

Hypervolume Niche Dynamics and Global Invasion Risk of Phenacoccus solenopsis under Climate Change

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Sources
2.1.1. Species Occurrence Data
2.1.2. Bioclimatic Variables
2.2. Niche Estimation
2.3. MaxEnt Modeling
2.3.1. Model Optimization
2.3.2. Model Establishment
2.3.3. Model Evaluation
3. Results
3.1. PCA of Climatic Variables
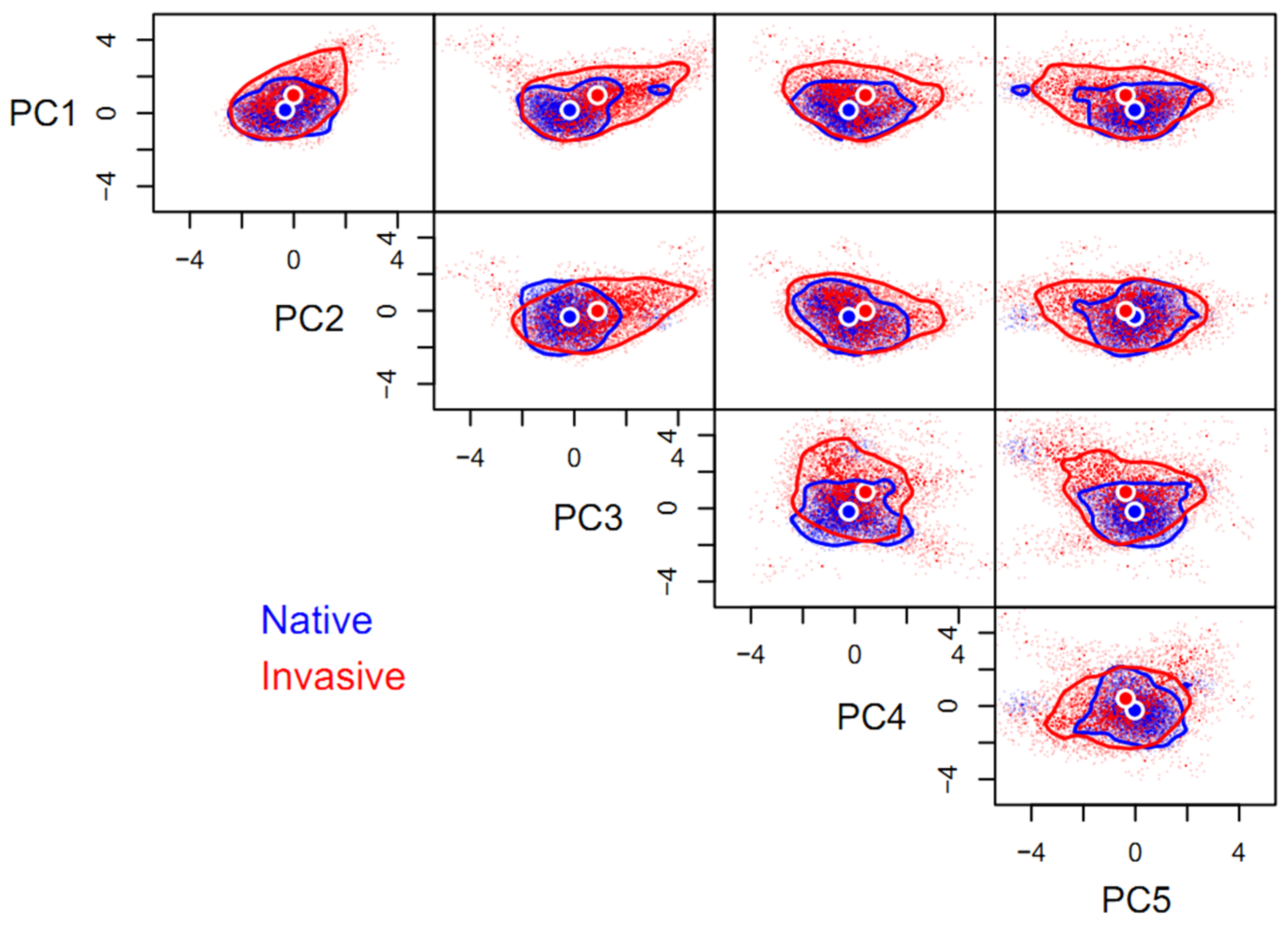
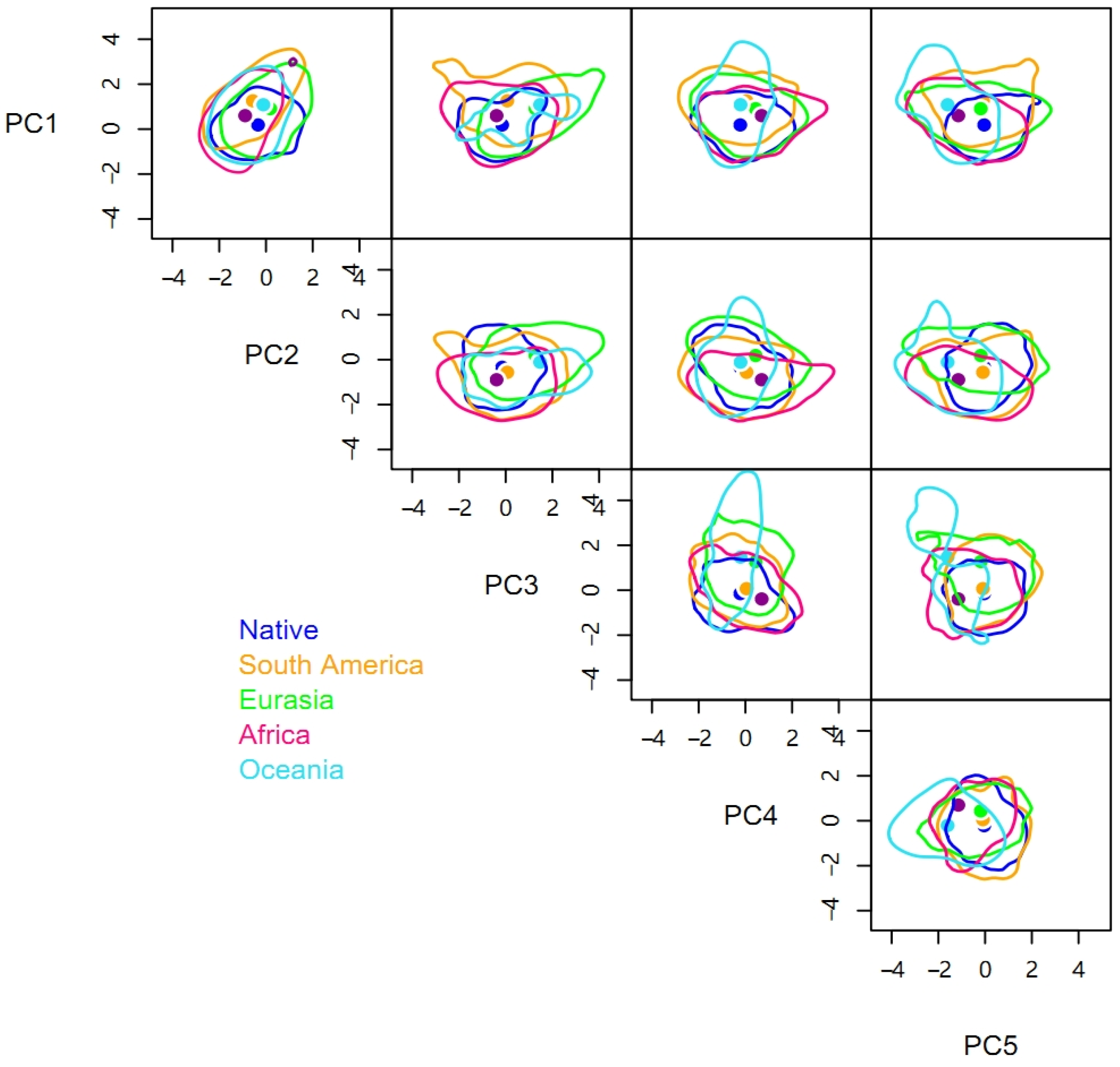
3.2. Niche Differences between Native and Invasive Ranges
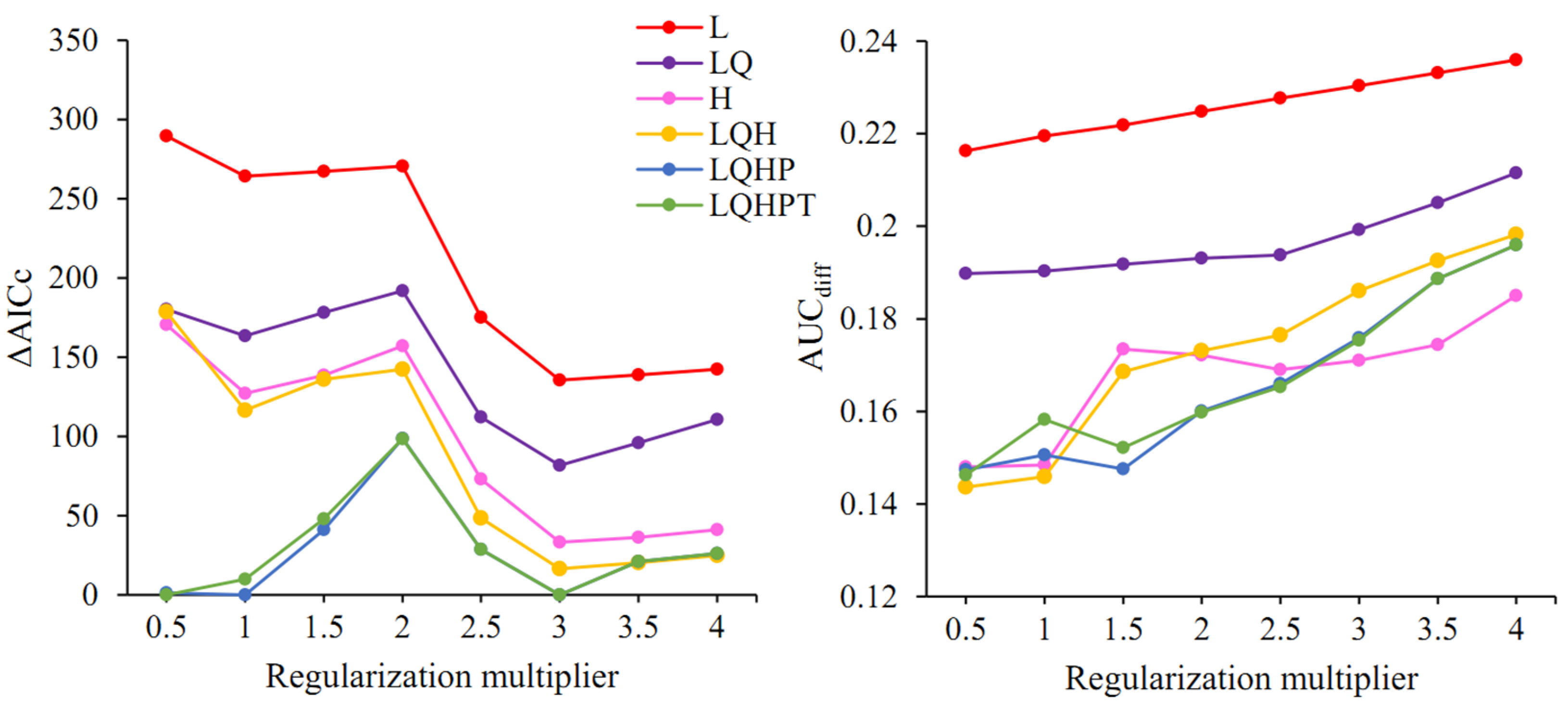
3.3. Model Optimization and Evaluation
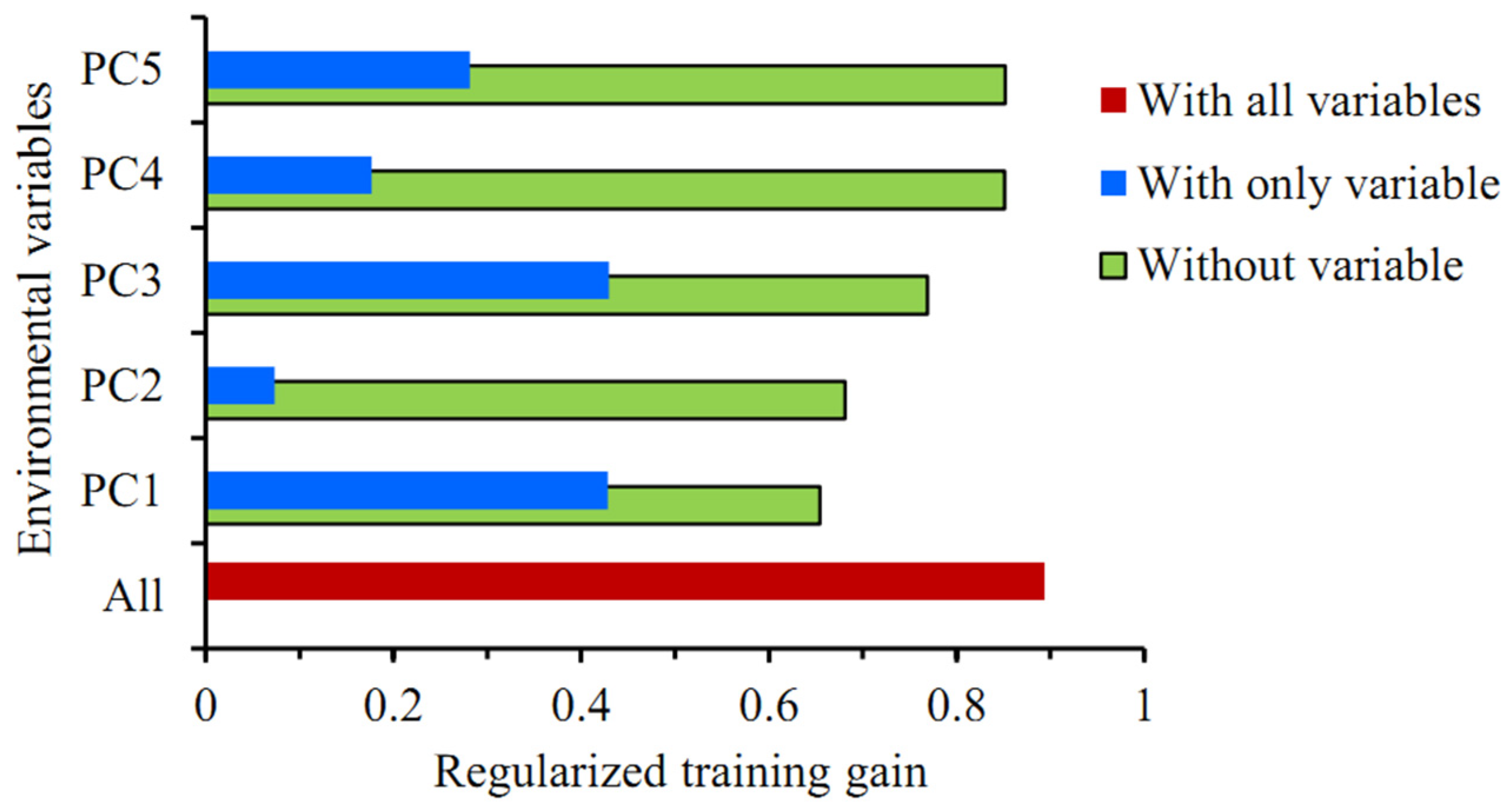
3.4. Importance of Variables
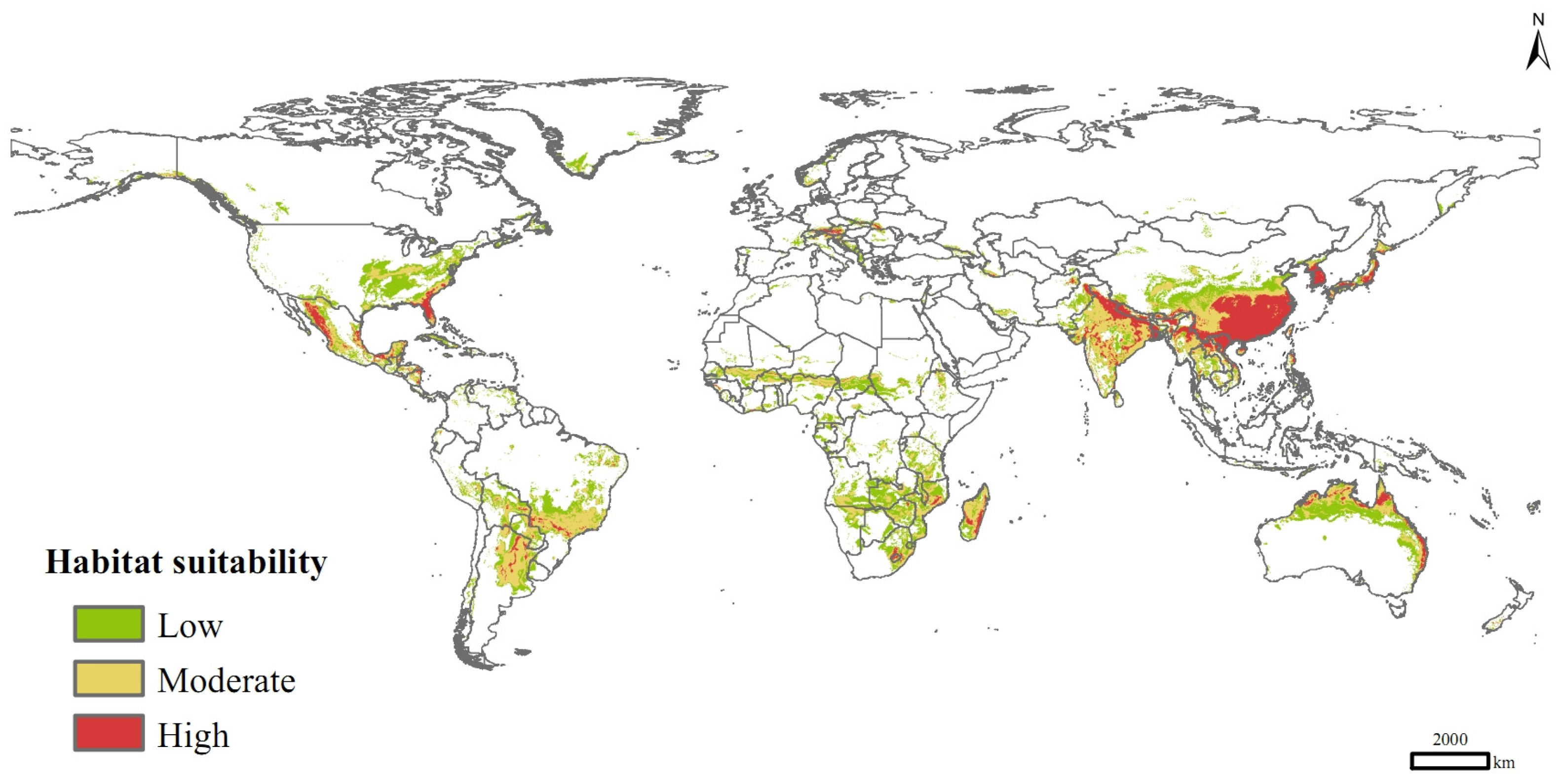
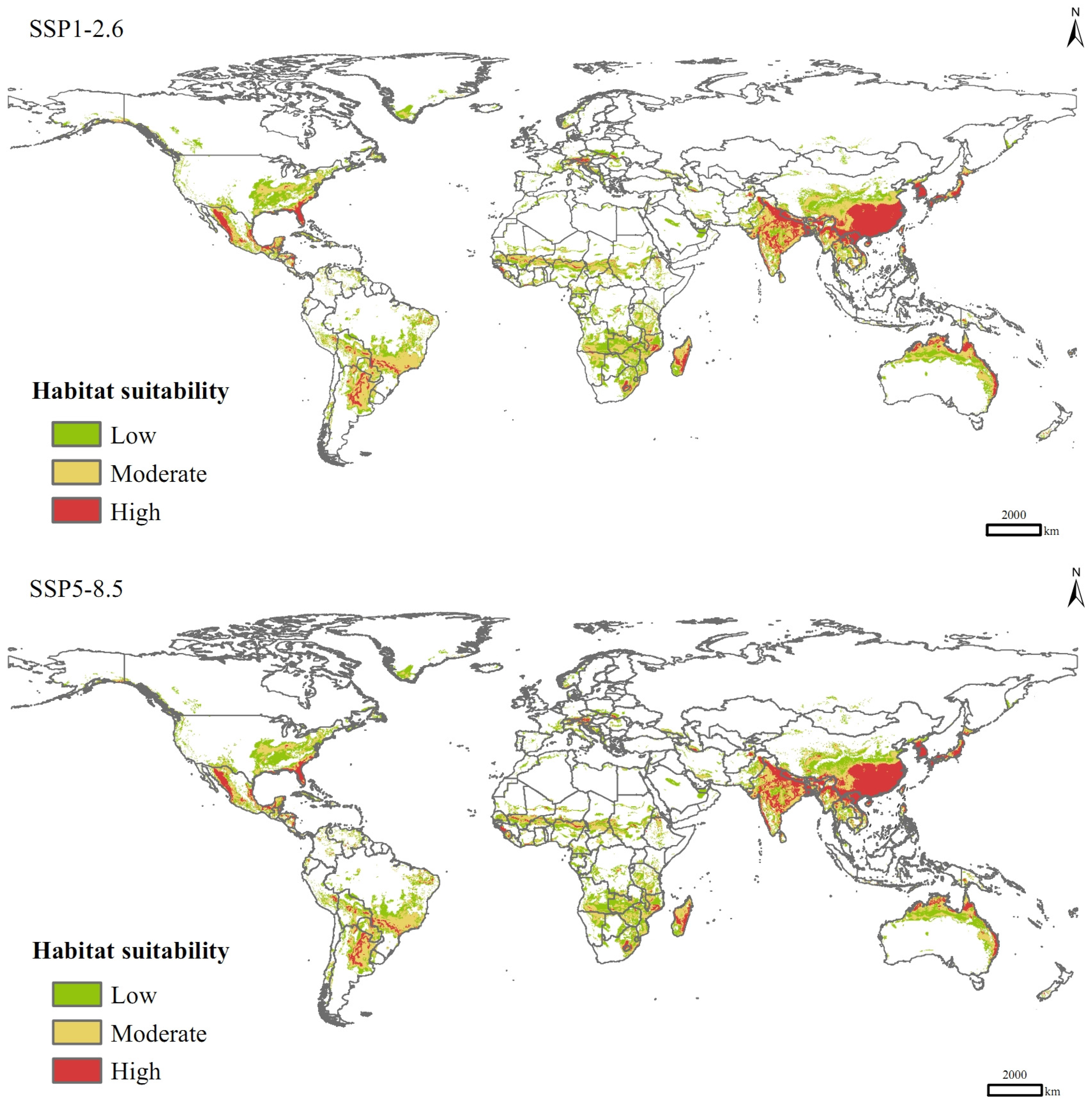
3.5. Potential Distribution in Current and Future Climate Scenarios
4. Discussion
4.1. Niche Shifts
4.2. SDM Construction
4.3. Invasion Risks and Management Recommendations
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, C.; Wolter, C.; Xian, W.; Jeschke, J.M. Most invasive species largely conserve their climatic niche. Proc. Natl. Acad. Sci. USA 2020, 117, 23643–23651. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wolter, C.; Courchamp, F.; Roura-Pascual, N.; Jeschke, J.M. Biological invasions reveal how niche change affects the transferability of species distribution models. Ecology 2022, 103, e3719. [Google Scholar] [CrossRef]
- Bates, O.K.; Bertelsmeier, C. Climatic niche shifts in introduced species. Curr. Biol. 2021, 31, R1252–R1266. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.P.; Gallardo, B.; Terblanche, J.S. A global assessment of climatic niche shifts and human influence in insect invasions. Glob. Ecol. Biogeogr. 2017, 26, 679–689. [Google Scholar] [CrossRef]
- Atwater, D.Z.; Barney, J.N. Climatic niche shifts in 815 introduced plant species affect their predicted distributions. Glob. Ecol. Biogeogr. 2021, 30, 1671–1684. [Google Scholar] [CrossRef]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1348. [Google Scholar] [CrossRef] [PubMed]
- Blonder, B.; Lamanna, C.; Violle, C.; Enquist, B.J. The n-dimensional hypervolume. Glob. Ecol. Biogeogr. 2014, 23, 595–609. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding remarks. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Blonder, B.; Morrow, C.B.; Maitner, B.; Harris, D.J.; Lamanna, C.; Violle, C.; Enquist, B.J.; Kerkhoff, A.J. New approaches for delineating n-dimensional hypervolumes. Methods Ecol. Evol. 2018, 9, 305–319. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Cardoso, P. Decomposing the causes for niche differentiation between species using hypervolumes. Front. Ecol. Evol. 2020, 8, 243. [Google Scholar] [CrossRef]
- Mammola, S.; Cardoso, P. Functional diversity metrics using kernel density n-dimensional hypervolumes. Methods Ecol. Evol. 2020, 11, 986–995. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, H.; Zhao, W.; Zhao, Q. Niche shifts and the potential distribution of Phenacoccus solenopsis (Hemiptera: Pseudococcidae) under climate change. PLoS ONE 2017, 12, e0180913. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Gao, X.; Hu, X.; Hou, Y.; Wang, F. Current and future potential distribution of the invasive scale Ceroplastes rusci (L., 1758) (Hemiptera: Coccidae) under climate niche. Pest Manag. Sci. 2023, 79, 1184–1192. [Google Scholar] [CrossRef] [PubMed]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Warren, D.L.; Wright, A.N.; Seifert, S.N.; Shaffer, H.B. Incorporating model complexity and spatial sampling bias into ecological niche models of climate change risks faced by 90 California vertebrate species of concern. Divers. Distrib. 2014, 20, 334–343. [Google Scholar] [CrossRef]
- Zhu, G.; Qiao, H. Effect of the Maxent model’s complexity on the prediction of species potential distributions. Biodivers. Sci. 2016, 24, 1189–1196. [Google Scholar] [CrossRef]
- Tinsley, J.D. An ants’-nest coccid from New Mexico. Can. Entomol. 1898, 30, 47–48. [Google Scholar] [CrossRef]
- García Morales, M.; Denno, B.D.; Miller, D.R.; Miller, G.L.; Ben-Dov, Y.; Hardy, N.B. ScaleNet: A literature-based model of scale insect biology and systematics. Database 2016, 2016, bav118. [Google Scholar]
- Waqas, M.S.; Shi, Z.; Yi, T.C.; Xiao, R.; Shoaib, A.A.; Elabasy, A.S.; Jin, D.C. Biology, ecology, and management of cotton mealybug Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae). Pest Manag. Sci. 2021, 77, 5321–5333. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S. Distribution Pattern and Genetic Structure of Phenacoccus Solenopsis Tinsley and Distribution Pattern of Its Parasitoid Wasps in China. Ph.D. Dissertation, Chinese Academy of Agricultural Sciences, Beijing, China, 2019. [Google Scholar]
- Zhou, A.; Lu, Y.; Zeng, L.; Xu, Y.; Liang, G. Effects of honeydew of Phenacoccus solenopsis on foliar foraging by Solenopsis invcta (Hymenoptera: Formicidae). Sociobiology 2012, 59, 71–79. [Google Scholar]
- Nagrare, V.S.; Kranthi, S.; Biradar, V.K.; Zade, N.; Sangode, V.; Kakde, G.; Shukla, R.M.; Shivare, D.; Khadi, B.M.; Kranthi, K.R. Widespread infestation of the exotic mealybug species, Phenacoccus solenopsis (Tinsley) (Hemiptera: Pseudococcidae), on cotton in India. Bull. Entomol. Res. 2009, 99, 537–541. [Google Scholar] [CrossRef]
- Culik, M.P.; Gullan, P.J. A new pest of tomato and other records of mealybugs (Hemiptera: Pseudococcidae) from Espirito Santo, Brazil. Zootaxa 2005, 964, 1–8. [Google Scholar] [CrossRef]
- Larraín, S. Incidencia de insectos y ácaros plagas en pepino dulce (Solanum muricatum Ait.) cultivado en la IV Región, Chile. Agric. Técnica 2002, 62, 15–26. [Google Scholar] [CrossRef]
- Granara de Willink, M.; Szumik, C. Central and south American Phenacoccinae (Hemiptera: Coccoidea: Pseudococcidae): Systematics and phylogeny. Rev. Soc. Entomológica Argent. 2007, 66, 29–129. [Google Scholar]
- Abbas, G.; Arif, M.J.; Saeed, S. Systematic status of a new species of genus Phenacoccus cockerell (Pseudococcidae), a serious pest of cotton, Gossypium hirsutum L., in Pakistan. Pak. Entomol. 2005, 27, 83–84. [Google Scholar]
- Fuchs, T.W.; Stewart, J.W.; Minzenmayer, R.; Rose, M. First record of Phenacoccus solenopsis Tinsley in cultivated cotton in the United States. Southwest. Entomol. 1991, 16, 215–221. [Google Scholar]
- Charleston, K.; Addison, S.; Miles, M.; Maas, S. The Solenopsis mealybug outbreak in Emerald. Aust. Cottongrower 2010, 31, 18–22. [Google Scholar]
- Akintola, A.J.; Ande, A.T. First record of Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) on Hibiscus rosa-sinensis in Nigeria. Agric. J. 2008, 3, 1–3. [Google Scholar]
- Zhang, H.; Xu, L.; Lyu, W.; Zhou, Y.; Wang, W.; Gao, R.; Cui, S.; Zhang, Z. Multidimensional climatic niche conservatism and invasion risk of Phenacoccus solenopsis. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2023, 34, 1649–1658. [Google Scholar]
- Wang, Y.S.; Dai, T.M.; Hu, T.; Wan, F.H.; Zhang, G. Range expansion of the invasive cotton mealybug, Phenacoccus solenopsis Tinsley: An increasing threat to agricultural and horticultural crops in China. J. Integr. Agric. 2020, 19, 881–885. [Google Scholar] [CrossRef]
- Xu, H.G.; Qiang, S. China’s Invasive Alien Species, revised ed.; Science Press: Beijing, China, 2018; pp. 815–818. [Google Scholar]
- Gebregergis, Z. Incidence of a new pest, the cotton mealybug Phenacoccus solenopsis Tinsley, on sesame in North Ethiopia. Int. J. Zool. 2018, 2018, 3531495. [Google Scholar] [CrossRef]
- Tong, H.; Yan, A.O.; Li, Z.; Wang, Y.; Jiang, M. Invasion biology of the cotton mealybug, Phenacoccus solenopsis Tinsley: Current knowledge and future directions. J. Integr. Agric. 2019, 18, 758–770. [Google Scholar] [CrossRef]
- Aroua, K.; Kaydan, M.B.; Ercan, C.; Biche, M. First Record of Phenacoccus solenopsis Tinsley (Hemiptera: Coccoidea: Pseudococcidae) in Algeria. Entomol. News 2020, 129, 63–66. [Google Scholar] [CrossRef]
- Ricupero, M.; Biondi, A.; Russo, A.; Zappalà, L.; Mazzeo, G. The cotton mealybug is spreading along the Mediterranean: First pest detection in Italian Tomatoes. Insects 2021, 12, 675. [Google Scholar] [CrossRef] [PubMed]
- Macharia, I.; Kibwage, P.; Heya, H.M.; Makathima, F.; Olubayo, D.; Guantai, M.; Kinuthia, W.; Ouvrard, D.; Watson, G.W. New records of scale insects and mealybugs (Hemiptera: Coccomorpha) in Kenya. EPPO Bull. 2021, 51, 639–647. [Google Scholar] [CrossRef]
- El Aalaoui, M.; Sbaghi, M. First record of the mealybug Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) and its seven parasitoids and five predators in Morocco. EPPO Bull. 2021, 51, 299–304. [Google Scholar] [CrossRef]
- Bader, A.K.; Al-Jboory, I.J. First record of cotton mealybug, Phenacoccus solenopsis Tinsley 1898 (Hemiptera: Pseudococcidae), from Saudi Arabia. EPPO Bull. 2020, 50, 557–560. [Google Scholar] [CrossRef]
- Abbes, K.; Harbi, A.; Wanassi, T.; Ricupero, M.; Mazzeo, G.; Russo, A.; Biondi, A.; Zappalà, L.; Chermiti, B. The cotton mealybug Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) is spreading in North Africa: First report in Tunisia. Orient. Insects 2023, 58, 1–15. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Kaiser, H.F. Coefficient alpha for a principal component and the Kaiser-Guttman rule. Psychol. Rep. 1991, 68, 855–858. [Google Scholar] [CrossRef]
- Pack, K.E.; Mieszkowska, N.; Rius, M. Rapid niche shifts as drivers for the spread of a non-indigenous species under novel environmental conditions. Divers. Distrib. 2022, 28, 596–610. [Google Scholar] [CrossRef]
- Hypervolume: High Dimensional Geometry, Set Operations, Projection, and Inference Using Kernel Density Estimation, Support Vector Machines, and Convex Hulls. Available online: https://CRAN.R-project.org/package=hypervolume (accessed on 3 January 2024).
- Kass, J.M.; Muscarella, R.; Galante, P.J.; Bohl, C.L.; Pinilla-Buitrago, G.E.; Boria, R.A.; Soley-Guardia, M.; Anderson, R.P. ENMeval 2.0: Redesigned for customizable and reproducible modeling of species’ niches and distributions. Methods Ecol. Evol. 2021, 12, 1602–1608. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; pp. 1–488. [Google Scholar]
- Cao, Y.; deWalt, R.E.; Robinson, J.L.; Tweddale, T.; Hinz, L.; Pessino, M. Using Maxent to model the historic distributions of stonefly species in Illinois streams: The effects of regularization and threshold selections. Ecol. Model. 2013, 259, 30–39. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Zhao, Y.; Deng, X.; Xiang, W.; Chen, L.; Ouyang, S. Predicting potential suitable habitats of Chinese fir under current and future climatic scenarios based on Maxent model. Ecol. Inform. 2021, 64, 101393. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Strubbe, D.; Broennimann, O.; Chiron, F.; Matthysen, E. Niche conservatism in non-native birds in Europe: Niche unfilling rather than niche expansion. Glob. Ecol. Biogeogr. 2013, 22, 962–970. [Google Scholar] [CrossRef]
- Early, R.; Sax, D.F. Climatic niche shifts between species’ native and naturalized ranges raise concern for ecological forecasts during invasions and climate change. Glob. Ecol. Biogeogr. 2014, 23, 1356–1365. [Google Scholar] [CrossRef]
- Guo, W.Y.; Lambertini, C.; Li, X.Z.; Meyerson, L.A.; Brix, H. Invasion of old world Phragmites australis in the new world: Precipitation and temperature patterns combined with human influences redesign the invasive niche. Glob. Chang. Biol. 2013, 19, 3406–3422. [Google Scholar] [CrossRef] [PubMed]
- Moya, W.; Jacome, G.; Yoo, C. Past, current, and future trends of red spiny lobster based on PCA with MaxEnt model in Galapagos Islands, Ecuador. Ecol. Evol. 2017, 7, 4881–4890. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Wang, F.; Zhu, Y.; Huang, F.; Lu, Y. Effects of temperature on the development and reproduction of Phenacoccus solenopsis (Hemiptera: Pseudococcidae). Acta Entomol. Sin. 2014, 57, 436–442. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PC | Current | 2081–2100 | |
|---|---|---|---|
| SSP1-2.6 | SSP5-8.5 | ||
| PC1 | 77.10 | 76.89 | 76.48 |
| PC2 | 17.45 | 16.81 | 17.16 |
| PC3 | 3.10 | 3.40 | 3.44 |
| PC4 | 1.59 | 2.00 | 1.99 |
| PC5 | 0.52 | 0.63 | 0.65 |
| Cumulative contribution | 99.75 | 99.73 | 99.73 |
| Native Range | South America | Eurasia | Africa | Oceania | |
|---|---|---|---|---|---|
| Native range | — | 1.19 | 1.79 | 1.76 | 2.54 |
| South America | 0.88 = 0.05 + 0.83 | — | 1.46 | 1.63 | 2.21 |
| Eurasia | 0.89 = 0.02 + 0.88 | 0.67 = 0.52 + 0.15 | — | 2.27 | 1.63 |
| Africa | 0.91 = 0.06 + 0.85 | 0.58 = 0.44 + 0.14 | 0.69 = 0.68 + 0.01 | — | 2.40 |
| Oceania | 0.94 = 0.02 + 0.92 | 0.75 = 0.33 + 0.42 | 0.71 = 0.41 + 0.30 | 0.64 = 0.32 + 0.32 | — |
| Hypervolume | 140.41 | 963.31 | 1197.40 | 1176.13 | 1917.27 |
| Habitat Suitability | Current | 2081–2100 | |
|---|---|---|---|
| SSP1-2.6 | SSP5-8.5 | ||
| Low | 1530 | 1800 (17.63%) | 1878 (22.70%) |
| Moderate | 1333 | 1777 (33.25%) | 1580 (18.48%) |
| High | 621 | 881 (41.93%) | 927 (49.25%) |
| Total | 3485 | 4458 (27.93%) | 4384 (25.82%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, S.; Zhang, H.; Liu, L.; Lyu, W.; Xu, L.; Zhang, Z.; Han, Y. Hypervolume Niche Dynamics and Global Invasion Risk of Phenacoccus solenopsis under Climate Change. Insects 2024, 15, 250. https://doi.org/10.3390/insects15040250
Cui S, Zhang H, Liu L, Lyu W, Xu L, Zhang Z, Han Y. Hypervolume Niche Dynamics and Global Invasion Risk of Phenacoccus solenopsis under Climate Change. Insects. 2024; 15(4):250. https://doi.org/10.3390/insects15040250
Chicago/Turabian StyleCui, Shaopeng, Huisheng Zhang, Lirui Liu, Weiwei Lyu, Lin Xu, Zhiwei Zhang, and Youzhi Han. 2024. "Hypervolume Niche Dynamics and Global Invasion Risk of Phenacoccus solenopsis under Climate Change" Insects 15, no. 4: 250. https://doi.org/10.3390/insects15040250
APA StyleCui, S., Zhang, H., Liu, L., Lyu, W., Xu, L., Zhang, Z., & Han, Y. (2024). Hypervolume Niche Dynamics and Global Invasion Risk of Phenacoccus solenopsis under Climate Change. Insects, 15(4), 250. https://doi.org/10.3390/insects15040250