
Expanding the Mesozoic Record of Early Brachyceran Fly Larvae, including New Larval Forms with Chimera-Type Morphologies

 ,
,  and
and 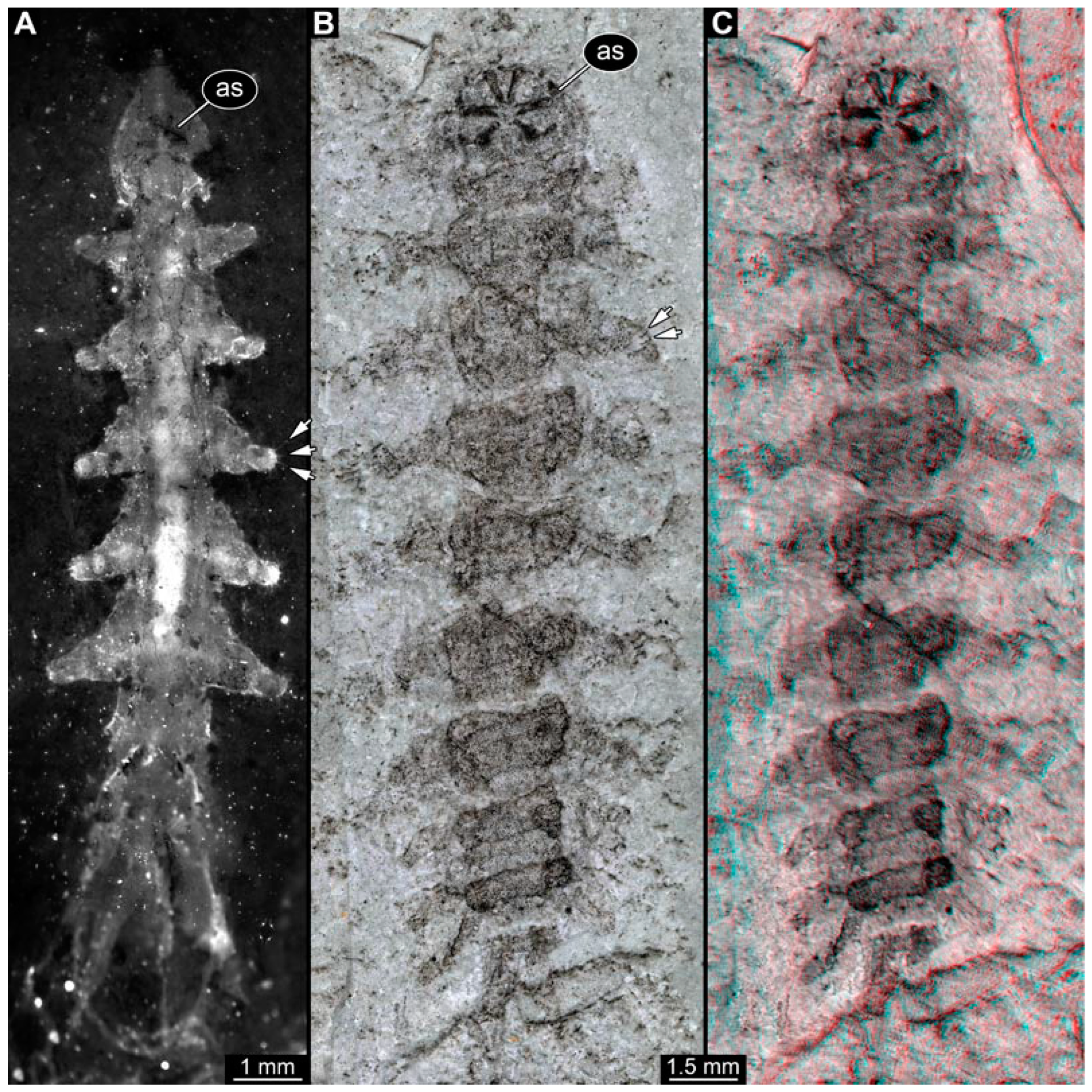{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Material
2.2. Documentation Methods
2.3. Measurements
3. Results
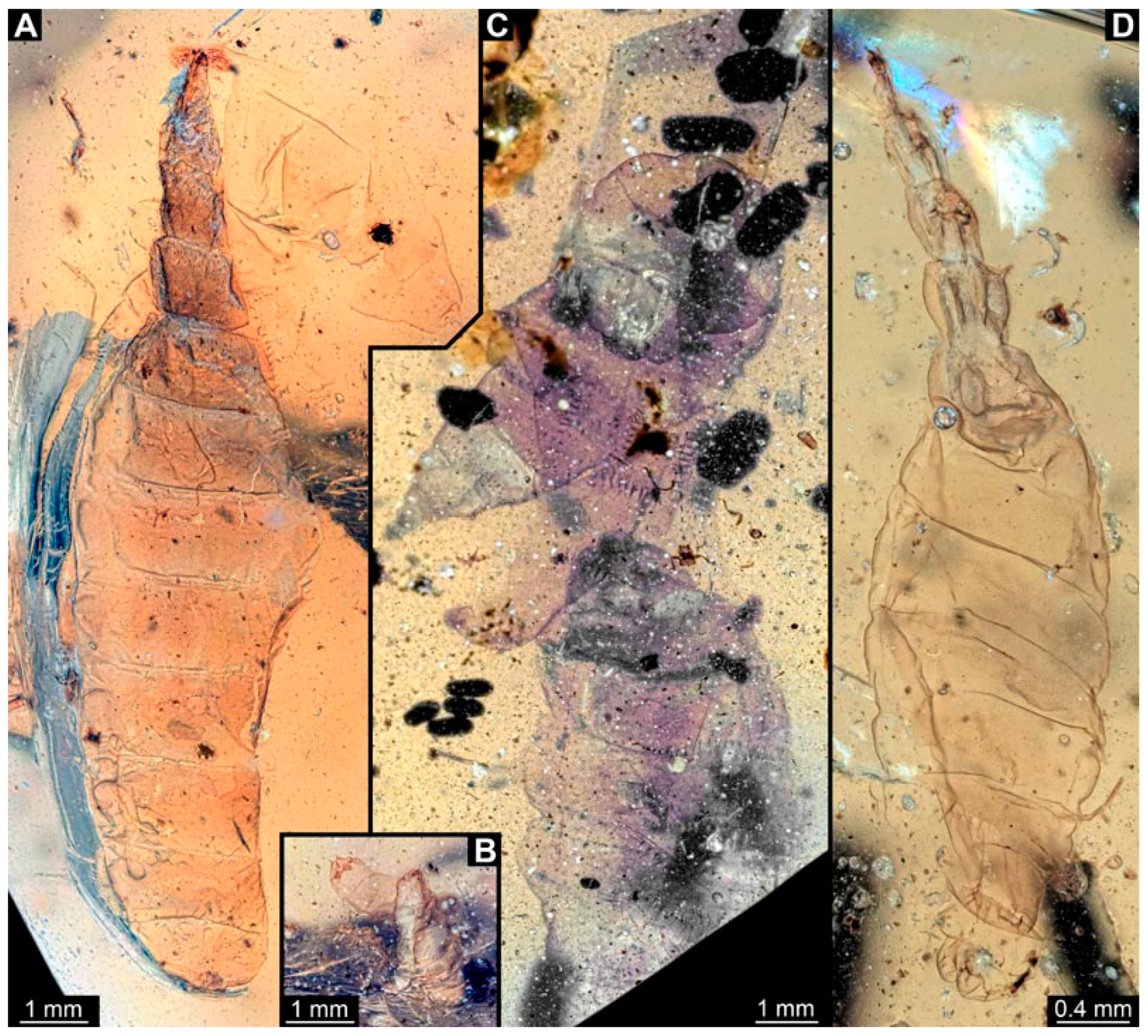
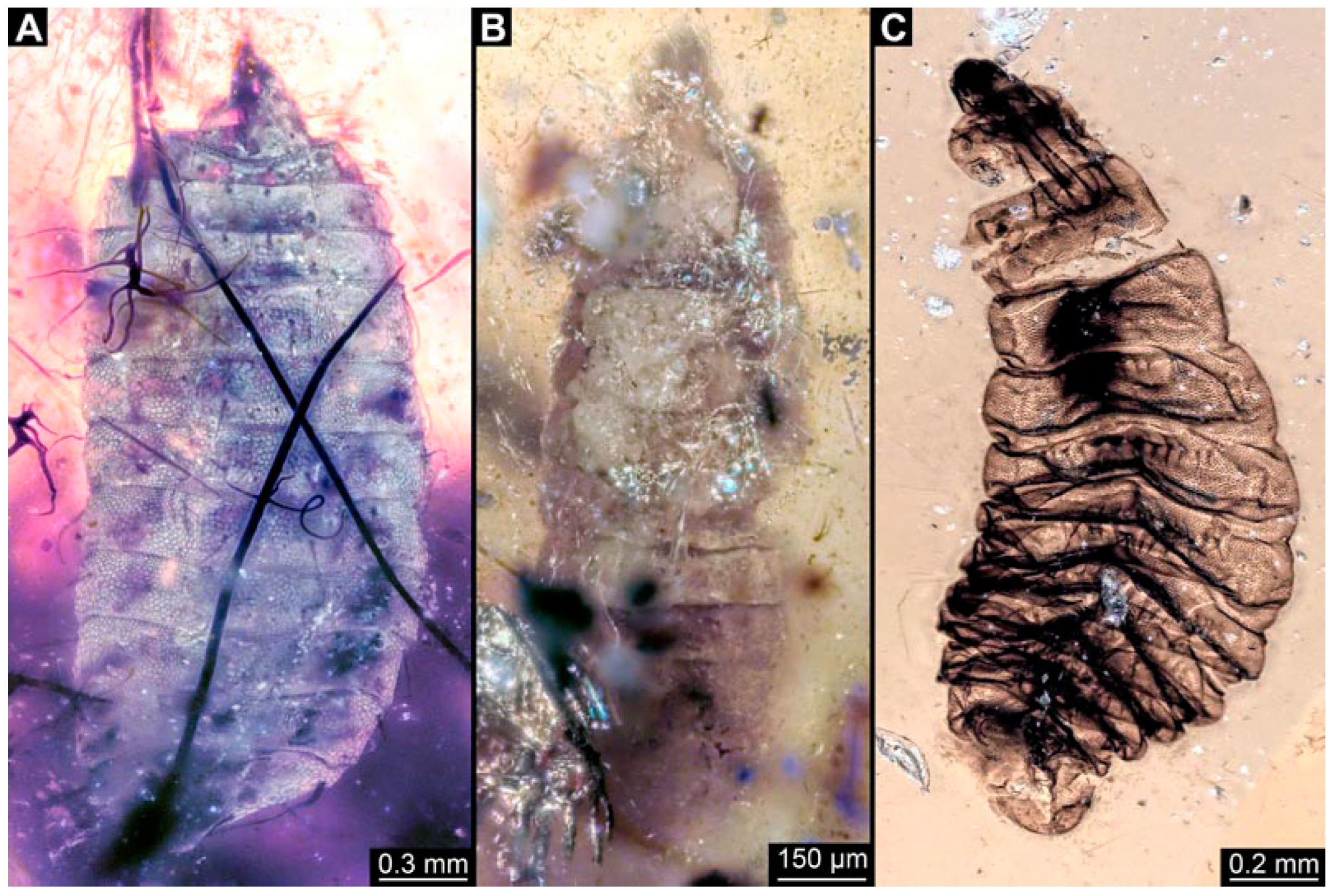
3.1. Short Descriptions of Specimens from Daohugou
3.2. Specimens of Stratiomyomorphan Morphotype 1 or Unclear Type
3.3. Specimens of Stratiomyomorphan Morphotype 2 or 7
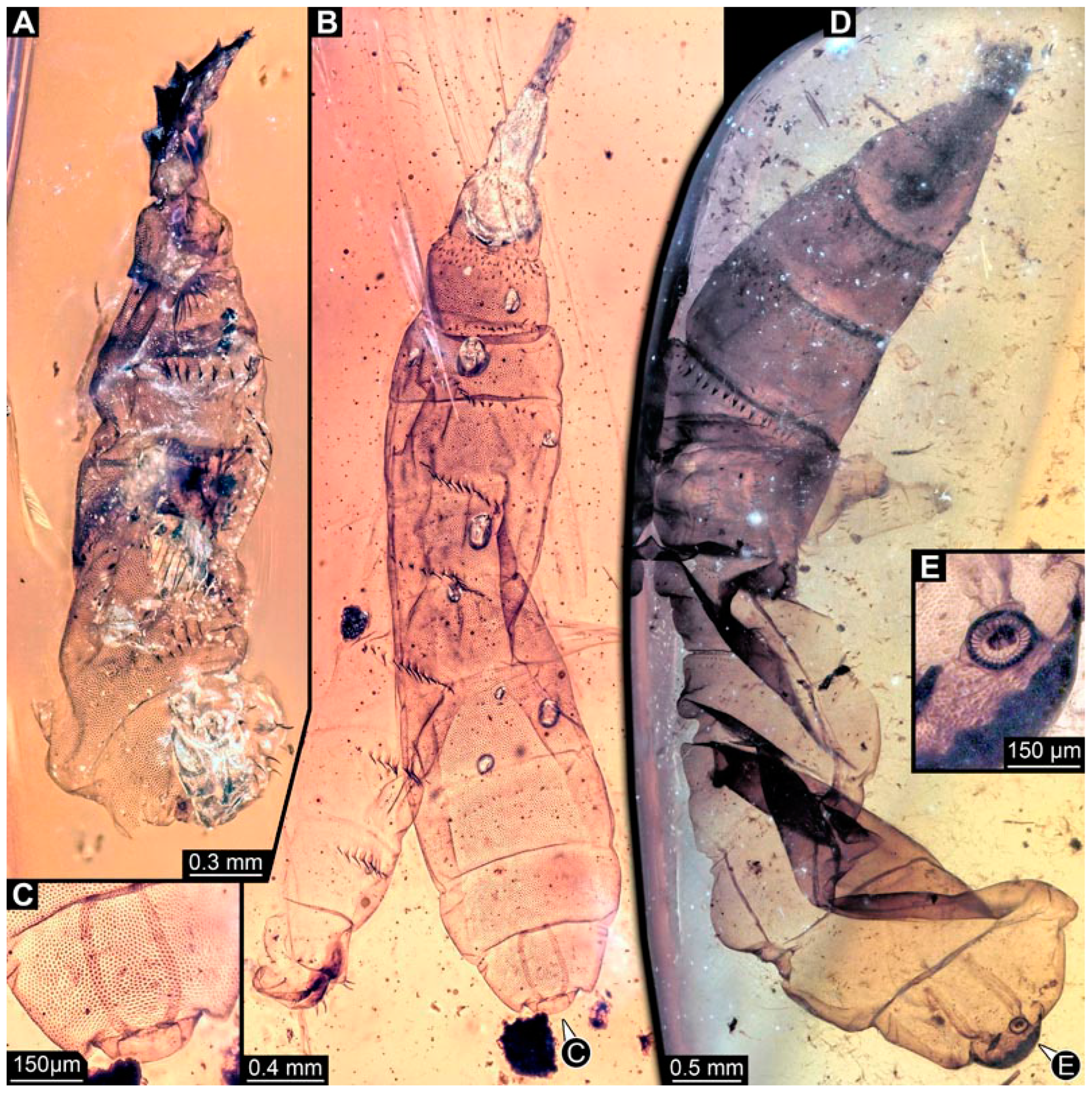
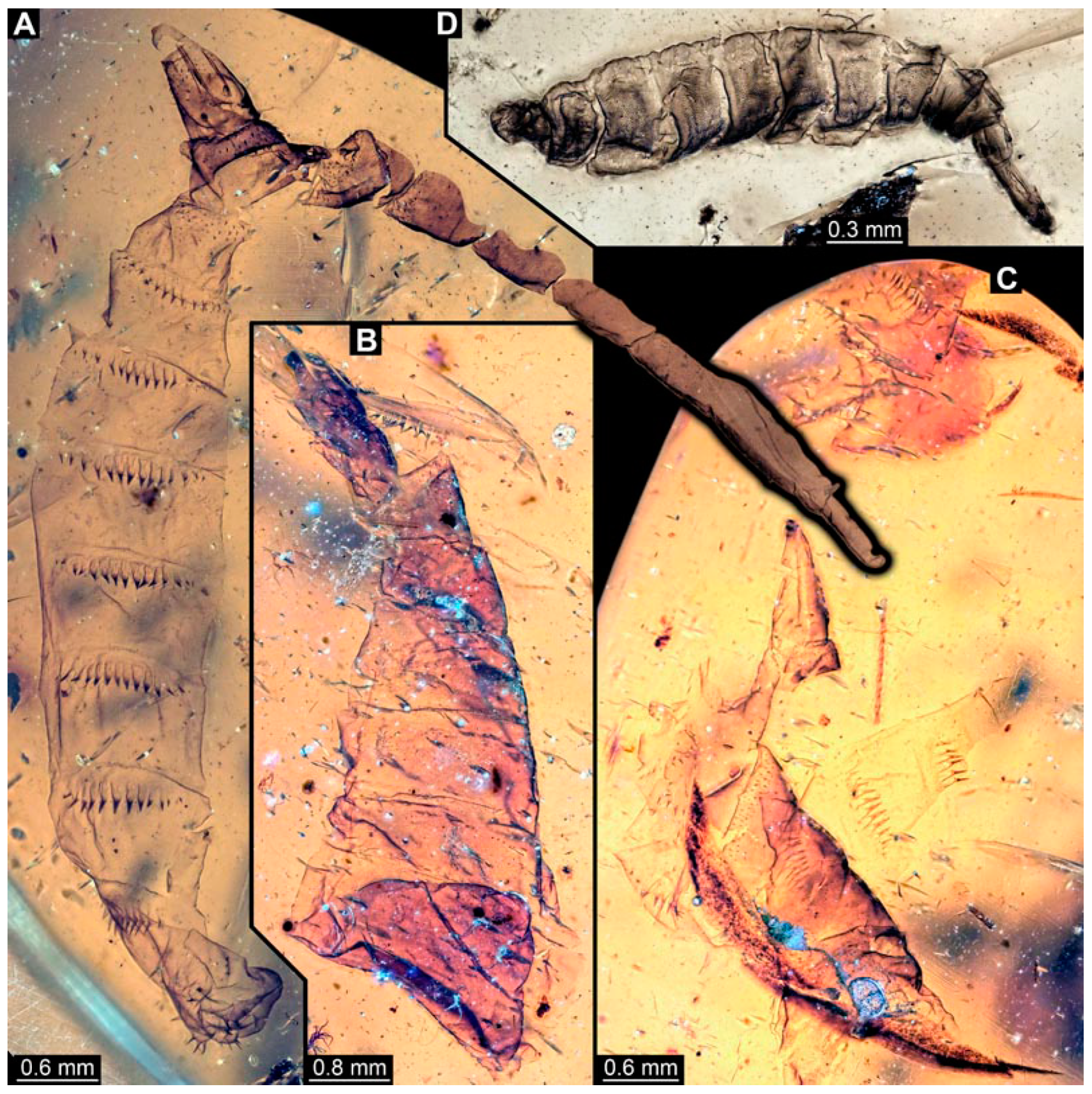
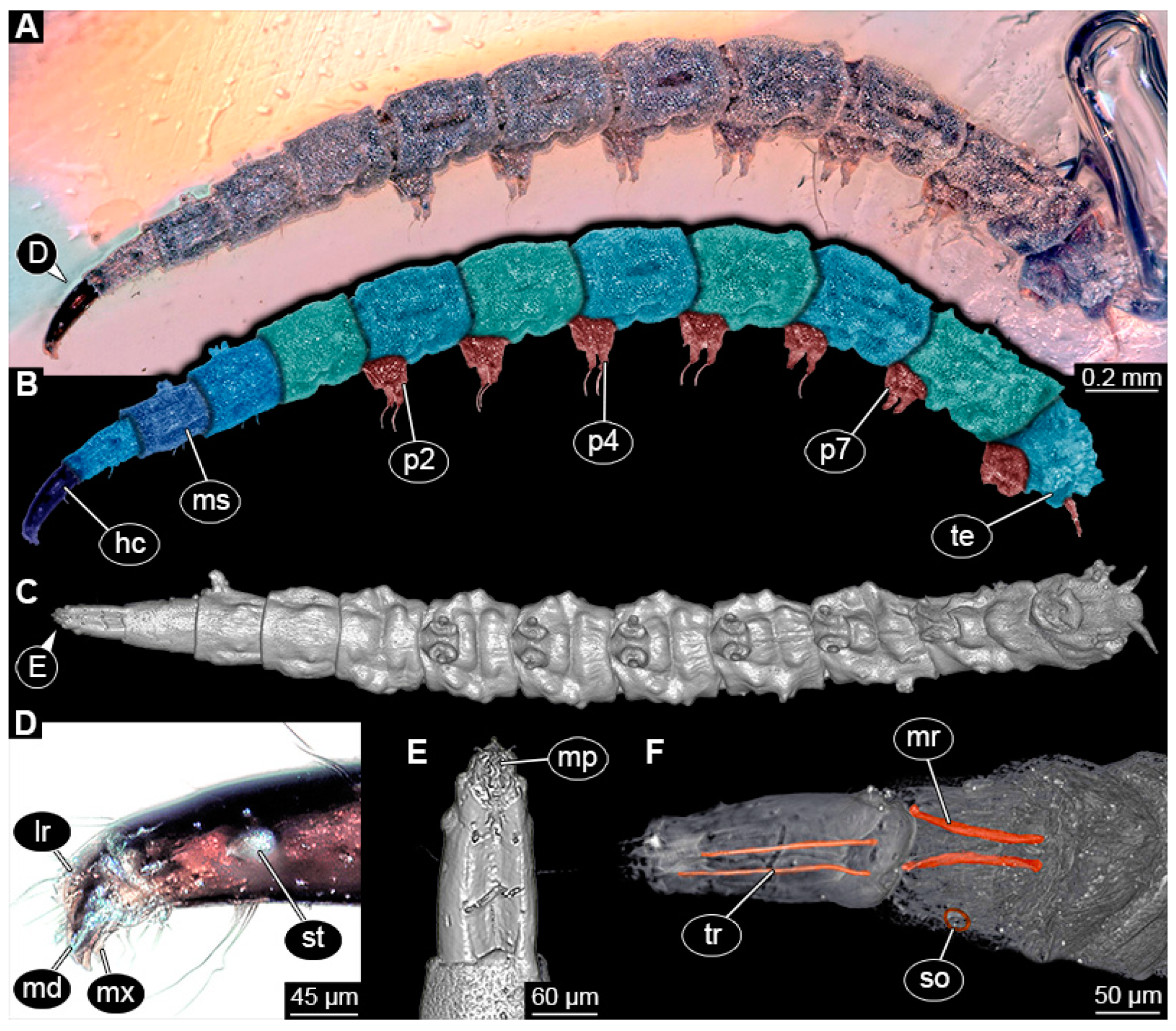
3.4. Short Descriptions of Specimens of Elongate Chimera Morphotype
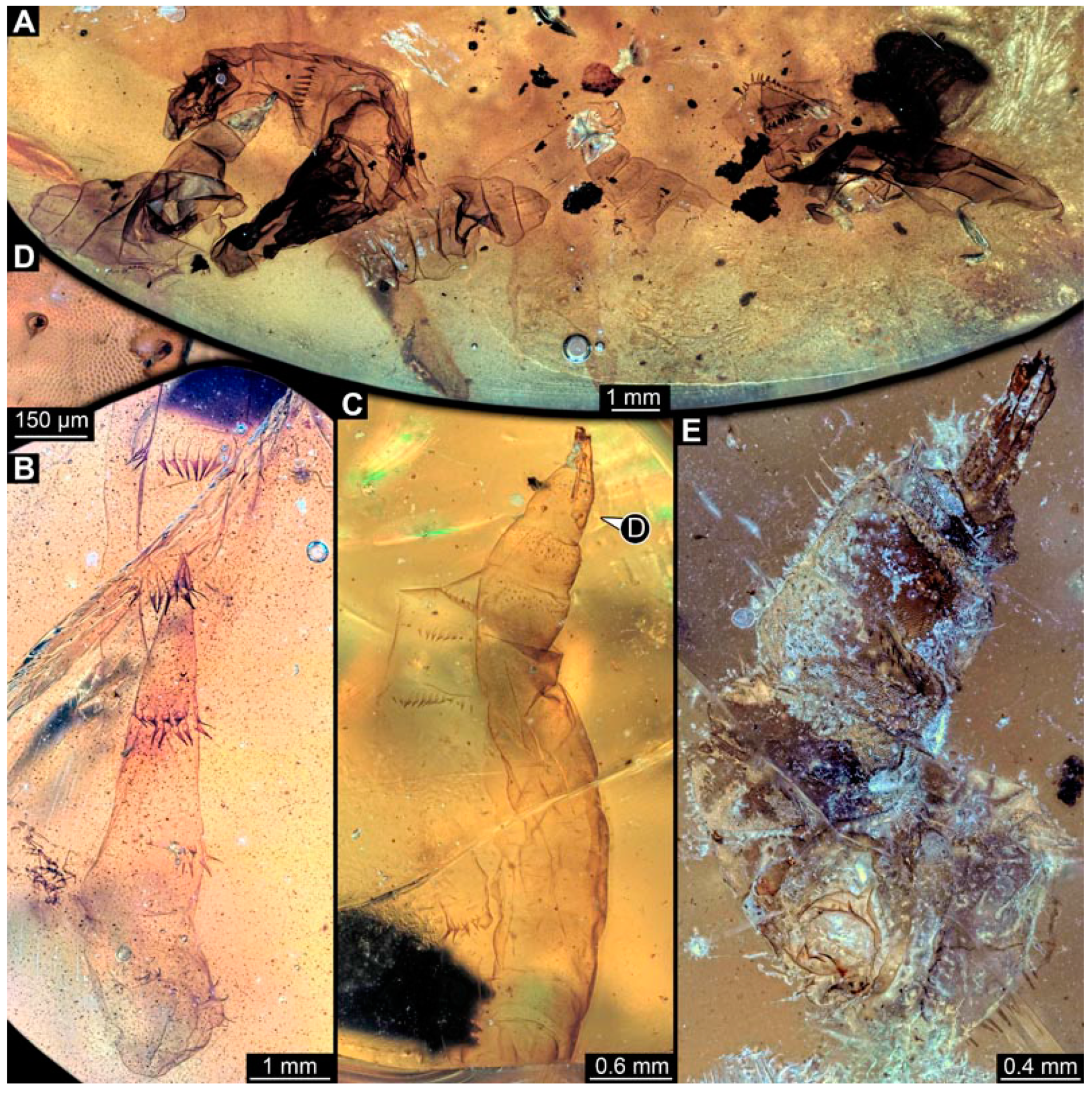
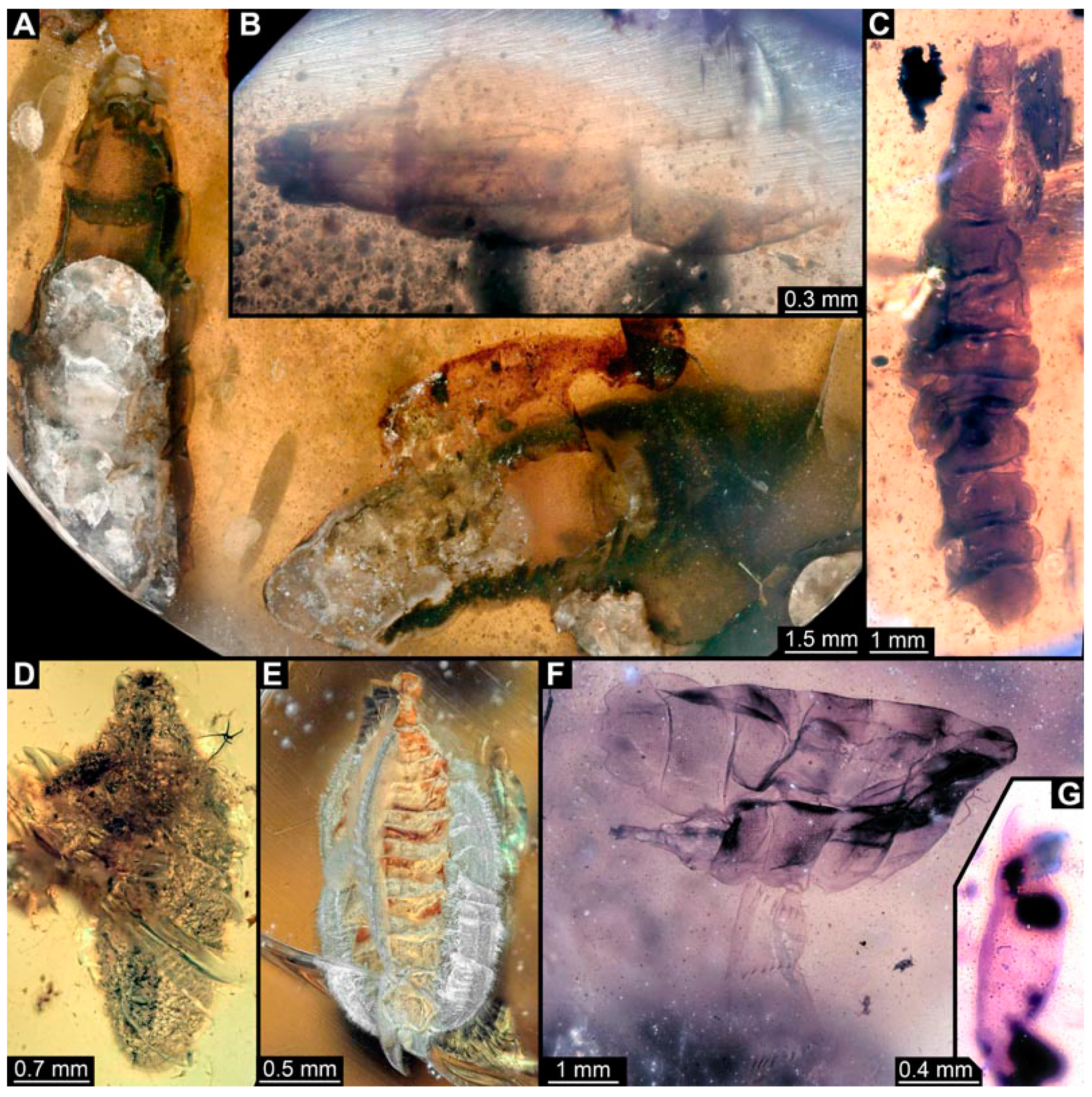
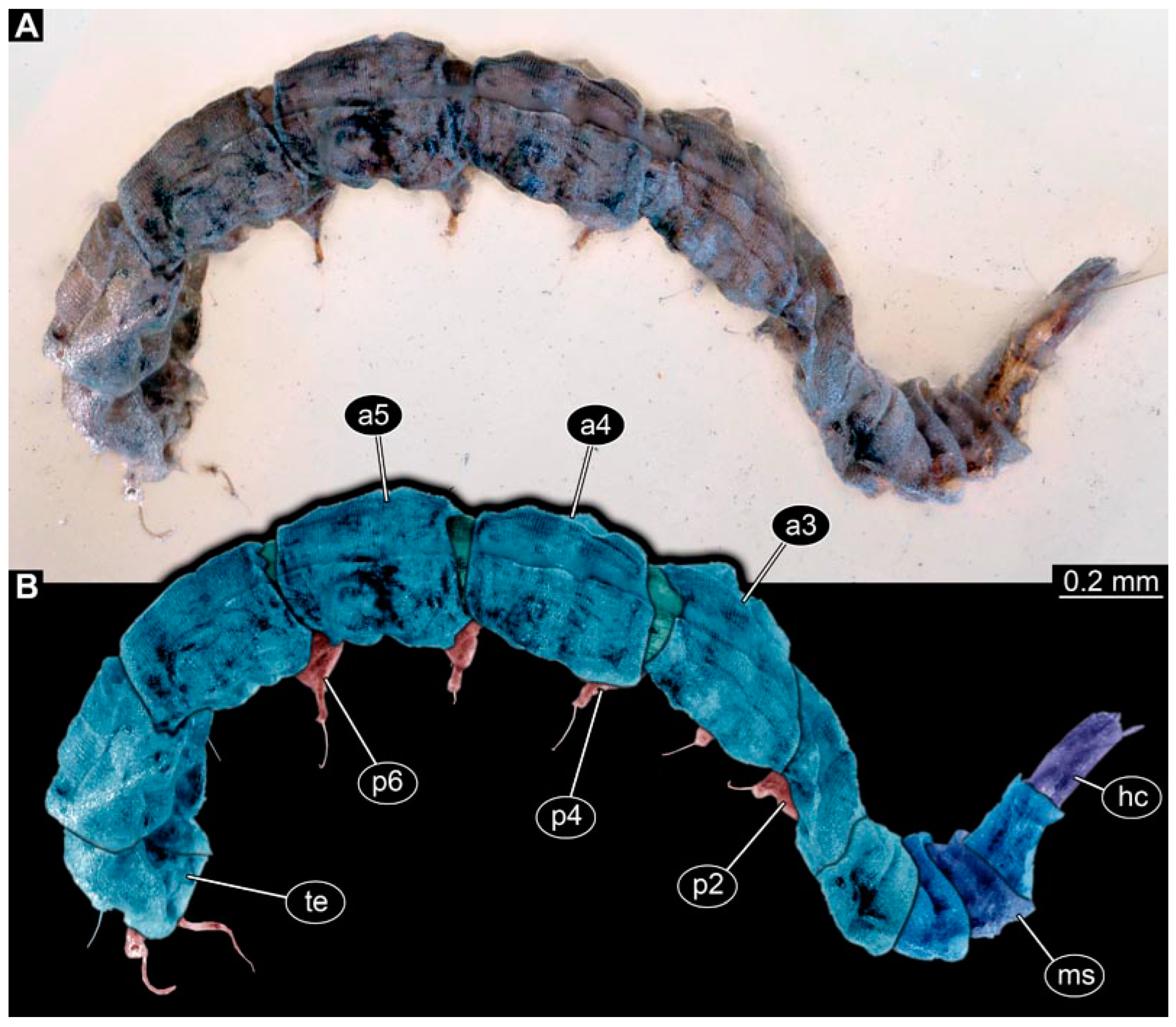
3.5. Short Descriptions of Specimen of Stouter Chimera Morphotype
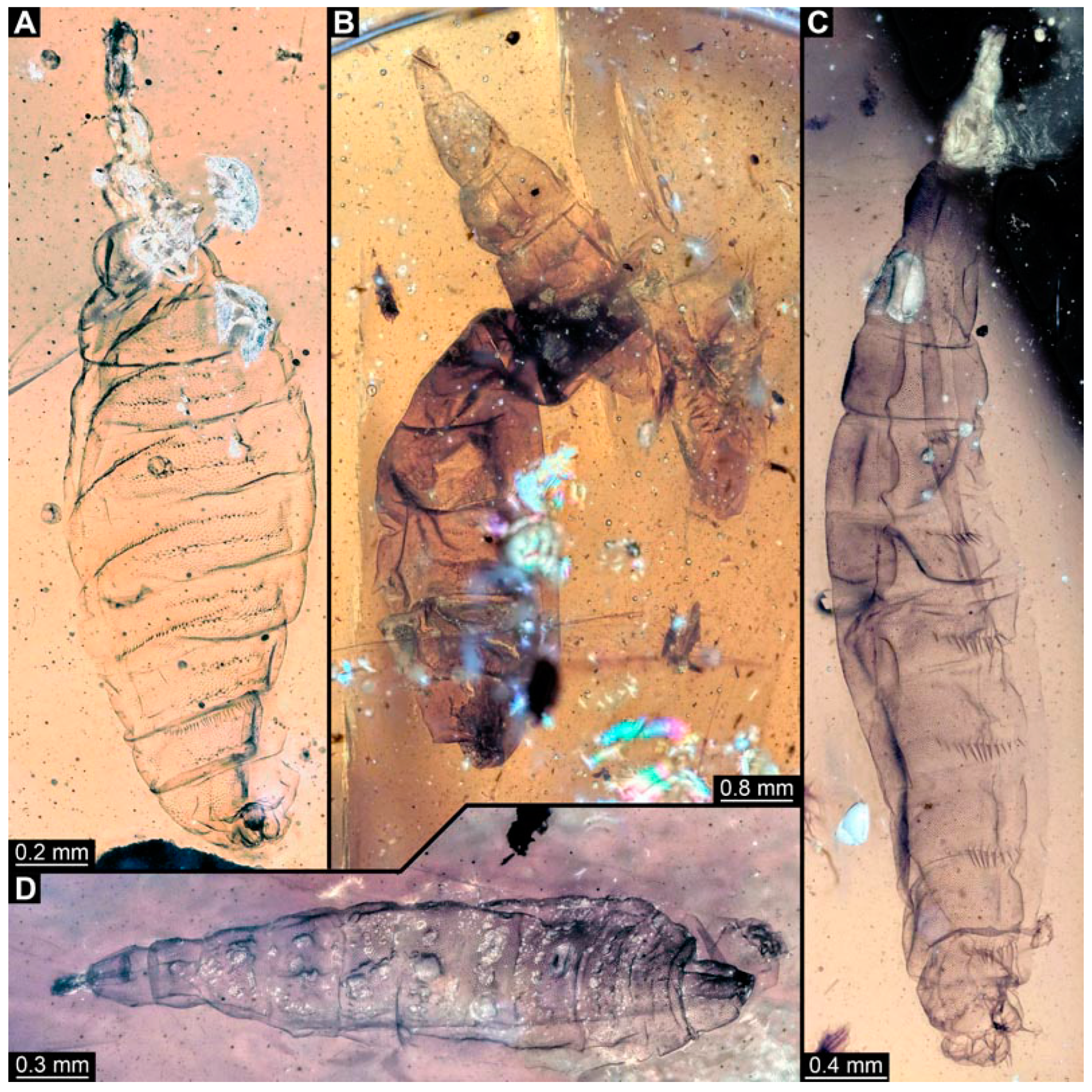
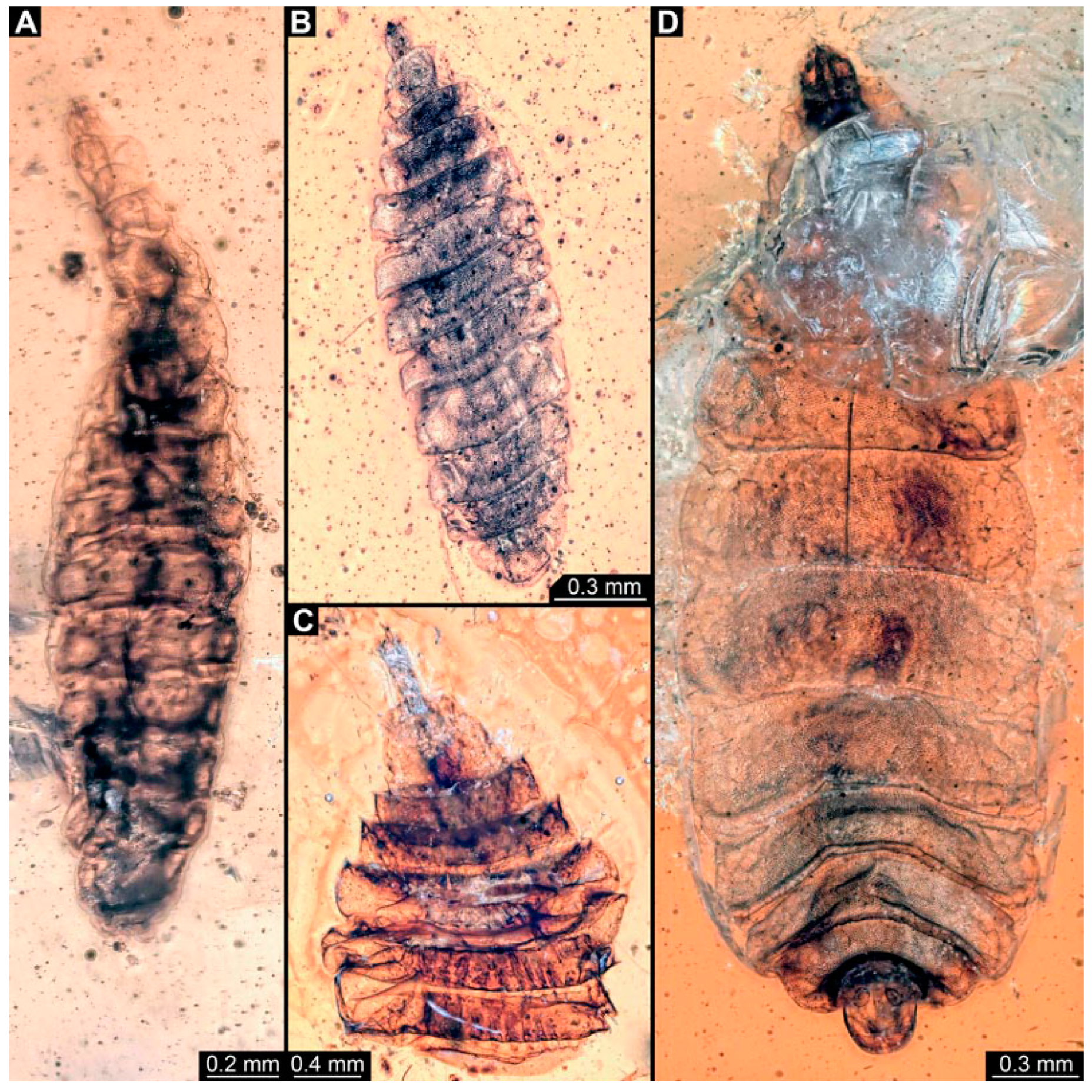
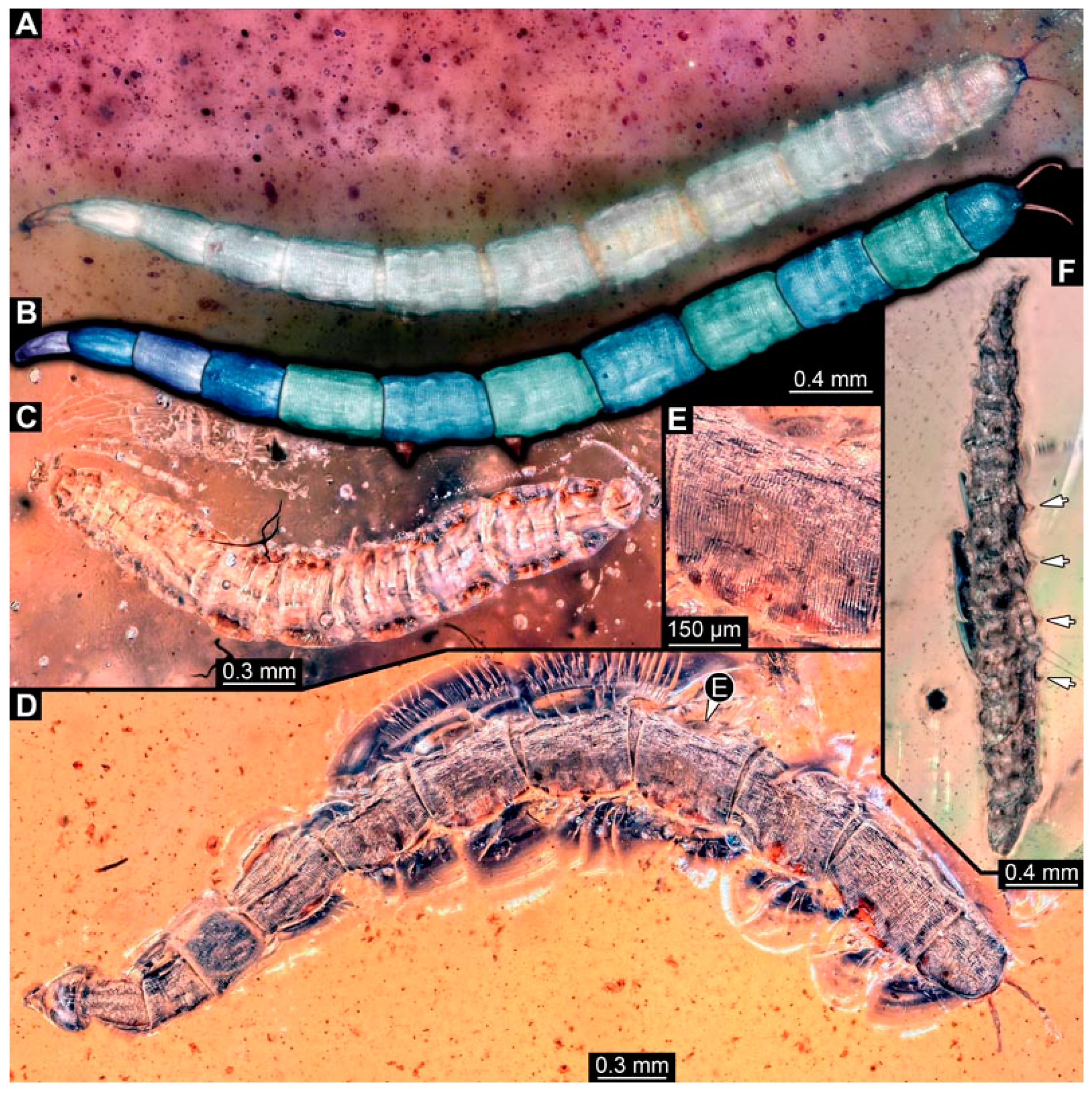
3.6. Short Descriptions of Specimen of Crochet-Bearing Morphotype
4. Discussion
4.1. Additional Specimens of Known Morphotypes: Qiyia jurassica Chen et al., 2014
4.2. Additional Specimens of Known Morphotypes: Stratiomyomorpha
4.3. Identity of New Morphotypes: Elongate Chimera Morphotype
4.4. Identity of New Morphotypes: Stouter Chimera Morphotype
4.5. Identity of New Morphotypes: Crochet Bearing Morphotype
4.6. Palaeoecology and Entrapment of Aquatic Organisms in Amber
4.7. Seeming Rarity of Brachyceran Larvae in the Fossil Record
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borkent, A.; Brown, B.V. How to Inventory Tropical Flies (Diptera)—One of the Megadiverse Orders of Insects. Zootaxa 2015, 3949, 301. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, D.A.; Engel, M.S. Evolution of the Insects; Cambridge University Press: Cambridge, UK, 2005; ISBN 978-0-521-82149-0. [Google Scholar]
- Haug, J.T. Why the Term “Larva” Is Ambiguous, or What Makes a Larva? Acta Zool. 2020, 101, 167–188. [Google Scholar] [CrossRef]
- Haug, J.T.; Labandeira, C.C.; Santiago-Blay, J.A.; Haug, C.; Brown, S. Life Habits, Hox Genes, and Affinities of a 311 Million-Year-Old Holometabolan Larva. BMC Evol. Biol. 2015, 15, 208. [Google Scholar] [CrossRef] [PubMed]
- Nel, A.; Roques, P.; Nel, P.; Prokop, J.; Steyer, J.S. The Earliest Holometabolous Insect from the Carboniferous: A “Crucial” Innovation with Delayed Success (Insecta Protomeropina Protomeropidae). Ann. Soc. Entomol. Fr. 2007, 43, 349–355. [Google Scholar] [CrossRef]
- Faria, L.; Pie, M.; Salles, F.; Soares, E. The Haeckelian Shortfall or the Tale of the Missing Semaphoronts. J. Zool. Syst. Evol. Res. 2021, 59, 359–369. [Google Scholar] [CrossRef]
- Yeo, D.; Puniamoorthy, J.; Ngiam, R.W.J.; Meier, R. Towards Holomorphology in Entomology: Rapid and Cost-effective Adult–Larva Matching Using NGS Barcodes. Syst. Entomol. 2018, 43, 678–691. [Google Scholar] [CrossRef]
- Zippel, A.; Haug, C.; Müller, P.; Haug, J.T. The First Fossil False Click Beetle Larva Preserved in Amber. PalZ 2022, 97, 209–215. [Google Scholar] [CrossRef]
- Baranov, V.; Schädel, M.; Haug, J.T. Fly Palaeo-Evo-Devo: Immature Stages of Bibionomorphan dipterans in Baltic and Bitterfeld Amber. PeerJ 2019, 7, e7843. [Google Scholar] [CrossRef] [PubMed]
- Baranov, V.; Wang, Y.; Gašparič, R.; Wedmann, S.; Haug, J.T. Eco-Morphological Diversity of Larvae of Soldier Flies and Their Closest Relatives in Deep Time. PeerJ 2020, 8, e10356. [Google Scholar] [CrossRef]
- Baranov, V.; Engel, M.; Hammel, J.; Hörnig, M.; van de Kamp, T.; Zuber, M.; Haug, J.T. Synchrotron-Radiation Computed Tomography Uncovers Ecosystem Functions of Fly Larvae in an Eocene Forest. Palaeontol. Electron. 2021, 24, 1–22. [Google Scholar] [CrossRef]
- Batelka, J.; Engel, M.S.; Prokop, J. A Remarkable Diversity of Parasitoid Beetles (Ripiphoridae) in Cretaceous Amber, with a Summary of the Mesozoic Record of Tenebrionoidea. Cretac. Res. 2018, 90, 296–310. [Google Scholar] [CrossRef]
- Lukashevich, E.D.; Krzeminski, W. New Jurassic Tanyderidae (Diptera) from Asia with First Find of Larvae. Zoosymposia 2009, 3, 155–172. [Google Scholar] [CrossRef]
- Lukashevich, E.D.; Przhiboro, A.A.; Marchal-Papier, F.; Grauvogel-Stamm, L. The Oldest Occurrence of Immature Diptera (Insecta), Middle Triassic, France. Ann. Soc. Entomol. Fr. 2010, 46, 4–22. [Google Scholar] [CrossRef]
- Lukashevich, E.D. Where the Immatures of Triassic Diptera Developed. Diversity 2023, 15, 582. [Google Scholar] [CrossRef]
- Peñalver, E.; Matamales-Andreu, R.; Nel, A.; Pérez-de La Fuente, R. Early Adaptations of True Flies (Diptera) to Moist and Aquatic Continental Environments. Pap. Palaeontol. 2022, 8, e1472. [Google Scholar] [CrossRef]
- Heiri, O.; Lotter, A.F. 9000 Years of Chironomid Assemblage Dynamics in an Alpine Lake: Long-Term Trends, Sensitivity to Disturbance, and Resilience of the Fauna. J. Paleolimnol. 2003, 30, 273–289. [Google Scholar] [CrossRef]
- Jiménez-Moreno, G.; Heiri, O.; García-Alix, A.; Anderson, R.S.; Jiménez-Espejo, F.J.; López-Blanco, C.; Jiménez, L.; Pérez-Martínez, C.; Rodrigo-Gámiz, M.; López-Avilés, A.; et al. Holocene Summer Temperature Reconstruction Based on a Chironomid Record from Sierra Nevada, Southern Spain. Quat. Sci. Rev. 2023, 319, 108343. [Google Scholar] [CrossRef]
- Amaral, A.P.; Gombos, D.; Haug, G.T.; Haug, C.; Gauweiler, J.; Hörnig, M.K.; Haug, J.T. Expanding the Fossil Record of Soldier Fly Larvae—An Important Component of the Cretaceous Amber Forest. Diversity 2023, 15, 247. [Google Scholar] [CrossRef]
- Liu, Y.; Hakim, M.; Huang, D. First Stratiomyomorphan Larvae in the Mid-Cretaceous Amber from Myanmar (Diptera: Brachycera). Cretac. Res. 2020, 106, 104265. [Google Scholar] [CrossRef]
- Poinar, G.; Jacobson, R.; Eisenberger, C.L. Early Cretaceous Phlebotomine Sand Fly Larvae (Diptera: Psychodidae). Proc. Entomol. Soc. Wash. 2006, 108, 785–792. [Google Scholar]
- Poinar, G., Jr. Early Cretaceous Trypanosomatids Associated with Fossil Sand Fly Larvae in Burmese Amber. Mem. Inst. Oswaldo Cruz 2007, 102, 635–637. [Google Scholar] [CrossRef] [PubMed]
- Baranov, V.; Haug, J.T.; Greenwalt, D.; Harbach, R. Diversity of Culicomorphan Dipterans in the Eocene Kishenehn Konservat-Lagerstätte (Montana, USA) and Its Palaeoecological Implications. Palaeontol. Electron. 2022, 25, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Badano, D.; Engel, M.S.; Basso, A.; Wang, B.; Cerretti, P. Diverse Cretaceous Larvae Reveal the Evolutionary and Behavioural History of Antlions and Lacewings. Nat. Commun. 2018, 9, 3257. [Google Scholar] [CrossRef] [PubMed]
- Baranov, V.; Hammel, J.; Gröhn, C.; Haug, J.T. Unique Fossils of Caddisfly Larvae from Baltic Amber and in Situ Amber Formation in Aquatic Ecosystems. Palaeontol. Electron. 2023, 26, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Haug, C.; Haug, G.T.; Zippel, A.; Van Der Wal, S.; Haug, J.T. The Earliest Record of Fossil Solid-Wood-Borer Larvae—Immature Beetles in 99 Million-Year-Old Myanmar Amber. Palaeoentomology 2021, 4, 390–404. [Google Scholar] [CrossRef]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.; Lei, W.; Li, Q.; Li, X. Age Constraint on Burmese Amber Based on U–Pb Dating of Zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Poinar, G. Burmese Amber: Evidence of Gondwanan Origin and Cretaceous Dispersion. Hist. Biol. 2018, 37, 155–163. [Google Scholar] [CrossRef]
- Brown, J.H.; Lomolino, M.V. Biogeography, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 1998; ISBN 978-0-87893-073-9. [Google Scholar]
- Lukashevich, E.D.; Mostovski, M.B. Hematophagous Insects in the Fossil Record. Paleontol. ZH. 2003, 37, 153–161. [Google Scholar]
- Ševčík, J.; Kaspřák, D.; Mantič, M.; Fitzgerald, S.; Ševčíková, T.; Tóthová, A.; Jaschhof, M. Molecular Phylogeny of the Megadiverse Insect Infraorder Bibionomorpha Sensu Lato (Diptera). PeerJ 2016, 4, e2563. [Google Scholar] [CrossRef]
- McAlpine, J.F. (Ed.) Manual of Nearctic Diptera; Research Branch, Agriculture Canada: Ottawa, ON, Canada, 1981; ISBN 978-0-660-10731-8. [Google Scholar]
- Lambkin, C.L.; Sinclair, B.J.; Pape, T.; Courtney, G.W.; Skevington, J.H.; Meier, R.; Yeates, D.K.; Bragoderov, V.; Wiegmann, B.M. The Phylogenetic Relationships among Infraorders and Superfamilies of Diptera Based on Morphological Evidence. Syst. Entomol. 2013, 38, 164–179. [Google Scholar] [CrossRef]
- Sinclair, B.J. A Phylogenetic Interpretation of the Brachycera (Diptera) Based on the Larval Mandible and Associated Mouthpart Structures. Syst. Entomol. 1992, 17, 233–252. [Google Scholar] [CrossRef]
- Song, N.; Xi, Y.-Q.; Yin, X.-M. Phylogenetic Relationships of Brachycera (Insecta: Diptera) Inferred from Mitochondrial Genome Sequences. Zool. J. Linn. Soc. 2022, 196, 720–739. [Google Scholar] [CrossRef]
- Woodley, N.E. Phylogeny and Classification of the “Orthorrhaphous” Brachycera. In Manual of Nearctic Diptera; McAlpine, J.F., Ed.; Research Agriculture: Ottawa, ON, Canada, 1989; Volume 3, pp. 1371–1395. [Google Scholar]
- Shin, S.; Bayless, K.M.; Winterton, S.L.; Dikow, T.; Lessard, B.D.; Yeates, D.K.; Wiegmann, B.M.; Trautwein, M.D. Taxon Sampling to Address an Ancient Rapid Radiation: A Supermatrix Phylogeny of Early Brachyceran Flies (Diptera). Syst. Entomol. 2018, 43, 277–289. [Google Scholar] [CrossRef]
- Wiegmann, B.M.; Trautwein, M.D.; Winkler, I.S.; Barr, N.B.; Kim, J.W.; Lambkin, C.; Yeates, D.K. Episodic Radiations in the Fly Tree of Life. Proc. Natl. Acad. Sci. USA 2011, 108, 5690–5695. [Google Scholar] [CrossRef] [PubMed]
- Yeates, D.K. Relationships of Extant Lower Brachycera (Diptera): A Quantitative Synthesis of Morphological Characters. Zool. Scr. 2002, 31, 105–121. [Google Scholar] [CrossRef]
- Wang, L.; Ding, S.; Cameron, S.L.; Li, X.; Liu, Y.; Yao, G.; Yang, D. Middle Jurassic Origin in India: A New Look at Evolution of Vermileonidae and Time-Scaled Relationships of Lower Brachyceran Flies. Zool. J. Linn. Soc. 2022, 194, 938–959. [Google Scholar] [CrossRef]
- Chen, J.; Wang, B.; Engel, M.S.; Wappler, T.; Jarzembowski, E.A.; Zhang, H.; Wang, X.; Zheng, X.; Rust, J. Extreme Adaptations for Aquatic Ectoparasitism in a Jurassic Fly Larva. eLife 2014, 3, e02844. [Google Scholar] [CrossRef] [PubMed]
- Greving, I.; Wilde, F.; Ogurreck, M.; Herzen, J.; Hammel, J.U.; Hipp, A.; Friedrich, F.; Lottermoser, L.; Dose, T.; Burmester, H.; et al. P05 Imaging Beamline at PETRA III: First Results. Proc. SPIE 2014, 9212, 166–173. [Google Scholar] [CrossRef]
- Wilde, F.; Ogurreck, M.; Greving, I.; Hammel, J.U.; Beckmann, F.; Hipp, A.; Lottermoser, L.; Khokhriakov, I.; Lytaev, P.; Dose, T.; et al. Micro-CT at the Imaging Beamline P05 at PETRA III. AIP Conf. Proc. 2016, 1741, 030035. [Google Scholar] [CrossRef]
- Courtney, G.; Sinclair, B.; Meier, R. Morphology and Terminology of Diptera Larvae. In Contributions to a Manual of Palaearctic Diptera (with Special Reference to Flies of Economic Importance); Papp, L., Darvas, B., Eds.; Science Herald Press: Budapest, Hungary, 2000; pp. 85–161. ISBN 978-963-04-8839-6. [Google Scholar]
- Sundermann, A.; Lohse, S.; Beck, L.A.; Haase, P. Key to the Larval Stages of Aquatic True Flies (Diptera), Based on the Operational Taxa List for Running Waters in Germany. Ann. Limnol.—Int. J. Lim. 2007, 43, 61–74. [Google Scholar] [CrossRef]
- James, M.T. Xylophagidae [Chapter] 34. In Manual of Neartic Diptera; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Agriculture Canada: Ottawa, ON, Canada, 1981; Volume 1, pp. 489–492. [Google Scholar]
- Lindner, E. Beiträge zur kenntnis der larven der Limoniidae (Diptera). Z. Morph. Ökol. Tiere 1959, 48, 209–319. [Google Scholar] [CrossRef]
- Smith, K.G.V. An Introduction to the Immature Stages of British Flies. In Handbook for the Identification of British Insects; Royal Entomological Society of London: London, UK, 1989; Volume 10, p. 280. [Google Scholar]
- Zloty, J.; Sinclair, B.J.; Pritchard, G. Discovered in Our Backyard: A New Genus and Species of a New Family from the Rocky Mountains of North America (Diptera, Tabanomorpha). Syst. Entomol. 2005, 30, 248–266. [Google Scholar] [CrossRef]
- Hennig, W. Die Sogenannten “niederen Brachycera” Im Baltischen Bernstein (Diptera: Fam. Xylophagidae, Xylomyidae, Rhagionidae, Tabanidae). Stuttg. Beitr. Naturkd. 1967, 174, 1–51. [Google Scholar]
- Zhang, J. On the Enigmatic Sinonemestrius Hong & Wang, 1990, with Description of a New Species Based on a Complete Fossil Fly (Diptera, Brachycera, Tabanomorpha, Heterostomidae). Dtsch. Entomol. Z. 2017, 64, 61–67. [Google Scholar] [CrossRef]
- Teskey, H.J. Key to Families—Larvae. In Manual of Neartic Diptera; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Agriculture Canada: Ottawa, ON, Canada, 1981; Volume 1, pp. 125–147. [Google Scholar]
- James, M.T.; Turner, W.J. Rhagionidae [Chapter] 33. In Manual of Neartic Diptera; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Agriculture Canada: Ottawa, ON, Canada, 1981; Volume 1, pp. 483–488. [Google Scholar]
- Nagatomi, A.; Stuckenberg, B.R. Insecta: Diptera, Athericidae. In Freshwater Invertebrates of the Malaysian Region; Yule, C.M., Sen, Y.-H., Eds.; Academy of Sciences Malaysia: Kuala Lumpur, Malaysia, 2004; pp. 791–797. [Google Scholar]
- Palmer, C.M.; Yeates, D.K. Phylogenetic Importance of Immature Stages: Solving the Riddle of Exeretonevra Macquart (Diptera: Xylophagidae). Ann. Entomol. Soc. Am. 2000, 93, 15–27. [Google Scholar] [CrossRef]
- Teskey, H.J.; Pechuman, L.L. Tabanidae [Chapter] 31. In Manual of Neartic Diptera; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Agriculture Canada: Ottawa, ON, Canada, 1981; Volume 1, pp. 463–478. [Google Scholar]
- Webb, D.W. Athericidae [Chapter] 32. In Manual of Neartic Diptera; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Agriculture Canada: Ottawa, ON, Canada, 1981; Volume 1, pp. 479–482. [Google Scholar]
- Wichard, W.; Gröhn, C.; Seredszus, F. Chironomidae—Non-Biting Midges—Zuckmücken. In Wasserinsekten im Baltischen Bernstein. Aquatic Insects in Baltic Amber; Verlag: Kessel, Germany, 2009; pp. 224–271. [Google Scholar]
- Xing, L.; Liu, Y.; McKellar, R.C.; Luque, J.; Li, G.; Wang, Y.; Yi, Q.; Sun, R.; Wang, E.; Audo, D. The First Shrimp Preserved in Mid-Cretaceous Kachin Amber: Systematics, Palaeoecology, and Taphonomy. Sci. Bull. 2021, 66, 1723–1726. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.R.; Dilcher, D.L. Aquatic Organisms as Amber Inclusions and Examples from a Modern Swamp Forest. Proc. Natl. Acad. Sci. USA 2007, 104, 16581–16585. [Google Scholar] [CrossRef]
- Schmidt, A.R.; Grabow, D.; Beimforde, C.; Perrichot, V.; Rikkinen, J.; Saint Martin, S.; Thiel, V.; Seyfullah, L.J. Marine Microorganisms as Amber Inclusions: Insights from Coastal Forests of New Caledonia. Foss. Rec. 2018, 21, 213–221. [Google Scholar] [CrossRef]
















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaral, A.P.; Haug, J.T.; Haug, C.; Linhart, S.; Müller, P.; Hammel, J.U.; Baranov, V. Expanding the Mesozoic Record of Early Brachyceran Fly Larvae, including New Larval Forms with Chimera-Type Morphologies. Insects 2024, 15, 270. https://doi.org/10.3390/insects15040270
Amaral AP, Haug JT, Haug C, Linhart S, Müller P, Hammel JU, Baranov V. Expanding the Mesozoic Record of Early Brachyceran Fly Larvae, including New Larval Forms with Chimera-Type Morphologies. Insects. 2024; 15(4):270. https://doi.org/10.3390/insects15040270
Chicago/Turabian StyleAmaral, André P., Joachim T. Haug, Carolin Haug, Simon Linhart, Patrick Müller, Jörg U. Hammel, and Viktor Baranov. 2024. "Expanding the Mesozoic Record of Early Brachyceran Fly Larvae, including New Larval Forms with Chimera-Type Morphologies" Insects 15, no. 4: 270. https://doi.org/10.3390/insects15040270
APA StyleAmaral, A. P., Haug, J. T., Haug, C., Linhart, S., Müller, P., Hammel, J. U., & Baranov, V. (2024). Expanding the Mesozoic Record of Early Brachyceran Fly Larvae, including New Larval Forms with Chimera-Type Morphologies. Insects, 15(4), 270. https://doi.org/10.3390/insects15040270