
Development of a Technique Using Artificial Membrane for In Vitro Rearing of Body Lice Pediculus humanus humanus

 ,
,  ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Body Louse Strain and Conservation Condition
2.2. In Vitro Louse Maintenance
2.2.1. Artificial Blood-Feeding Apparatus: Rearing System
2.2.2. Blood Meal Support
2.2.3. Blood Meal Composition
2.3. In Vivo Louse Maintenance
2.4. Rearing Follow-Up
2.4.1. Surveillance
2.4.2. Electron Microscopy
2.4.3. Blood Meal
2.5. Strategy Validation
2.6. Cost Calculations and Statistical Analysis
3. Results
3.1. General Observation and Electron Microscopy
3.2. Blood Meal
3.2.1. Anticoagulants
3.2.2. Support
3.2.3. Additions
3.3. Blood Meal Inspection In Vivo
3.3.1. Culture
3.3.2. Optical Microscopy
3.4. Other Observations
3.5. Strategy Validation
3.6. Cost
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raoult, D.; Roux, V. The Body Louse as a Vector of Reemerging Human Diseases. Clin. Infect. Dis. 1999, 29, 888–911. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, R.; Drancourt, M.; Raoult, D. The Role of Louse-Transmitted Diseases in Historical Plague Pandemics. Lancet Infect. Dis. 2021, 21, e17–e25. [Google Scholar] [CrossRef]
- Amanzougaghene, N.; Fenollar, F.; Raoult, D.; Mediannikov, O. Where Are We With Human Lice? A Review of the Current State of Knowledge. Front. Cell. Infect. Microbiol. 2020, 9, 474. [Google Scholar] [CrossRef]
- Pietri, J.E.; Ray, R. A Simplified Protocol for in Vitro Rearing of Human Body Lice. Parasite 2020, 27, 8. [Google Scholar] [CrossRef]
- Veracx, A.; Raoult, D. Biology and Genetics of Human Head and Body Lice. Trends Parasitol. 2012, 28, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.N.; Eddy, G.W. Techniques for Rearing and Handling Body Lice, Oriental Rat Fleas, and Cat Fleas. Bull. World Health Organ. 1954, 10, 127. [Google Scholar]
- Takano-lee, M.; Yoon, K.S.; Edman, J.D.; Mullens, B.A.; Clark, J.M. In Vivo and In Vitro Rearing of Pediculus Humanus Capitis (Anoplura: Pediculidae). J. Med. Entomol. 2003, 40, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Culpepper, G.H. Rearing and Maintaining a Laboratory Colony of Body Lice on Rabbits. Am. J. Trop. Med. 1948, 28, 499–504. [Google Scholar] [CrossRef]
- Culpepper, G.H. Rearing Body Lice on Rabbits. J. Econ. Entomol. 1946, 39, 660. [Google Scholar] [CrossRef]
- Kim, J.H.; Min, J.S.; Kang, J.S.; Kwon, D.H.; Yoon, K.S.; Strycharz, J.; Koh, Y.H.; Pittendrigh, B.R.; Clark, J.M.; Lee, S.H. Comparison of the Humoral and Cellular Immune Responses between Body and Head Lice Following Bacterial Challenge. Insect Biochem. Mol. Biol. 2011, 41, 332–339. [Google Scholar] [CrossRef]
- Ben-Yakir, D.; Mumcuoglu, K.Y.; Manor, O.; Ochanda, J.; Galun, R. Immunization of Rabbits with a Midgut Extract of the Human Body Louse Pediculus Humanus Humanus: The Effect of Induced Resistance on the Louse Population. Med. Vet. Entomol. 1994, 8, 114–118. [Google Scholar] [CrossRef]
- Bland, D.M.; Hinnebusch, B.J. Feeding Behavior Modulates Biofilm-Mediated Transmission of Yersinia Pestis by the Cat Flea, Ctenocephalides Felis. PLoS Neglected Trop. Dis. 2016, 10, e0004413. [Google Scholar] [CrossRef]
- Fournier, P.-E.; Minnick, M.F.; Lepidi, H.; Salvo, E.; Raoult, D. Experimental Model of Human Body Louse Infection Using Green Fluorescent Protein-Expressing Bartonella Quintana. Infect. Immun. 2001, 69, 1876–1879. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, A.; Mougabure Cueto, G.; Picollo, M.I. Pediculus Humanus Capitis (Head Lice) and Pediculus Humanus Humanus (Body Lice): Response to Laboratory Temperature and Humidity and Susceptibility to Monoterpenoids. Parasitol. Res. 2009, 105, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Houhamdi, L.; Lepidi, H.; Drancourt, M.; Raoult, D. Experimental Model to Evaluate the Human Body Louse as a Vector of Plague. J. Infect. Dis. 2006, 194, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Houhamdi, L.; Fournier, P.-E.; Fang, R.; Lepidi, H.; Raoult, D. An Experimental Model of Human Body Louse Infection with Rickettsia Prowazekii. J. Infect. Dis. 2002, 186, 1639–1646. [Google Scholar] [CrossRef] [PubMed]
- Mumcuoglu, K.Y.; Miller, J.; Rosen, L.J.; Galun, R. Systemic Activity of Ivermectin on the Human Body Louse (Anoplura: Pediculidae). J. Med. Entomol. 1990, 27, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Amanzougaghene, N.; Fenollar, F.; Nappez, C.; Ben-Amara, A.; Decloquement, P.; Azza, S.; Bechah, Y.; Chabrière, E.; Raoult, D.; Mediannikov, O. Complexin in Ivermectin Resistance in Body Lice. PLoS Genet. 2018, 14, e1007569. [Google Scholar] [CrossRef]
- Kasai, S.; Mihara, M.; Takahashi, M.; Agui, N.; Tomita, T. An Artificial Blood Feeding System for Body Louse, Pediculus Humanus. Med. Entomol. Zool. 2003, 54, 343–351. [Google Scholar] [CrossRef]
- Haddon, W., Jr. An Artificial Membrane and Apparatus for the Feeding of the Human Body Louse Pediculus Humanus Corporis. Am. J. Trop. Med. Hyg. 1956, 5, 315–325. [Google Scholar] [CrossRef]
- Mumcuoglu, K.Y.; Danilevich, M.; Zelig, O.; Grinbaum, H.; Friger, M.; Meinking, T.L. Effects of Blood Type and Blood Handling on Feeding Success, Longevity and Egg Production in the Body Louse, Pediculus Humanus Humanus. Med. Vet. Entomol. 2011, 25, 12–16. [Google Scholar] [CrossRef]
- Haddon, W., Jr. The Maintenance of the Human Body Louse Pediculus Humanus Corporis through Complete Cycles of Growth by Serial Feeding through Artificial Membranes. Am. J. Trop. Med. Hyg. 1956, 5, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Sangare, A.K.; Rolain, J.M.; Gaudart, J.; Weber, P.; Raoult, D. Synergistic Activity of Antibiotics Combined with Ivermectin to Kill Body Lice. Int. J. Antimicrob. Agents 2016, 47, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Tsurukawa, C.; Kawada, H. Experiment on Mosquito Blood Feeding Using the Artificial Feeding Device. Med. Entomol. Zool. 2014, 65, 151–155. [Google Scholar] [CrossRef]
- Takano-Lee, M.; Velten, R.K.; Edman, J.D.; Mullens, B.A.; Clark, J.M. An Automated Feeding Apparatus for In Vitro Maintenance of the Human Head Louse, Pediculus Capiti s (Anoplura: Pediculidae). J. Med. Entomol. 2003, 40, 795–799. [Google Scholar] [CrossRef]
- Yoon, K.S.; Strycharz, J.P.; Gao, J.-R.; Takano-Lee, M.; Edman, J.D.; Clark, J.M. An Improved in Vitro Rearing System for the Human Head Louse Allows the Determination of Resistance to Formulated Pediculicides. Pestic. Biochem. Physiol. 2006, 86, 195–202. [Google Scholar] [CrossRef]
- Sonnberg, S.; Oliveira, F.A.; Araujo de Melo, I.L.; de Melo Soares, M.M.; Becher, H.; Heukelbach, J. Ex Vivo Development of Eggs from Head Lice (Pediculus Humanus Capitis). Open Dermatol. J. 2010, 4, 82–89. [Google Scholar] [CrossRef]
- Callan, M.B.; Shofer, F.S.; Catalfamo, J.L. Effects of Anticoagulant on pH, Ionized Calcium Concentration, and Agonist-Induced Platelet Aggregation in Canine Platelet-Rich Plasma. Am. J. Vet. Res. 2009, 70, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Eitzman, D.T.; Chi, L.; Saggin, L.; Schwartz, R.S.; Lucchesi, B.R.; Fay, W.P. Heparin Neutralization by Platelet-Rich Thrombi. Role of Platelet Factor 4. Circulation 1994, 89, 1523–1529. [Google Scholar] [CrossRef]
- Habedank, B.; Schrader, G.; Scheurer, S.; Schein, E. Investigations on the in Vitro Feeding and in Vitro Breeding of the Human Body Louse Pediculus Humanus (Anoplura: Pediculidae). In Proceedings of the 3rd International Conference on Urban Pests, Prague, Czech Republic, 19–22 July 1999; pp. 241–248. [Google Scholar]
- Sasaki-Fukatsu, K.; Koga, R.; Nikoh, N.; Yoshizawa, K.; Kasai, S.; Mihara, M.; Kobayashi, M.; Tomita, T.; Fukatsu, T. Symbiotic Bacteria Associated with Stomach Discs of Human Lice. Appl. Environ. Microbiol. 2006, 72, 7349–7352. [Google Scholar] [CrossRef]
- Hutchinson, R.H. A Note on the Life Cycle and Fertility of the Body Louse (Pediculus Corporis). J. Econ. Entomol. 1918, 11, 404–406. [Google Scholar] [CrossRef]
- Nuttall, G.H. The Biology of Pediculus Humanus. Parasitology 1917, 10, 80–185. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
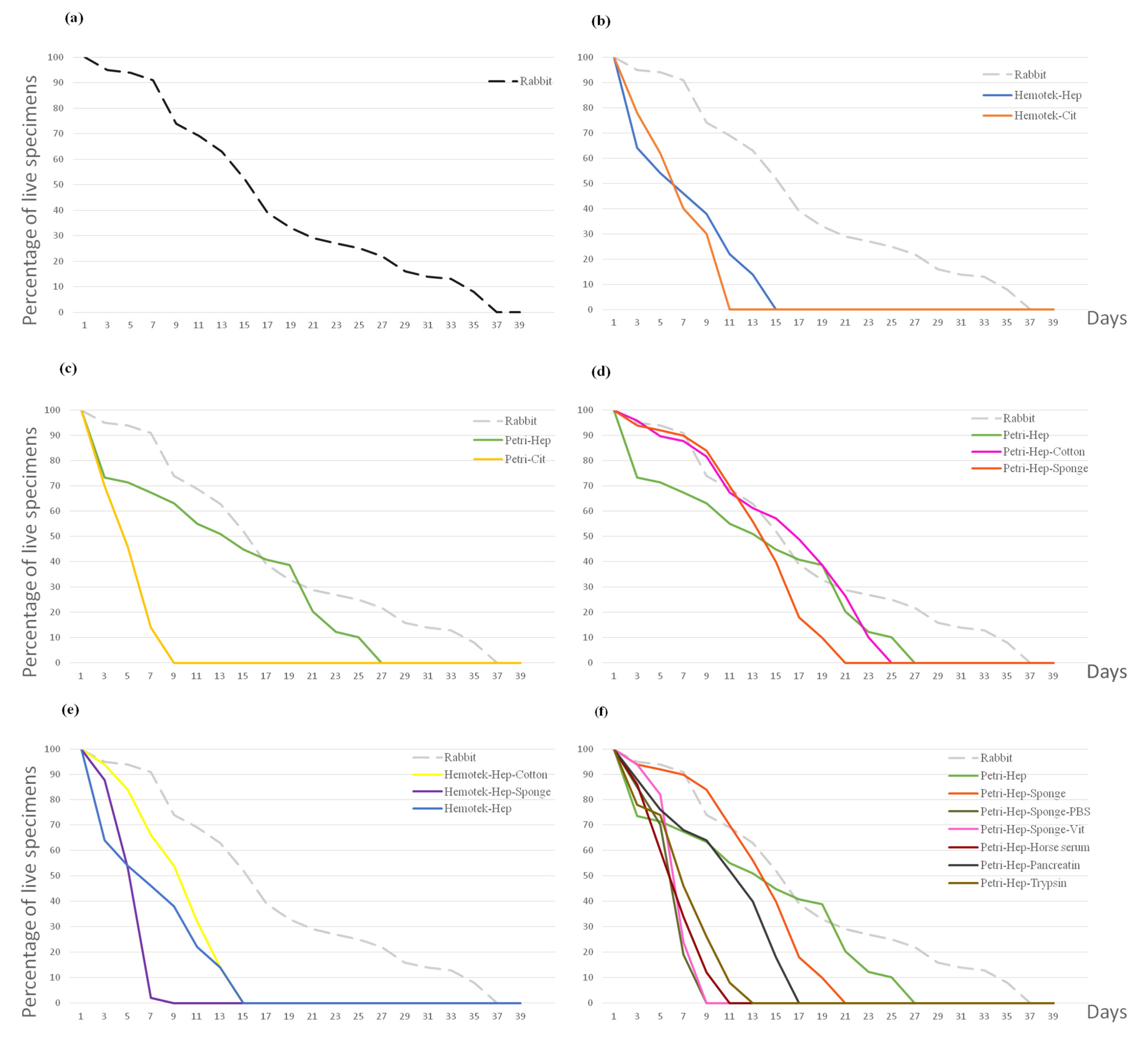
| System, Anticoagulant, Support | Blood Type/s | Days | 1 | 3 | 5 | 7 | 9 | 11 | 13 | 15 | 17 | 19 | 21 | 23 | 25 | 27 | 29 | 31 | 33 | 35 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rabbit host | Live | 100 | 95 | 94 | 91 | 74 | 69 | 63 | 52 | 39 | 33 | 29 | 27 | 25 | 22 | 16 | 14 | 13 | 8 | |
| Exuviae | 0 | 0 | 0 | 52 | 33 | 22 | 18 | 17 | 15 | 5 | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | ||
| Feeding % | 98 | 95 | 96 | 97 | 97 | 92 | 96 | 92 | 92 | 84 | 82 | 85 | 88 | 90 | 87 | 85 | 76 | 70 | ||
| Petri, Heparin | 2 O+ | Live | 500 | 444 | 414 | 399 | 341 | 289 | 265 | 240 | 221 | 191 | 169 | 138 | 115 | 94 | 34 | 22 | 12 | 4 |
| Exuviae | 0 | 0 | 0 | 15 | 250 | 40 | 11 | 155 | 24 | 7 | 73 | 30 | 15 | 0 | 0 | 0 | 0 | 0 | ||
| Feeding % | 96 | 95 | 95 | 93 | 93 | 93 | 91 | 89 | 94 | 89 | 92 | 90 | 86 | 74 | 64 | 31 | 8 | |||
| Petri, Heparin | 2 AB+ | Live | 500 | 477 | 439 | 371 | 305 | 265 | 218 | 175 | 145 | 113 | 71 | 50 | 36 | 16 | 6 | |||
| Exuviae | 0 | 0 | 0 | 0 | 27 | 126 | 46 | 34 | 47 | 8 | 2 | 5 | 1 | 0 | 0 | |||||
| Feeding % | 95 | 95 | 98 | 98 | 98 | 94 | 92 | 95 | 94 | 82 | 72 | 78 | 62 | 50 | ||||||
| Petri, Heparin | 2 O+, O− | Live | 49 | 36 | 35 | 33 | 31 | 27 | 25 | 22 | 20 | 19 | 10 | 6 | 5 | |||||
| Exuviae | 0 | 0 | 0 | 8 | 20 | 0 | 0 | 16 | 5 | 0 | 4 | 2 | 0 | |||||||
| Feeding % | 87 | 94 | 100 | 93 | 90 | 92 | 100 | 90 | 95 | 89 | 70 | 66 | 40 | |||||||
| Petri, Heparin, Cotton | 2 O+, O− | Live | 49 | 47 | 44 | 43 | 40 | 33 | 30 | 28 | 24 | 19 | 13 | 5 | ||||||
| Exuviae | 0 | 0 | 0 | 13 | 23 | 1 | 0 | 20 | 8 | 0 | 7 | 0 | ||||||||
| Feeding % | 87 | 95 | 100 | 97 | 92 | 90 | 96 | 89 | 95 | 94 | 84 | 80 | ||||||||
| Petri, Heparin, Sponge | O+ | Live | 50 | 47 | 46 | 45 | 42 | 35 | 28 | 20 | 9 | 5 | ||||||||
| Exuviae | 0 | 0 | 0 | 10 | 22 | 0 | 0 | 16 | 3 | 0 | ||||||||||
| Feeding % | 96 | 95 | 95 | 95 | 91 | 89 | 85 | 77 | 90 | 60 | ||||||||||
| Hemotek, Heparin | O+ | Live | 50 | 32 | 27 | 23 | 19 | 11 | 7 | First-stage larvae | ||||||||||
| Exuviae | 0 | 0 | 0 | 0 | 14 | 7 | 0 | Second-stage larvae | ||||||||||||
| Feeding % | 98 | 93 | 92 | 82 | 78 | 72 | 57 | Third-stage larvae | ||||||||||||
| Petri, Citrate | A+ | Live | 50 | 35 | 23 | 7 | Adult | |||||||||||||
| Exuviae | 0 | 0 | 0 | 0 | ||||||||||||||||
| Feeding % | 98 | 88 | 78 | |||||||||||||||||
| Hemotek, Citrate | A+ | Live | 50 | 39 | 31 | 20 | 15 | |||||||||||||
| Exuviae | 0 | 0 | 0 | 0 | 0 | |||||||||||||||
| Feeding % | 61 | 80 | 55 | 66 | 60 | |||||||||||||||
| Condition | Petri Dish with Heparinised Blood Cohort: 500 | Petri Dish with Heparinised Blood Cohort: 500 | Petri Dish with Heparinised Blood Cohort: 50 | Rabbit Host Cohort: 100 |
|---|---|---|---|---|
| Engorgement | 81% | 86% | 85% | 89% |
| Longevity in days | 35 | 29 | 25 | 35 |
| First-stage larvae | 6 | 8 | 6 | 6 |
| Second-stage larvae | 3 | 6 | 8 | 4 |
| Third-stage larvae | 6 | 6 | 6 | 6 |
| Adult | 19 | 9 | 5 | 19 |
| No. of eggs/F | 0.45 | 0 | 0 | 1.8 |
| Egg hatch | 43.6% | 0 | 0 | 79% |
| Systems | ||||
|---|---|---|---|---|
| Materials | Rabbit Host | Petri Dish (Six Vessels) | Hemotek System (Six Vessels) | |
| Lice maintenance | Incubator *** | 0.085 | 0.085 | 0.085 |
| Cloth * | 0.32 | 0.32 | 0.32 | |
| Pots * | 0.23 | 0.23 | 0.23 | |
| Rearing | Heating pad ** | 0.107 | NU | NU |
| Shaving machine ** | 0.641 | NU | NU | |
| Syringes * | 0.1 | NU | NU | |
| Anaesthetising product * | Rompun 3.462 | NU | NU | |
| Imalgene 2.534 | NU | NU | ||
| Vacuum cleaner *** | 0.04 | NU | NU | |
| Rabbits ** | 6.41 | NU | NU | |
| Cages *** | 0.341 | NU | NU | |
| Absorbent sheet * | 2.4 | NU | NU | |
| Water bath ** | NU | 0.213 | 0.213 | |
| Heating plate ** | NU | 0.427 | NU | |
| Hemotek system *** | NU | NU | 1.068 | |
| Petri dish * | NU | 0.349 | NU | |
| Blood * | NU | 5.28 | 5.28 | |
| Fridge *** | 0.128 | 0.12 | 0.12 | |
| Parafilm * | NU | 0.11 | 0.057 | |
| Technician equipment | Technician salary | 30.194 | 15.097 | 15.097 |
| Disposable lab gowns* | 0.48 | 0.48 | 0.48 | |
| Gloves * | 0.08 | 0.08 | 0.08 | |
| Lab coat * | 0.18 | 0.18 | 0.18 | |
| Hair net * | 0.02 | 0.02 | 0.02 | |
| Over socks * | 0.12 | NU | NU | |
| Technician training *** | 0.512 | NU | NU | |
| Post-rearing | Floor and cage cleaning * | 0.066 | NU | NU |
| Rabbit food * | 2.721 | NU | NU | |
| Rabbit maintenance * | 0.705 | NU | NU | |
| Surfasafe * | 0.014 | 0.014 | 1.125 | |
| Bleach * | NU | NU | 0.22 | |
| Total | 51.894€ | 23.007€ | 24.577€ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hammoud, A.; Louni, M.; Abou-Chacra, L.; Haddad, G.; Mazzotti, N.; Fenollar, F.; Mediannikov, O. Development of a Technique Using Artificial Membrane for In Vitro Rearing of Body Lice Pediculus humanus humanus. Insects 2024, 15, 145. https://doi.org/10.3390/insects15030145
Hammoud A, Louni M, Abou-Chacra L, Haddad G, Mazzotti N, Fenollar F, Mediannikov O. Development of a Technique Using Artificial Membrane for In Vitro Rearing of Body Lice Pediculus humanus humanus. Insects. 2024; 15(3):145. https://doi.org/10.3390/insects15030145
Chicago/Turabian StyleHammoud, Alissa, Meriem Louni, Linda Abou-Chacra, Gabriel Haddad, Noelle Mazzotti, Florence Fenollar, and Oleg Mediannikov. 2024. "Development of a Technique Using Artificial Membrane for In Vitro Rearing of Body Lice Pediculus humanus humanus" Insects 15, no. 3: 145. https://doi.org/10.3390/insects15030145
APA StyleHammoud, A., Louni, M., Abou-Chacra, L., Haddad, G., Mazzotti, N., Fenollar, F., & Mediannikov, O. (2024). Development of a Technique Using Artificial Membrane for In Vitro Rearing of Body Lice Pediculus humanus humanus. Insects, 15(3), 145. https://doi.org/10.3390/insects15030145