google Scholar] [CrossRef]
- Georgescu, M.; Broadbent, A.M.; Balling, R.C. Effect of Increased Greenhouse Gas Concentration on Mean, Extreme, and Timing of Precipitation over Arizona (USA). Int. J. Climatol. 2022, 42, 3776–3792. [Google Scholar] [CrossRef]
- Lian, X.; Piao, S.L.; Chen, A.P.; Huntingford, C.; Fu, B.J.; Li, L.Z.X.; Huang, J.P.; Sheffield, J.; Berg, A.M.; Keenan, T.F.; et al. Multifaceted Characteristics of Dryland Aridity Changes in a Warming World. Nat. Rev. Earth Environ. 2021, 2, 232–250. [Google Scholar] [CrossRef]
- Williams, A.P.; Cook, B.I.; Smerdon, J.E. Rapid Intensification of the Emerging Southwestern North American Megadrought in 2020–2021. Nat. Clim. Chang. 2022, 12, 232–234. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2014. [Google Scholar]
- Larsen, E.A.; Belitz, M.W.; Guralnick, R.P.; Ries, L. Consistent Trait-Temperature Interactions Drive Butterfly Phenology in Both Incidental and Survey Data. Sci. Rep. 2022, 12, 13370. [Google Scholar] [CrossRef]
- Gezon, Z.J.; Lindborg, R.J.; Savage, A.; Daniels, J.C. Drifting Phenologies Cause Reduced Seasonality of Butterflies in Response to Increasing Temperatures. Insects 2018, 9, 174. [Google Scholar] [CrossRef]
- Kharouba, H.M.; Paquette, S.R.; Kerr, J.T.; Vellend, M. Predicting the Sensitivity of Butterfly Phenology to Temperature over the Past Century. Glob. Chang. Biol. 2014, 20, 504–514. [Google Scholar] [CrossRef]
- Dennis, E.B.; Morgan, B.J.T.; Brereton, T.M.; Roy, D.B.; Fox, R. Using Citizen Science Butterfly Counts to Predict Species Population Trends. Conserv. Biol. 2017, 31, 1350–1361. [Google Scholar] [CrossRef]
- Dennis, E.B.; Morgan, B.J.T.; Roy, D.B.; Brereton, T.M. Urban Indicators for UK Butterflies. Ecol. Indic. 2017, 76, 184–193. [Google Scholar] [CrossRef]
- Settele, J.; Kudrna, O.; Harpke, A.; Kühn, I.; van Swaay, C.; Verovnik, R.; Warren, M.; Wiemers, M.; Hanspach, J.; Hickler, T.; et al. Climatic Risk Atlas of European Butterflies. BioRisk 2008, 1, 1–712. [Google Scholar] [CrossRef]
- WallisDeVries, M.F.; Baxter, W.; van Vliet, A.J.H. Beyond Climate Envelopes: Effects of Weather on Regional Population Trends in Butterflies. Oecologia 2011, 167, 559–571. [Google Scholar] [CrossRef]
- Gutbrodt, B.; Mody, K.; Dorn, S. Drought Changes Plant Chemistry and Causes Contrasting Responses in Lepidopteran Herbivores. Oikos 2011, 120, 1732–1740. [Google Scholar] [CrossRef]
- Klockmann, M.; Gunter, F.; Fischer, K. Heat Resistance throughout Ontogeny: Body Size Constrains Thermal Tolerance. Glob. Chang. Biol. 2017, 23, 686–696. [Google Scholar] [CrossRef]
- Long, O.M.; Warren, R.; Price, J.; Brereton, T.M.; Botham, M.S.; Franco, A.M.A. Sensitivity of UK Butterflies to Local Climatic Extremes: Which Life Stages Are Most at Risk? J. Anim. Ecol. 2017, 86, 108–116. [Google Scholar] [CrossRef]
- Welti, E.A.R.; Zajicek, P.; Frenzel, M.; Ayasse, M.; Bornholdt, T.; Buse, J.; Classen, A.; Dziock, F.; Engelmann, R.A.; Englmeier, J.; et al. Temperature Drives Variation in Flying Insect Biomass across a German Malaise Trap Network. Insect Conserv. Divers. 2022, 15, 168–180. [Google Scholar] [CrossRef]
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T.; et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 1999, 399, 579–584. [Google Scholar] [CrossRef]
- Dubertret, F.; Tourneau, F.M.L.; Villarreal, M.L.; Norman, L.M. Monitoring Annual Land Use/Land Cover Change in the Tucson Metropolitan Area with Google Earth Engine (1986–2020). Remote Sens. 2022, 14, 2127. [Google Scholar] [CrossRef]
- Hutchinson, C.F.; Unruh, J.D.; Bahre, C.J. Land Use vs. Climate as Causes of Vegetation Change: A Study in SE Arizona. Glob. Environ. Chang. 2000, 10, 47–55. [Google Scholar] [CrossRef]
- Jenerette, G.D.; Wu, J. Analysis and Simulation of Land-Use Change in the Central Arizona—Phoenix Region, USA. Landsc. Ecol. 2001, 16, 611–626. [Google Scholar] [CrossRef]
- Jacobs, K.L.; Garfin, G.M.; Morehouse, B.J. Climate Science and Drought Planning: The Arizona Experience. J. Am. Water Resour. Assoc. 2005, 41, 437–446. [Google Scholar] [CrossRef]
- Scanlon, B.R.; Reedy, R.C.; Faunt, C.C.; Pool, D.; Uhlman, K. Erratum: Enhancing Drought Resilience with Conjunctive Use and Managed Aquifer Recharge in California and Arizona (Source Title (2016) 11 (035013)). Environ. Res. Lett. 2016, 11, 049501. [Google Scholar] [CrossRef]
- Krauss, J.; Bommarco, R.; Guardiola, M.; Heikkinen, R.K.; Helm, A.; Kuussaari, M.; Lindborg, R.; Ockinger, E.; Partel, M.; Pino, J.; et al. Habitat Fragmentation Causes Immediate and Time-Delayed Biodiversity Loss at Different Trophic Levels. Ecol. Lett. 2010, 13, 597–605. [Google Scholar] [CrossRef]
- Brereton, T.; Roy, D.B.; Middlebrook, I.; Botham, M.; Warren, M. The Development of Butterfly Indicators in the United Kingdom and Assessments in 2010. J. Insect Conserv. 2011, 15, 139–151. [Google Scholar] [CrossRef]
- Thomas, J.A. Monitoring Change in the Abundance and Distribution of Insects Using Butterflies and Other Indicator Groups. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 339–357. [Google Scholar] [CrossRef]
- Samways, M.J. Insect Conservation: A Synthetic Management Approach. Annu. Rev. Entomol. 2007, 52, 465–487. [Google Scholar] [CrossRef]
- Prudic, K.L.; Cruz, T.M.P.; Winzer, J.I.B.; Oliver, J.C.; Melkonoff, N.A.; Verbais, H.; Hogan, A. Botanical Gardens Are Local Hotspots for Urban Butterflies in Arid Environments. Insects 2022, 13, 865. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Average Spring Min Temp (°C) | Average Spring Max Temp (°C) | Average Summer/Fall Min Temp (°C) | Average Summer/Fall Max Temp (°C) | Average Annual Precipitation (mm) |
|---|---|---|---|---|---|
| 1. Cottonwood | 7.28 (4.74) | 26.45 (6.84) | 14.67 (5.24) | 32.60 (5.52) | 328.73 (85.13) |
| 2. McDowell Sonoran Preserve | 14.97 (5.25) | 27.61 (6.43) | 22.82 (4.43) | 34.50 (5.01) | 332.18 (128.28) |
| 3. Grand Canyon Desert View | 4.26 (5.64) | 19.87 (7.03) | 11.32 (5.31) | 26.64 (5.89) | 340.72 (85.13) |
| 4. Grand Canyon South Rim | −0.40 (5.06) | 17.66 (7.05) | 6.90 (5.17) | 24.65 (5.73) | 372.68 (95.20) |
| 5. Sycamore Creek | 11.95 (5.24) | 26.70 (6.65) | 19.63 (4.52) | 33.26 (5.17) | 433.27 (183.62) |
| 7. Ramsey Canyon | 8.97 (4.98) | 25.82 (6.03) | 15.14 (4.09) | 29.69 (4.09) | 458.69 (116.32) |
| 8. Boyce Thompson Arboretum | 12.06 (5.32) | 25.78 (6.48) | 19.21 (4.33) | 31.99 (4.75) | 463.56 (185.35) |
| 9. Grand Canyon North Rim | 2.56 (5.65) | 17.59 (7.12) | 9.61 (5.16) | 24.66 (5.95) | 498.12 (148.32) |
| 10. Atascosa Highlands | 8.32 (4.78) | 25.92 (5.95) | 15.17 (4.22) | 30.05 (3.95) | 499.72 (108.85) |
| 11. Sabino Canyon | 9.67 (5.34) | 24.68 (6.42) | 16.55 (4.04) | 29.84 (4.39) | 552.50 (190.40) |
| 12. Portal | 4.85 (4.98) | 21.60 (6.00) | 11.17 (4.21) | 25.47 (4.01) | 585.46 (149.55) |
| 13. Santa Rita Mountains | 9.24 (5.28) | 23.60 (6.32) | 15.65 (3.71) | 27.48 (4.16) | 657.68 (175.22) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rowe, H.I.; Johnson, B.; Broatch, J.; Cruz, T.M.P.; Prudic, K.L. Winter Rains Support Butterfly Diversity, but Summer Monsoon Rainfall Drives Post-Monsoon Butterfly Abundance in the Arid Southwest of the US. Insects 2024, 15, 5. https://doi.org/10.3390/insects15010005
Rowe HI, Johnson B, Broatch J, Cruz TMP, Prudic KL. Winter Rains Support Butterfly Diversity, but Summer Monsoon Rainfall Drives Post-Monsoon Butterfly Abundance in the Arid Southwest of the US. Insects. 2024; 15(1):5. https://doi.org/10.3390/insects15010005
Chicago/Turabian StyleRowe, Helen Ivy, Bradly Johnson, Jennifer Broatch, Terese Maxine Papag Cruz, and Kathleen L. Prudic. 2024. "Winter Rains Support Butterfly Diversity, but Summer Monsoon Rainfall Drives Post-Monsoon Butterfly Abundance in the Arid Southwest of the US" Insects 15, no. 1: 5. https://doi.org/10.3390/insects15010005
APA StyleRowe, H. I., Johnson, B., Broatch, J., Cruz, T. M. P., & Prudic, K. L. (2024). Winter Rains Support Butterfly Diversity, but Summer Monsoon Rainfall Drives Post-Monsoon Butterfly Abundance in the Arid Southwest of the US. Insects, 15(1), 5. https://doi.org/10.3390/insects15010005