
Diagnostic Tool for the Identification of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) Using Real-Time PCR

 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection
2.2. DNA Extraction
2.3. Design of the Primers and Probe for the qPCR Assays
2.4. Validation Method for the qPCR Probe
3. Results
3.1. DNA Extraction
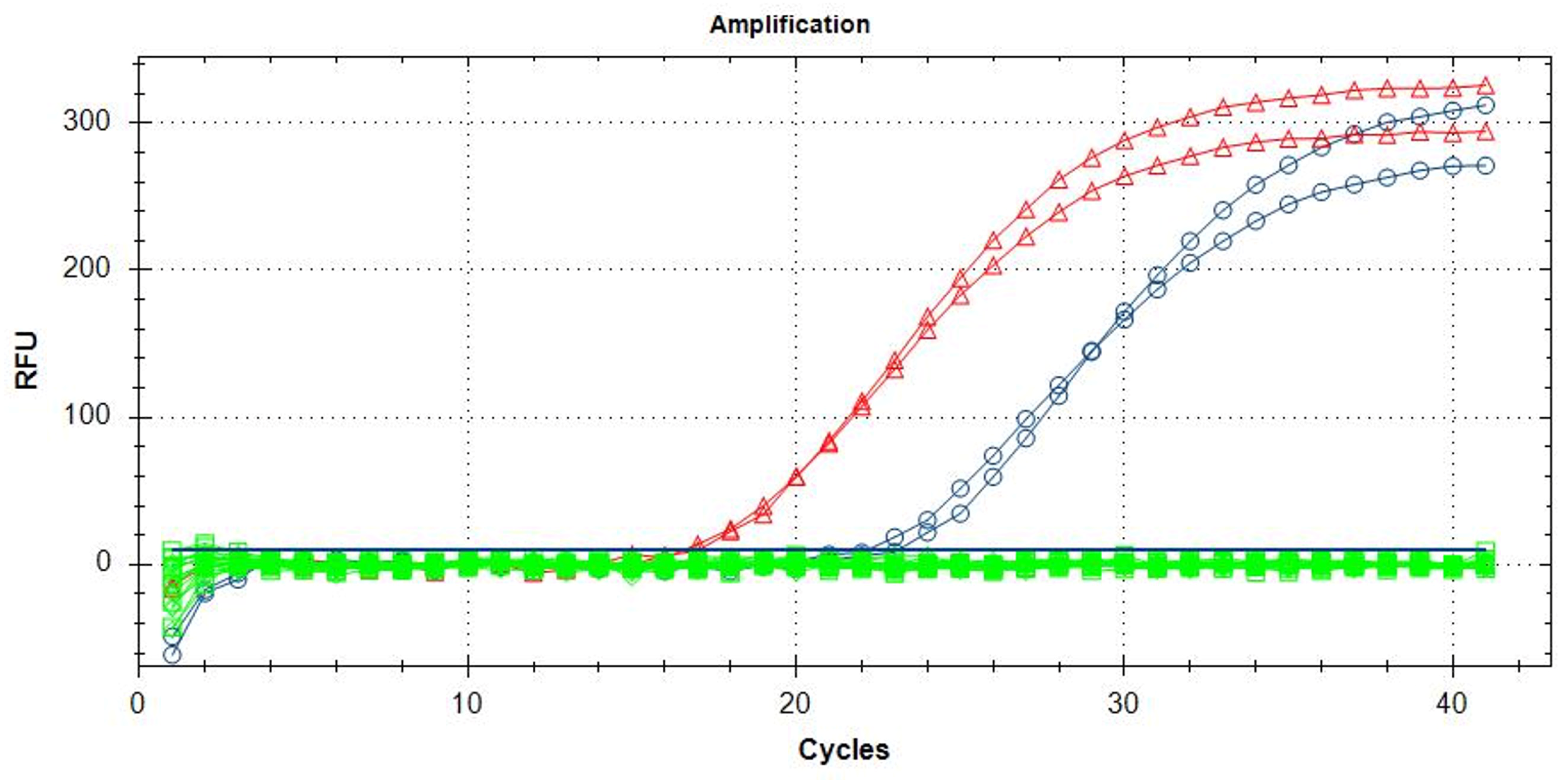
3.2. Assay Conditions of the TaqMan Probe Protocol
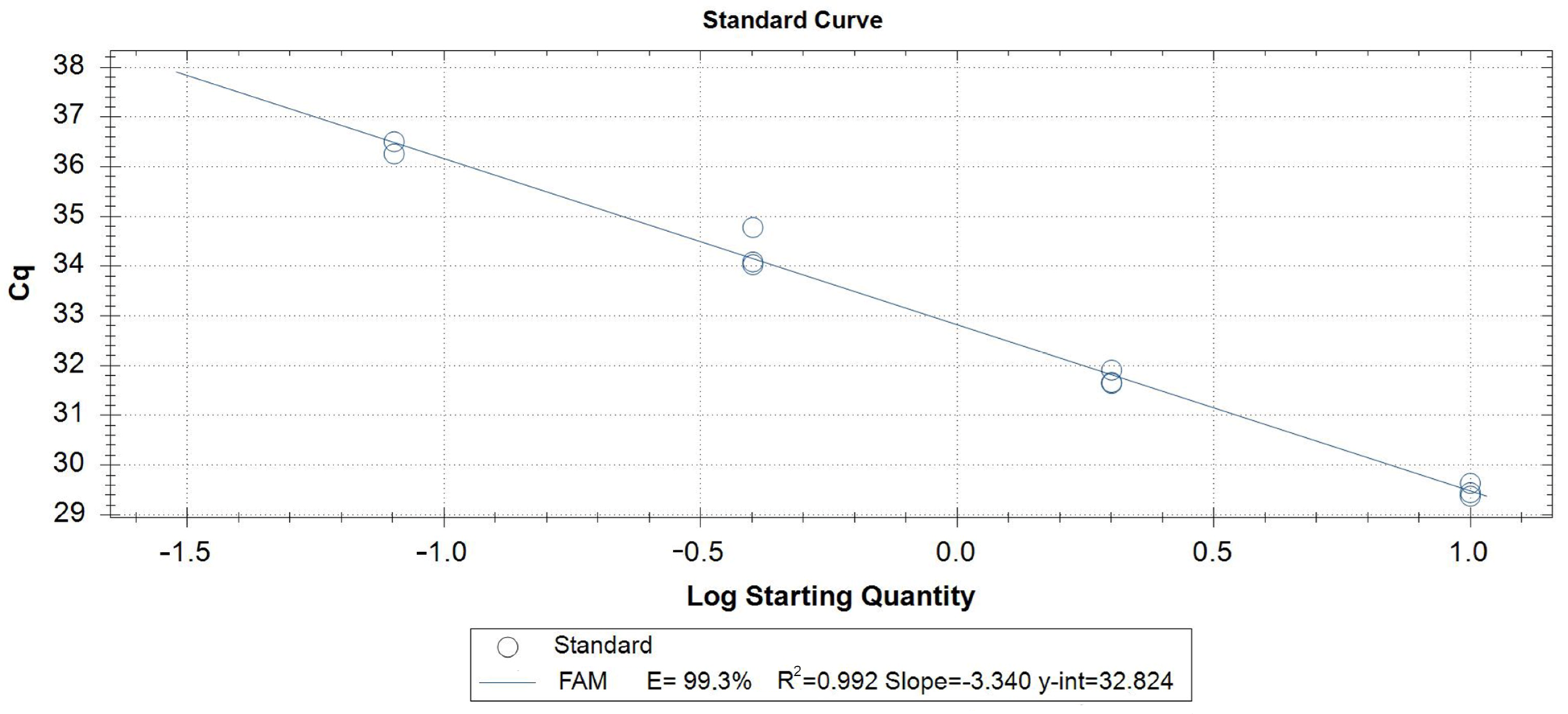
3.3. Validation of the Proposed Methods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drew, R.A.I.; Hancock, D.L. The Bactrocera dorsalis complex of fruit flies (Diptera: Tephritidae: Dacinae) in Asia. Bull. Entomol. Res. Suppl. Ser. 1994, 2, 1–68. [Google Scholar] [CrossRef]
- Clarke, A.R.; Armstrong, K.F.; Carmichael, A.E.; Milne, J.R.; Raghu, S.; Roderick, G.K.; Yeates, D.K. Invasive phytophagous pests arising through a recent tropical evolutionary radiation: The Bactrocera dorsalis complex of fruit flies. Annu. Rev. Entomol. 2005, 50, 293–319. [Google Scholar] [CrossRef] [PubMed]
- Doorenweerd, C.; Jose, M.S.; Barr, N.; Rubinoff, D.; Geib, S. Genomic data reveal new species and the limits of mtDNA barcode diagnostics to contain a global pest species complex (Diptera: Tephritidae: Dacinae). Syst. Entomol. 2023, 12616. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority); Loomans, A.; Diakaki, M.; Kinkar, M.; Schenk, M.; Vos, S. Pest survey card on Bactrocera dorsalis. EFSA Support. Publ. 2019, 16, 24. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). Pest survey card on Bactrocera dorsalis. EFSA Support. Publ. 2022, 2022, EN-7206. Available online: https://efsa.europa.eu/plants/planthealth/monitoring/surveillance/bactrocera-dorsalis (accessed on 1 September 2023).
- Doorenweerd, C.; Leblanc, L.; Norrbom, A.L.; San Jose, M.; Rubinoff, R. A global checklist of the 932 fruit fly species in the tribe Dacini (Diptera, Tephritidae). ZooKeys 2018, 730, 17–54. [Google Scholar] [CrossRef]
- San Jose, M.; Doorenweerd, C.; Geib, S.; Barr, N.; Dupuis, J.R.; Leblanc, L.; Kauwe, A.; Morris, K.Y.; Rubinoff, D. Interspecific gene flow obscures phylogenetic relationships in an important insect pest species complex. Mol. Phylogenet. Evol. 2023, 188, 107892 . [Google Scholar] [CrossRef]
- Drew, R.A.; Romig, M.C. Keys to the Tropical Fruit Flies (Tephritidae: Dacinae) of South-East Asia: Indomalaya to North-West Australasia; CABI: Oxfordshire, UK, 2013. [Google Scholar]
- Schutze, M.K.; Aketarawong, N.; Amornsak, W.; Armstrong, K.F.; Augustinos, A.A.; Barr, N.; Bo, W.; Bourtzis, K.; Boykin, L.M.; Cáceres, C. Synonymization of key pest species within the Bactrocera dorsalis species complex (Diptera: Tephritidae): Taxonomic changes based on a review of 20 years of integrative morphological, molecular, cytogenetic, behavioural and chemoecological data. Syst. Entomol. 2015, 40, 456–471. [Google Scholar] [CrossRef]
- Schutze, M.K.; Mahmood, K.; Pavasovic, A.; Bo, W.; Newman, J.; Clarke, A.R.; Krosch, M.N.; Cameron, S.L. One and the same: Integrative taxonomic evidence that Bactrocera invadens (Diptera: Tephritidae) is the same species as the Oriental fruit fly Bactrocera dorsalis. Syst. Entomol. 2015, 40, 472–486. [Google Scholar] [CrossRef]
- EFSA PLH Panel; Mertens, J.; Schenk, M.; Delbianco, A.; Graziosi, I.; Vos, S. Pest survey card on Bactrocera zonata. EFSA Support. Publ. 2021, 18, EN-1999. [Google Scholar] [CrossRef]
- Nugnes, F.; Russo, E.; Viggiani, G.; Bernardo, U. First record of an invasive fruit fly belonging to Bactrocera dorsalis complex (Diptera: Tephritidae) in Europe. Insects 2018, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Boykin, L.; Schutze, M.; Krosch, M.; Chomic, A.; Chapman, T.; Englezou, A.; Armstrong, K.; Clarke, A.; Hailstones, D.; Cameron, S. Multi-gene phylogenetic analysis of south-east Asian pest members of the Bactrocera dorsalis species complex (Diptera: Tephritidae) does not support current taxonomy. J. Appl. Entomol. 2014, 138, 235–253. [Google Scholar] [CrossRef]
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; 601 pp. + addendum; CAB International: Wallingford, UK, 1994. [Google Scholar]
- EPPO Bactrocera dorsalis. EPPO Datasheets on Pests Recommended for Regulation. 2023. Available online: https://gd.eppo.int (accessed on 27 July 2023).
- De Villiers, M.; Hattingh, V.; Kriticos, D.J.; Brunel, S.; Vayssières, J.F.; Sinzogan, A.; Billah, M.K.; Mohamed, S.A.; Mwatawala, M.; Abdelgader, H.; et al. The potential distribution of Bactrocera dorsalis: Considering phenology and irrigation patterns. Bull. Entomol. Res. 2016, 106, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Song, Y.; Meng, L.; Liu, R.; Lin, J.; Zhao, M.; Nie, C.; Li, Y.; Ji, Q. Climate warming affects phenology of Bactrocera dorsalis: A case study of Fujian and Guangxi, China. Bull. Insectology 2023, 76, 73–81. [Google Scholar]
- Jaffar, S.; Arif, S.; Rizvi, H.; Lu, Y. Understanding the invasion, ecological adaptations, and management strategies of Bactrocera dorsalis in China: A Review. Horticulturae 2023, 9, 1004. [Google Scholar] [CrossRef]
- Jiang, F.; Liang, L.; Wang, J.; Zhu, S. Chromosome-level genome assembly of Bactrocera dorsalis reveals its adaptation and invasion mechanisms. Commun. Biol. 2022, 5, 25. [Google Scholar] [CrossRef]
- Mutamiswa, R.; Nyamukondiwa, C.; Chikowore, G.; Chidawanyika, F. Overview of oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) in Africa: From invasion, bio-ecology to sustainable management. Crop Prot. 2021, 141, 105492. [Google Scholar] [CrossRef]
- Regulation (EU) 2019/2072 of 28 November 2019, Establishing Uniform Conditions for the Implementation of Regulation (EU) 2016/2031 of the European Parliament and the Council, as Regards Protective Measures against Pests of Plants, and Repealing Commission Regulation (EC) No 690/2008 and Amending Commission Implementing Regulation (EU) 2018/2019. 10.12.2019 Official Journal of the European Union L 319/1. Available online: http://data.europa.eu/eli/reg_impl/2019/2072/2020-10-06 (accessed on 18 January 2023).
- EU. Commission Delegated Regulation (EU) 2019/1702 of 1 August 2019 Supplementing Regulation (EU) 2016/2031 of the European Parliament and of the Council by Establishing the List of Priority Pests. 2019. Available online: http://data.europa.eu/eli/reg_del/2019/1702/oj (accessed on 23 May 2023).
- Europhyt. Available online: https://webgate.ec.europa.eu/europhyt/ (accessed on 27 July 2023).
- Vitiello, M.; de Benedetta, F.; Gargiulo, S.; Griffo, R.; Nugnes, F.; Bernardo, U. Bactrocera dorsalis in Campania: Insediamento o incursione? Entomata 2020, 13, 83–92. [Google Scholar]
- Gargiulo, S.; Nugnes, F.; De Benedetta, F.; Bernardo, U. Bactrocera latifrons in Europe: The importance of the right attractant for detection. Bull. Insectology 2021, 74, 311–320. [Google Scholar]
- Bernardo, U.; Miele, F.; Ascolese, R.; Carbone, C.; Griffo, R.; Nugnes, F. La Genesi del Processo Invasivo di Bactrocera dorsalis in Italia. Un Evento Ineludibile? CNIE: Palermo, Italy, 2023. [Google Scholar]
- Nugnes, F.; Miele, F.; Carbone, C.; Ascolese, R.; Figlioli, L.; Bernardo, U. Studi Preliminari Sulla Variabilità Genetica di Bactrocera dorsalis e Dinamica dell’infestazione in Campania; CNIE: Palermo, Italy, 2023. [Google Scholar]
- Griffo, R.; Notaro, L.; Nugnes, F.; Ghezzi, M.; Bernardo, U. Piano d’azione Della Regione Campania per Bactrocera dorsalis; CNIE: Palermo, Italy, 2023. [Google Scholar]
- Balmès, V.; Mouttet, R. Development and validation of a simplified morphological identification key for larvae of tephritid species most commonly intercepted at import in Europe. EPPO Bull. 2017, 47, 91–99. [Google Scholar] [CrossRef]
- Pace, R.; Ascolese, R.; Miele, F.; Russo, E.; Griffo, R.V.; Bernardo, U.; Nugnes, F. The bugs in the bags: The risk associated with the introduction of small quantities of fruit and plants by airline passengers. Insects 2022, 13, 617. [Google Scholar] [CrossRef] [PubMed]
- Kamayev, I.O.; Galinskaya, T.V.; Ovtshinnikova, O.G. Variability of the mandibular sclerite and its value in diagnostics of the third instar larvae of the Mediterranean Fruit Fly Ceratitis capitata (Wiedemann, 1824) (Diptera, Tephritidae). Entomol. Rev. 2020, 100, 1254–1264. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Norrbom, A.L.; Korytkowski, C.A.; Zucchi, R.A.; Uramoto, K.; Venable, G.L.; McCormick, J.; Dallwitz, M.J. Anastrepha and Toxotrypana: Descriptions, Illustrations, and Interactive Keys. 2012 onwards Version: 9 April 2019. Available online: https://www.delta-intkey.com/anatox/index.htm (accessed on 10 January 2023).
- Virgilio, M.; White, I.; De Meyer, M. A set of multi-entry identification keys to African frugivorous flies (Diptera, Tephritidae). ZooKeys 2014, 428, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Plant Health Australia. The Australian Handbook for the Identification of Fruit Flies. Version 3.1. Plant Health Australia. Canberra, ACT. 2018. Available online: https://www.fruitflyidentification.org.au/ (accessed on 27 July 2023).
- Rizzo, D.; Luchi, N.; Da, D.; Linda, L.; Francesco, B.; Giovanni, N.; Bruscoli, T.; Salemi, C.; Griffo, R.V.; Garonna, A.P.; et al. Development of a loop—Mediated isothermal amplification (LAMP) assay for the identification of the invasive wood borer Aromia bungii (Coleoptera: Cerambycidae) from frass. Biotech 2021, 11, 85. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, D.; Taddei, A.; Da Lio, D.; Nugnes, F.; Barra, E.; Stefani, L.; Bartolini, L.; Griffo, R.V.; Spigno, P.; Cozzolino, L.; et al. Identification of the Red-Necked Longhorn Beetle Aromia bungii (Faldermann, 1835) (Coleoptera: Cerambycidae) with real-Time PCR on frass. Sustainability 2020, 12, 6041. [Google Scholar] [CrossRef]
- Ioos, R.; Fourrier, C.; Iancu, G.; Gordon, T.R. Sensitive detection of Fusarium circinatum in pine seed by combining an enrichment procedure with a real-time polymerase chain reaction using dual-labeled probe chemistry. Phytopathology 2009, 99, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Kralik, P.; Ricchi, M. A basic guide to real time PCR in microbial diagnostics: Definitions, parameters, and everything. Front. Microbiol. 2017, 8, 108. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment Software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- EPPO. PM 7/98 (5) Specific requirements for laboratories preparing accreditation for a plant pest diagnostic activity. EPPO Bull. 2021, 51, 468–498. [Google Scholar] [CrossRef]
- EN ISO 16140; Standards for Validation and Verification of Microbiology Methods. International Organization for Standardization: Geneva, Switzerland, 2003.
- Rizzo, D.; Moricca, S.; Bracalini, M.; Benigno, A.; Bernardo, U.; Luchi, N.; Da Lio, D.; Nugnes, F.; Cappellini, G.; Salemi, C.; et al. Rapid detection of Pityophthorus juglandis (Blackman) (Coleoptera, Curculionidae) with the loop-mediated isothermal amplification (lamp) method. Plants 2021, 10, 1048. [Google Scholar] [CrossRef] [PubMed]
- Teter, S.; Steffen, L. Real-Time qPCR: Guidelines for a Comparison of Reagent Performance. 2017. Available online: https://ita.promega.com/resources/pubhub/applications-notes/an299-real-time-qpcr-guidelines-for-a-comparison-of-reagent-performance/ (accessed on 1 September 2023).
- de Benedetta, F.; Gargiulo, S.; Miele, F.; Figlioli, L.; Innangi, M.; Audisio, P.; Nugnes, F.; Bernardo, U. The spread of Carpophilus truncatus is on the razor’s edge between an outbreak and a pest invasion. Sci. Rep. 2022, 12, 18841. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.; Tseng, S.-P.; Hsu, P.-W.; Lu, C.-W.; Yang, C.-C.S.; Lin, C.-C. Introduction of a non-native lineage is linked to the recent black cocoa ant, Dolichoderus thoracicus (Smith, 1860), outbreaks in Taiwan. Taiwania 2022, 67, 271–279. [Google Scholar] [CrossRef]
- Bernardo, U.; Nugnes, F.; Gargiulo, S.; Nicoletti, R.; Becchimanzi, A.; Stinca, A.; Viggiani, G. An integrative study on Asphondylia spp. (Diptera: Cecidomyiidae), causing flower galls on Lamiaceae, with description, phenology, and associated fungi of two new species. Insects 2021, 12, 958. [Google Scholar] [CrossRef] [PubMed]
- Tajadini, M.; Panjehpour, M.; Javanmard, S.H. Comparison of SYBR Green and TaqMan methods in quantitative real time polymerase chain reaction analysis of four adenosine receptor subtypes. Adv. Biomed. Res. 2014, 3, 85. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, D.; Da Lio, D.; Bartolini, L.; Cappellini, G.; Bruscoli, T.; Salemi, C.; Aronadio, A.; Del Nista, D.; Pennacchio, F.; Boersma, N.; et al. Development of three molecular diagnostic tools for the identification of the false codling moth (Lepidoptera: Tortricidae). J. Econ. Entomol. 2021, 114, 1796–1807. [Google Scholar] [CrossRef] [PubMed]
- Kitano, D.; Takakura, K.I. Simple and on-site DNA purification for LAMP reaction applicable to non-adult tephritid fruit fly (Diptera: Tephiritidae). J. Appl. Entomol. 2020, 144, 824–829. [Google Scholar] [CrossRef]
- Afroz, S.; Nomana, S.; Zhang, Y.; Ali, Y.; Mahmud, R.; Li, Z. Species identification of economic important adult fruit flies based on DNA Barcoding (MT DNA COI) and larvae based on species specific primers from Central and South part of Bangladesh. Malays. J. Sustain. Agric. 2022, 6, 103–109. [Google Scholar] [CrossRef]
- Andrews, K.J.; Bester, R.; Manrakhan, A.; Maree, H.J. A multiplex PCR assay for the identification of fruit flies (Diptera: Tephritidae) of economic importance in South Africa. Sci. Rep. 2022, 12, 13089. [Google Scholar] [CrossRef]
- Dhami, M.K.; Gunawardana, D.N.; Voice, D.; Kumarasinghe, L. A real-time PCR toolbox for accurate identification of invasive fruit fly species. J. Appl. Entomol. 2016, 140, 536–552. [Google Scholar] [CrossRef]
- Koohkanzade, M.; Marzieh, M.; Dhami, M.K.; Fekrat, L.; Namaghi, S.H. Rapid identification of Bactrocera zonata (Dip.: Tephritidae) using TaqMan real-time PCR assay. PLoS ONE 2018, 13, e0205136. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Sample Code | Life Stage | Geographical Origin of Samples | Supplier |
|---|---|---|---|---|
| Anastrepha fraterculus (Wiedemann, 1830) | MR 000752 | Adult | Ecuador | UoF |
| MR 000795 | Adult | Ecuador | UoF | |
| Anastrepha leptozona (Hendel, 1914) | MR 000817 | Adult | Ecuador | UoF |
| MR 000823 | Adult | Ecuador | UoF | |
| MR 001658 | Adult | Ecuador | UoF | |
| Anastrepha ludens (Loew, 1873) | MR 000753 | Adult | USA | UoF |
| MR 000820 | Adult | USA | UoF | |
| MR 001659 | Adult | USA | UoF | |
| MR 001710 | Adult | USA | UoF | |
| MR 001721 | Adult | USA | UoF | |
| Anastrepha obliqua (Macquart, 1835) | MR 000821 | Adult | Ecuador | UoF |
| MR 001715 | Adult | Ecuador | UoF | |
| Anastrepha serpentina (Wiedemann, 1830) | MR 000283 | Adult | USA | UoF |
| MR 000822 | Adult | USA | UoF | |
| MR 001701 | Adult | USA | UoF | |
| MR 001689 | Adult | USA | UoF | |
| MR 001722 | Adult | USA | UoF | |
| Bactrocera dorsalis (Hendel, 1912) | MR 001712 | Adult | Italy | UoF |
| MR 001720 | Adult | Italy | IPSP | |
| MR 001709 | Adult | Italy | IPSP | |
| MR 001718 | Adult | Italy | IPSP | |
| MR 001716 | Adult | Italy | IPSP | |
| MR 000814 | Adult | Italy | IPSP | |
| MR 001563 | Adult | Italy | IPSP | |
| MR 001713 | Adult | Italy | IPSP | |
| MR 001638 | Adult | Italy | IPSP | |
| MR 001714 | Adult | Italy | IPSP | |
| MR 001717 | Adult | Italy | IPSP | |
| MR 001711 | Larva | Italy | IPSP | |
| MR 001719 | Larva | Italy | IPSP | |
| MR 000801 | Larva | Italy | IPSP | |
| MR 001683 | Larva | Italy | IPSP | |
| MR 001684 | Larva | Italy | IPSP | |
| MR 000239 | Larva | Italy | IPSP | |
| MR 001697 | Larva | Italy | IPSP | |
| MR 001747 | Larva | Italy | IPSP | |
| MR 000764 | Larva | Italy | IPSP | |
| Bactrocera latifrons (Hendel, 1915) | MR 000752 | Adult | Italy | IPSP |
| MR 000823 | Adult | Italy | IPSP | |
| Bactrocera oleae (Rossi, 1790) | MR 001658 | Adult | Italy | UoF |
| Bactrocera zonata (Saunders, 1842) | MR 000753 | Adult | Afghanistan | UoF |
| Ceratitis capitata (Wiedemann, 1824) | MR 000820 | Adult | Italy | UoF |
| MR 000821 | Larva | Italy | UoF | |
| MR 000283 | Adult | Italy | IPSP | |
| MR 000822 | Larva | Italy | PPS-T | |
| Cydia pomonella (Linnaeus, 1758) | MR 000259 | Adult | Italy | UoF |
| MR 000790 | Adult | Italy | UoF | |
| Grapholita (Aspila) molesta (Busck, 1916) | MR 001617 | Adult | Italy | UoF |
| Rhagoletis cerasi (Linnaeus, 1758) | MR 001618 | Adult | Italy | UoF |
| Rhagoletis completa (Cresson, 1929) | MR 001648 | Larva | Italy | UoP |
| MR 001619 | Adult | Italy | UoF | |
| Thaumatotibia leucotreta (Meyrick, 1913) | MR 001678 | Adult | South Africa | XSIT |
| MR 001620 | Adult | South Africa | XSIT | |
| MR 001621 | Larvae | Interception at the port of Leghorn (Italy) | PPS-T |
| Name | Sequence | Size (bp) | Reference Sequence |
|---|---|---|---|
| Bdors_235F | CACCAGTCATATTGTGAG | 105 | OP056621.1 |
| Bdors_340R | GTGTCATGAAGAATAATATCTAC | ||
| Bdors_308P | FAM-ATGACGACCTATGCTCTGAACTTGT-BHQ1 |
| Sample | DNA Conc (ng/µL) ± SD | A260/280 Ratio | Cq (18S) |
|---|---|---|---|
| Adult | 25.0 ± 2.68 | 1.9 ± 0.12 | 18.4 ± 0.81 |
| Larva | 203.2 ± 90.50 | 1.8 ± 0.08 | 16.2 ± 0.14 |
| Dilutions 1:5 | qPCR Probe |
|---|---|
| 10.0 ng/µL | 17.5 ± 0.26 |
| 2.0 ng/µL | 20.4 ± 0.19 |
| 0.4 ng/µL | 22.1 ± 0.12 |
| 0.08 ng/µL | 23.9 ± 0.20 |
| 0.016 ng/µL | 27.4 ± 0.13 |
| 3.2 pg/µL | 28.3 ± 0.51 |
| 0.64 pg/µL | 30.2 ± 0.13 |
| 0.128 pg/µL | 33.0 ± 0.09 |
| 25.6 fg/µL | - |
| Sample | qPCR Probe Test | |
|---|---|---|
| Repeatability | Reproducibility | |
| 1 | 24.5 ± 0.66 | 24.9 ± 0.40 |
| 2 | 24.5 ± 0.62 | 24.5 ± 0.20 |
| 3 | 24.1 ± 0.25 | 24.4 ± 0.15 |
| 4 | 24.2 ± 0.31 | 24.3 ± 0.22 |
| 5 | 24.3 ± 0.04 | 24.6 ± 0.35 |
| 6 | 24.4 ± 0.40 | 24.3 ± 0.17 |
| 7 | 24.4 ± 0.27 | 24.2 ± 0.21 |
| 8 | 24.5 ± 0.41 | 24.5 ± 0.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizzo, D.; Zubieta, C.G.; Sacchetti, P.; Marrucci, A.; Miele, F.; Ascolese, R.; Nugnes, F.; Bernardo, U. Diagnostic Tool for the Identification of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) Using Real-Time PCR. Insects 2024, 15, 44. https://doi.org/10.3390/insects15010044
Rizzo D, Zubieta CG, Sacchetti P, Marrucci A, Miele F, Ascolese R, Nugnes F, Bernardo U. Diagnostic Tool for the Identification of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) Using Real-Time PCR. Insects. 2024; 15(1):44. https://doi.org/10.3390/insects15010044
Chicago/Turabian StyleRizzo, Domenico, Claudia Gabriela Zubieta, Patrizia Sacchetti, Andrea Marrucci, Fortuna Miele, Roberta Ascolese, Francesco Nugnes, and Umberto Bernardo. 2024. "Diagnostic Tool for the Identification of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) Using Real-Time PCR" Insects 15, no. 1: 44. https://doi.org/10.3390/insects15010044
APA StyleRizzo, D., Zubieta, C. G., Sacchetti, P., Marrucci, A., Miele, F., Ascolese, R., Nugnes, F., & Bernardo, U. (2024). Diagnostic Tool for the Identification of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) Using Real-Time PCR. Insects, 15(1), 44. https://doi.org/10.3390/insects15010044