
Lucilia silvarum Meigen (Diptera: Calliphoridae) Is a Primary Colonizer of Domestic Cats (Felis catus)


Abstract
:Simple Summary
Abstract
1. Introduction
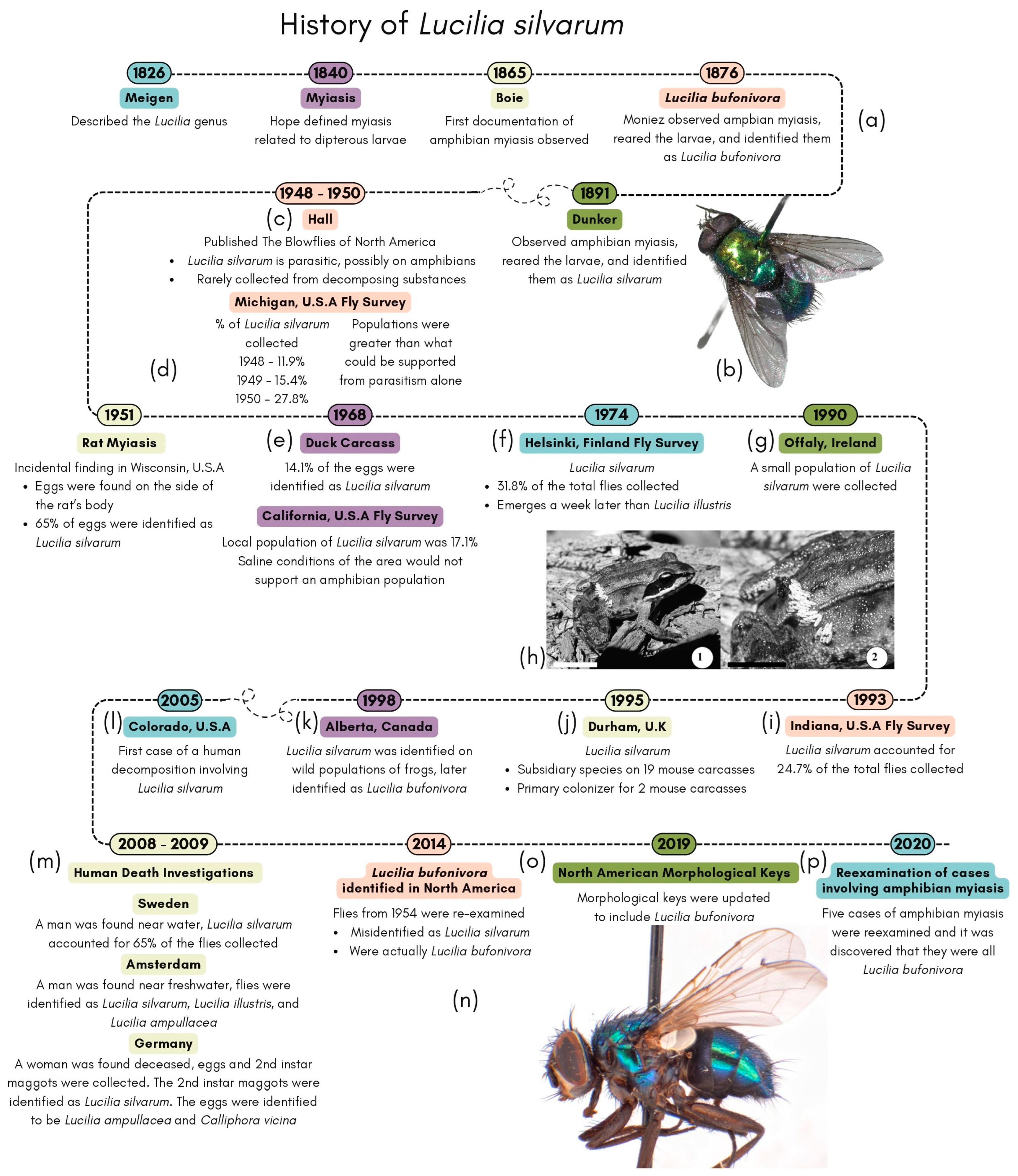
1.1. The History of Lucilia silvarum

1.2. The Misidentification of Lucilia silvarum and the Reexamination of Amphibian Myiasis Cases
1.3. Previous Studies Involving Cats
2. Materials and Methods
Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huntington, T.; Weidner, L.; Hall, R. Introduction: Current Perceptions and Status of Forensic Entomology. In Forensic Entomology: The Utility of Arthropods in Legal Investigations; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. xxiii–xxxiv. [Google Scholar] [CrossRef]
- Brundage, A.; Byrd, J.H. Forensic Entomology in Animal Cruelty Cases. Vet. Pathol. 2016, 53, 898–909. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.R.; Merritt, R.W. The Role of Aquatic Organisms in Forensic Investigations. In Forensic Entomology: The Utility of Arthropods in Legal Investigations; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar] [CrossRef]
- Nabity, P.D.; Higley, L.G.; Heng-Moss, T.M. Effects of Temperature on Development of Phormia regina (Diptera: Calliphoridae) and Use of Developmental Data in Determining Time Intervals in Forensic Entomology. J. Med. Entomol. 2006, 43, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Boatright, S.A.; Tomberlin, J.K. Effects of Temperature and Tissue Type on the Development of Cochliomyia macellaria (Diptera: Calliphoridae). J. Med. Entomol. 2010, 47, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Bambaradeniya, Y.T.B.; Karunaratne, W.A.I.P.; Tomberlin, J.K.; Goonerathne, I.; Kotakadeniya, R.B.; Magni, P.A. Effect of Temperature and Tissue Type on the Development of the Forensic Fly Chrysomya megacephala (Diptera: Calliphoridae). J. Med. Entomol. 2019, 56, 1571–1581. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Evans, L.; Wall, R. Growth Rates of the Blowfly, Lucilia sericata, on Different Body Tissues. Forensic Sci. Int. 2006, 156, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Weidner, L.M.; Nigoghosian, G.; Hanau, C.G.; Jennings, D.E. Analysis of Alternative Food Sources for Rearing Entomological Evidence. J. Med. Entomol. 2020, 57, 1407–1410. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.M.; Bauer, A.; Tomberlin, J.K. Effects of Photoperiod on the Development of Forensically Important Blow Fly Chrysomya rufifacies (Diptera: Calliphoridae). J. Med. Entomol. 2020, 57, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Nabity, P.D.; Higley, L.G.; Heng-Moss, T.M. Light-Induced Variability in Development of Forensically Important Blow Fly Phormia regina (Diptera: Calliphoridae). J. Med. Entomol. 2007, 44, 351–358. [Google Scholar] [CrossRef]
- Hanski, I. Carrion Fly Community Dynamics: Patchiness, Seasonality and Coexistence. Ecol. Entomol. 1987, 12, 257–266. [Google Scholar] [CrossRef]
- Okpara, P.; VanLaerhoven, S. Density, Temperature, and Comingled Species Affect Fitness within Carrion Communities: Coexistence in Phormia regina and Lucilia sericata (Diptera: Calliphoridae). Insects 2023, 14, 139. [Google Scholar] [CrossRef]
- MacInnis, A.E.; Higley, L.G. Competition Among Three Forensically Important Blow Fly Species (Diptera: Calliphoridae): Phormia regina, Lucilia sericata, and Chrysomya rufifacies. Environ. Entomol. 2020, 49, 1473–1479. [Google Scholar] [CrossRef] [PubMed]
- Byrd, J.H.; Butler, J.F. Effects of Temperature on Cochliomyia macellaria (Diptera: Calliphoridae) Development. J. Med. Entomol. 1996, 33, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Brundage, A. Diptera Development: A Forensic Science Perspective; IntechOpen: London, UK, 2020; Available online: www.intechopen.com (accessed on 20 July 2022).
- e Castro, C.P.; Sousa, J.P.; Arnaldos, M.I.; Gaspar, J.; García, M.D. Blowflies (Diptera: Calliphoridae) Activity in Sun Exposed and Shaded Carrion in Portugal. Ann. Société Entomol. Fr. 2011, 47, 128–139. [Google Scholar] [CrossRef]
- LeBlanc, H.; Perrault, K.; Ly, J. The Role of Decomposition Volatile Organic Compounds in Chemical Ecology. In Forensic Entomology: The Utility of Arthropods in Legal Investigations; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 485–495. [Google Scholar] [CrossRef]
- Byrd, J.; Tomberlin, J. Insects of Forensic Importance. In Forensic Entomology: The Utility of Arthropods in Legal Investigations; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 16–60. [Google Scholar] [CrossRef]
- Sanford, M.; Byrd, J.; Tomberlin, J.; Wallace, J. Entomological Evidence Collection Methods. In Forensic Entomology: The Utility of Arthropods in Legal Investigations; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 64–84. [Google Scholar] [CrossRef]
- Smith, K.V. A Manual of Forensic Entomology; Trustees of the British Museum (Natural History): London, UK, 1986. [Google Scholar] [CrossRef]
- LeBlanc, H.; Logan, J. Exploiting Insect Olfaction in Forensic Entomology. In Current Concepts in Forensic Entomology; Amendt, J., Goff, M.L., Campobasso, C.P., Grassberger, M., Eds.; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar] [CrossRef]
- Ashworth, J.R.; Wall, R. Responses of the Sheep Blowflies Lucilia sericata and Lucilia cuprina to Odour and the Development of Semiochemical Baits. Med. Vet. Entomol. 1994, 8, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Cragg, J.B. The Olfactory Behaviour of Lucilia Species (Diptera) under Natural Conditions. Ann. Appl. Biol. 1956, 44, 467–477. [Google Scholar] [CrossRef]
- Yoho, K. Necrophilous Insect Attraction to Cadaveric Volatile Organic Compounds. Master’s Thesis, Purdue University, West Lafayette, IN, USA, 2019. [Google Scholar]
- Recinos-Aguilar, Y.M.; García-García, M.D.; Malo, E.A.; Cruz-López, L.; Rojas, J.C. The Colonization of Necrophagous Larvae Accelerates the Decomposition of Chicken Carcass and the Emission of Volatile Attractants for Blowflies (Diptera: Calliphoridae). J. Med. Entomol. 2019, 56, 1590–1597. [Google Scholar] [CrossRef]
- Recinos-Aguilar, Y.M.; García-García, M.D.; Malo, E.A.; Cruz-López, L.; Cruz-Esteban, S.; Rojas, J.C. The Succession of Flies of Forensic Importance Is Influenced by Volatiles Organic Compounds Emitted During the First Hours of Decomposition of Chicken Remains. J. Med. Entomol. 2020, 57, 1411–1420. [Google Scholar] [CrossRef]
- Wall, R.; Fisher, P. Visual and Olfactory Cue Interaction in Resource-location by the Blowfly, Lucilia sericata. Physiol. Entomol. 2001, 26, 212–218. [Google Scholar] [CrossRef]
- Anderson, G. Factors That Influence Insect Succession on Carrion. In Forensic Entomology: The Utility of Arthropods in Legal Investigations; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 104–132. [Google Scholar] [CrossRef]
- Dautartas, A.; Kenyhercz, M.W.; Vidoli, G.M.; Meadows Jantz, L.; Mundorff, A.; Steadman, D.W. Differential Decomposition Among Pig, Rabbit, and Human Remains. J. Forensic Sci. 2018, 63, 1673–1683. [Google Scholar] [CrossRef]
- Matuszewski, S.; Hall, M.J.R.; Moreau, G.; Schoenly, K.G.; Tarone, A.M.; Villet, M.H. Pigs vs People: The Use of Pigs as Analogues for Humans in Forensic Entomology and Taphonomy Research. Int. J. Legal Med. 2020, 134, 793–810. [Google Scholar] [CrossRef]
- Mohr, R.M.; Tomberlin, J.K. Environmental Factors Affecting Early Carcass Attendance by Four Species of Blow Flies (Diptera: Calliphoridae) in Texas. J. Med. Entomol. 2014, 51, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Schoenly, K.G.; Haskell, N.H.; Hall, R.D.; Gbur, J.R. Comparative Performance and Complementarity of Four Sampling Methods and Arthropod Preference Tests from Human and Porcine Remains at the Forensic Anthropology Center in Knoxville, Tennessee. J. Med. Entomol. 2007, 44, 881–894. [Google Scholar] [CrossRef] [PubMed]
- Payne, J.A. A Summer Carrion Study of the Baby Pig Sus. scrofa Linnaeus. Ecology 1965, 46, 592–602. [Google Scholar] [CrossRef]
- Johnson, M.D. Seasonal and Microseral Variations in the Insect Populations on Carrion. Am. Midl. Nat. 1975, 93, 79–90. [Google Scholar] [CrossRef]
- Early, M.; Goff, M.L. Arthropod Succession Patterns in Exposed Carrion on the Island of O’Ahu, Hawaiian Islands, USA1. J. Med. Entomol. 1986, 23, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Sanford, M.R. Forensic Entomology of Decomposing Humans and Their Decomposing Pets. Forensic Sci. Int. 2015, 247, e11–e17. [Google Scholar] [CrossRef] [PubMed]
- Watson, E.J.; Carlton, C.E. Spring Succession of Necrophilous Insects on Wildlife Carcasses in Louisiana. J. Med. Entomol. 2003, 40, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Watson, E.J.; Carlton, C.E. Insect Succession and Decomposition of Wildlife Carcasses During Fall and Winter in Louisiana. J. Med. Entomol. 2005, 42, 193–203. [Google Scholar] [CrossRef]
- Nelder, M.P.; McCreadie, J.W.; Major, C.S. Blow Flies Visiting Decaying Alligators: Is Succession Synchronous or Asynchronous? Psyche 2009, 2009, 575362. [Google Scholar] [CrossRef]
- Swiger, S.L.; Hogsette, J.A.; Butler, J.F. Larval Distribution and Behavior of Chrysomya rufifacies (Macquart) (Diptera: Calliphoridae) Relative to Other Species on Florida Black Bear (Carnivora: Ursidae) Decomposing Carcasses. Neotrop. Entomol. 2014, 43, 21–26. [Google Scholar] [CrossRef]
- Lord, W.D.; Burger, J.F. Arthropods Associated with Herring Gull (Larus argentatus) and Great Black-Backed Gull (Larus marinus) Carrion on Islands in the Gulf of Maine. Environ. Entomol. 1984, 13, 1261–1268. [Google Scholar] [CrossRef]
- Bennett, G.F.; Whitworth, T.L. Studies on the Life History of Some Species of Protocalliphora (Diptera: Calliphoridae). Can. J. Zool. 1991, 69, 2048–2058. [Google Scholar] [CrossRef]
- Sawyer, S.J.; Eubanks, M.D.; Beasley, J.C.; Barton, B.T.; Puckett, R.T.; Tomeček, J.M.; Tomberlin, J.K. Vertebrate and Invertebrate Competition for Carrion in Human-impacted Environments Depends on Abiotic Factors. Ecosphere 2022, 13, e4151. [Google Scholar] [CrossRef]
- Richards, S.L.; Connelly, C.R.; Day, J.F.; Hope, T.; Ortiz, R. Arthropods Associated with Carrion in a Salt Marsh Habitat in Southeastern Florida. Fla. Entomol. 2015, 98, 613–619. [Google Scholar] [CrossRef]
- De Jong, G.D. An Annotated Checklist of the Calliphoridae (Diptera) of Colorado, with Notes on Carrion Associations and Forensic Importance. J. Kans. Entomol. Soc. 1994, 67, 378–385. Available online: https://www.jstor.org/stable/25085544 (accessed on 11 July 2023).
- Cammack, J.A.; Nelder, M.P. Cool-Weather Activity of the Forensically Important Hairy Maggot Blow Fly Chrysomya rufifacies (Macquart) (Diptera: Calliphoridae) on Carrion in Upstate South Carolina, United States. Forensic Sci. Int. 2010, 195, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Reed, H.B. A Study of Dog Carcass Communities in Tennessee, with Special Reference to the Insects. Am. Midl. Nat. 1958, 59, 213. [Google Scholar] [CrossRef]
- Bolek, M.G.; Coggins, J.R. Observations on Myiasis by the Calliphorid, Bufolucilia silvarum, in the Eastern American Toad (Bufo americanus americanus) from Southeastern Wisconsin. J. Wildl. Dis. 2002, 38, 598–603. [Google Scholar] [CrossRef]
- Bolek, M.G.; Janovy, J. Observations on Myiasis by the Calliphorids, Bufolucilia silvarum and Bufolucilia elongata, in Wood Frogs, Rana sylvatica, From Southeastern Wisconsin. J. Parasitol. 2004, 90, 1169–1171. [Google Scholar] [CrossRef]
- Eaton, B.R.; Moenting, A.E.; Paszkowski, C.A.; Shpeley, D. Myiasis by Lucilia silvarum (Calliphoridae) in Amphibian Species in Boreal Alberta, Canada. J. Parasitol. 2008, 94, 949–952. [Google Scholar] [CrossRef]
- Greenberg, B. Nocturnal Oviposition Behavior of Blow Flies (Diptera: Calliphoridae). J. Med. Entomol. 1990, 27, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Patrican, L.A.; Vaidyanathan, R. Arthropod Succession in Rats Euthanized with Carbon Dioxide and Sodium Pentobarbital. J. N. Y. Entomol. Soc. 1995, 103, 197–207. [Google Scholar]
- Tomberlin, J.K.; Adler, P.H. Seasonal Colonization and Decomposition of Rat Carrion in Water and on Land in an Open Field in South Carolina. J. Med. Entomol. 1998, 35, 704–709. Available online: https://academic.oup.com/jme/article/35/5/704/2221718 (accessed on 11 July 2023). [CrossRef] [PubMed]
- De Jong, G.D.; Hoback, W.W. Effect of Investigator Disturbance in Experimental Forensic Entomology: Succession and Community Composition. Med. Vet. Entomol. 2006, 20, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Denno, R.F.; Cothran, W.R. Competitive Interactions and Ecological Strategies of Sarcophagid and Calliphorid Flies Inhabiting Rabbit Carrion. Ann. Entomol. Soc. Am. 1976, 69, 109–113. [Google Scholar] [CrossRef]
- De Jong, G.D.; Chadwick, J.W. Decomposition and Arthropod Succession on Exposed Rabbit Carrion During Summer at High Altitudes in Colorado, USA. J. Med. Entomol. 1999, 36, 833–845. Available online: https://academic.oup.com/jme/article/36/6/833/904279 (accessed on 11 July 2023). [CrossRef]
- Joy, J.E.; Herrell, M.L.; Rogers, P.C. Larval Fly Activity on Sunlit Versus Shaded Raccoon Carrion in Southwestern West Virginia with Special Reference to the Black Blowfly (Diptera: Calliphoridae). J. Med. Entomol. 2002, 39, 392–397. Available online: https://academic.oup.com/jme/article/39/2/392/884916 (accessed on 11 July 2023). [CrossRef]
- Slone, D.H.; Gruner, S.V. Thermoregulation in Larval Aggregations of Carrion-Feeding Blow Flies (Diptera: Calliphoridae). J. Med. Entomol. 2007, 44, 516–523. [Google Scholar] [CrossRef]
- Bugajski, K.N.; Seddon, C.C.; Williams, R.E. A Comparison of Blow Fly (Diptera: Calliphoridae) and Beetle (Coleoptera) Activity on Refrigerated Only versus Frozen-Thawed Pig Carcasses in Indiana. J. Med. Entomol. 2011, 48, 1231–1235. [Google Scholar] [CrossRef]
- Bugajski, K.; Tolle, C. Effect of Water on Blow Fly (Diptera: Calliphoridae) Colonization of Pigs in Northwest Indiana. Proc. Indiana Acad. Sci. 2014, 123, 67–71. [Google Scholar]
- Zurawski, K.N.; Benbow, M.E.; Miller, J.R.; Merritt, R.W. Examination of Nocturnal Blow Fly (Diptera: Calliphoridae) Oviposition on Pig Carcasses in Mid-Michigan. J. Med. Entomol. 2009, 46, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Weidner, L.M.; Monzon, M.A.; Hamilton, G.C. Death Eaters Respond to the Dark Mark of Decomposition Day and Night: Observations of Initial Insect Activity on Piglet Carcasses. Int. J. Legal Med. 2016, 130, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Mañas-Jordá, S.; León-Cortés, J.L.; García-García, M.D.; Caballero, U.; Infante, F. Dipteran Diversity and Ecological Succession on Dead Pigs in Contrasting Mountain Habitats of Chiapas, Mexico. J. Med. Entomol. 2018, 55, 59–68. [Google Scholar] [CrossRef]
- Ambercrombie, J. Myiasis in the Eastern Box Turtle Caused by Phaenicia coeruleiviridis (Diptera: Calliphoridae). J. Wash. Acad. Sci. 1977, 67, 155–156. [Google Scholar]
- Abell, D.H.; Wasti, S.S.; Hartmann, G.C. Saprophagous Arthropod Fauna Associated with Turtle Carrion. Appl. Entomol. Zool. 1982, 17, 301–307. [Google Scholar] [CrossRef]
- Byrd, J.H.; Brundage, A. Forensic Entomology. In Veterinary Forensic Medicine and Forensic Sciences; Byrd, J.H., Norris, P., Bradley-Siemens, N., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 67–112. [Google Scholar] [CrossRef]
- Arias-Robledo, G.; Stark, T.; Wall, R.L.; Stevens, J.R. The Toad Fly Lucilia bufonivora: Its Evolutionary Status and Molecular Identification. Med. Vet. Entomol. 2019, 33, 131–139. [Google Scholar] [CrossRef]
- Zumpt, F. Myiasis in Man and Animals in the Old World; Butterworth & Co.: London, UK, 1965. [Google Scholar]
- Dodge, H. A Possible Case of Blowfly Myiasis in a Rat, with Notes on the Bionomics of Bufolucilia silvarum (Mg.); Entomological News: Philadelphia, PA, USA, 1952; Volume 62–63. [Google Scholar]
- Pezzi, M.; Bonacci, T.; Leis, M.; Mamolini, E.; Marchetti, M.G.; Krčmar, S.; Chicca, M.; Del Zingaro, C.N.F.; Faucheux, M.J.; Scapoli, C. Myiasis in Domestic Cats: A Global Review. Parasit. Vectors 2019, 12, 372. [Google Scholar] [CrossRef] [PubMed]
- Weddeling, K.; Kordges, T. Lucilia bufonivora Infestation (Myiasis) of Amphibians in Northrhine Westphalia-Distribution, Host Species, Ecology and Phenology. Z. Feldherpetol 2008, 15, 183–202. [Google Scholar]
- Whitworth, T.L.; Bolek, M.G.; Arias-Robledo, G. Lucilia bufonivora, Not Lucilia silvarum (Diptera: Calliphoridae), Causes Myiasis in Anurans in North America with Notes about Lucilia elongata and Lucilia thatuna. J. Med. Entomol. 2021, 58, 88–92. [Google Scholar] [CrossRef]
- Wolff, B.; Tracy, L. Pseudacris crucifer (Spring Peeper) Myiasis. Herpetol. Rev. 2013, 44, 498–499. [Google Scholar]
- Bleakney, J.S. First North American Record of Bufolucilia silvarum (Meigen) (Diptera: Calliphoridae) Parasitizing Bufo Terrestris Americanus Holbrook. Can. Entomol. 1963, 95, 107. [Google Scholar] [CrossRef]
- Anderson, R.C.; Bennett, G.F. Ophthalmic Myiasis in Amphibians in Algonquin Park, Ontario, Canada. Can. J. Zool. 1963, 41, 1169–1170. [Google Scholar] [CrossRef]
- Briggs, J.L. A Case of Bufolucilia elongata Shannon 1924 (Diptera: Calliphoridae) Myiasis in the American Toad, Bufo americanus Holbrook 1836. J. Parasitol. 1975, 61, 412. [Google Scholar] [CrossRef]
- Thompson, M.; Bolek, M.; Rea, R. Calliphorid Parasitism Causing Myiasis in Amphibians: A New Record for Anaxyrus boreas (Western Toad) in British Columbia, Canada. Herpetol. Rev. 2021, 52, 303–307. [Google Scholar]
- Jones, N.; Whitworth, T.; Marshall, S.A. Blow Flies of North America Keys to the Subfamilies and Genera of Calliphoridae, and to the Species of the Subfamilies Calliphorinae, Luciliinae and Chrysomyinae. Can. J. Arthropod Identif. 2019, 39. [Google Scholar] [CrossRef]
- Marshall, S.; Whitworth, T.; Roscoe, L. Blow Flies (Diptera: Calliphoridae) of Eastern Canada with a Key to Calliphoridae Subfamilies and Genera of Eastern North America, and a Key to the Eastern Canadian Species of Calliphorinae, Luciliinae, and Chrysomyiinae. Can. J. Arth. Identif. 2011, 11, 1–93. [Google Scholar]
- Hall, D. The Blowflies of North America; Thomas Say Foundation: Baltimore, MD, USA, 1948; Volume 4. [Google Scholar]
- Whitworth, T. Keys to the Genera and Species of Blow Flies (Diptera: Calliphoridae) of America North of Mexico. Proc. Entomol. Soc. 2006, 108, 698–725. [Google Scholar]
- Haskell, N. Factors Affecting Diurnal Flight and Oviposition Periods of Blow Flies (Diptera: Calliphoridae) in Indiana. Ph.D. Dissertation, Purdue University, West Lafayette, IN, USA, 1993. [Google Scholar]
- Davies, L. Seasonal and Spatial Changes in Blowfly Production from Small and Large Carcasses at Durham in Lowland Northeast England. Med. Vet. Entomol. 1999, 13, 245–251. [Google Scholar] [CrossRef]
- Brothers, D. Notes on the Saprophagous Activity of Bufolucilia silvarum (Meigen) (Diptera: Calliphoridae); The Pan-Pacific Entomologist: San Francisco, CA, USA, 1970; Volume 46. [Google Scholar]
- Blackith, R.E.; Blackith, R.M. Insect Infestations of Small Corpses. J. Nat. Hist. 1990, 24, 699–709. [Google Scholar] [CrossRef]
- Adair, T.W.; Kondratieff, B.C. Three Species of Insects Collected from an Adult Human Corpse Above 3300 m in Elevation: A Review of a Case from Colorado. J. Forensic Sci. 2006, 51, 1164–1165. [Google Scholar] [CrossRef]
- Fremdt, H.; Szpila, K.; Huijbregts, J.; Lindström, A.; Zehner, R.; Amendt, J. Lucilia silvarum Meigen, 1826 (Diptera: Calliphoridae)—A New Species of Interest for Forensic Entomology in Europe. Forensic Sci. Int. 2012, 222, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Tantawi, T.I.; Whitworth, T. First Record of Lucilia bufonivora Moniez, 1876 (Diptera: Calliphoridae) from North America and Key to North American Species of the L. bufonivora Species Group. Zootaxa 2014, 3881, 101. [Google Scholar] [CrossRef] [PubMed]
- Arias-Robledo, G.; Wall, R.; Szpila, K.; Shpeley, D.; Whitworth, T.; Stark, T.; King, R.A.; Stevens, J.R. Ecological and Geographical Speciation in Lucilia bufonivora: The Evolution of Amphibian Obligate Parasitism. Int. J. Parasitol. Parasites Wildl. 2019, 10, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Bagsby, K.E.; Byrd, J.; Stern, A.; Hans, K.R. Initial Blow Fly (Diptera: Calliphoridae) Colonization of Cats (Felis catus) in Indiana. J. Med. Entomol. 2024, in press. [Google Scholar] [CrossRef]
- Perez, A.E.; Haskell, N.H.; Wells, J.D. Commonly Used Intercarcass Distances Appear to Be Sufficient to Ensure Independence of Carrion Insect Succession Pattern. Ann. Entomol. Soc. Am. 2015, 109, 72–80. [Google Scholar] [CrossRef]
- Woods, A.; Omernik, J.; Brockman, C.; Gerber, T.; Hosteter, W.; Azevedo, S. Level III and IV Ecoregions by State. US Geological Survey. Available online: https://www.epa.gov/eco-research/ecoregion-download-files-state-region-5 (accessed on 13 December 2023).
- King, K.; Toddey, D.; Eastern Corn Belt (ECB). USDA. Available online: https://ltar.ars.usda.gov/wp-content/uploads/2021/08/ECB_Climate_Final_Edits.pdf (accessed on 13 December 2023).
- Hofer, I.M.J.; Hart, A.J.; Martín-Vega, D.; Hall, M.J.R. Estimating Crime Scene Temperatures from Nearby Meteorological Station Data. Forensic Sci. Int. 2020, 306, 110028. [Google Scholar] [CrossRef] [PubMed]
- Archer, M.S. The Effect of Time After Body Discovery on the Accuracy of Retrospective Weather Station Ambient Temperature Corrections in Forensic Entomology. J. Forensic Sci. 2004, 49, JFS2003258. [Google Scholar] [CrossRef]
- Matuszewski, S.; Mądra-Bielewicz, A. Validation of Temperature Methods for the Estimation of Pre-Appearance Interval in Carrion Insects. Forensic Sci. Med. Pathol. 2016, 12, 50–57. [Google Scholar] [CrossRef]
- Hans, K.R.; Yoho, K.; Robbins, H.; Weidner, L.M. A Review of the Forensic Entomology Literature in the Midwestern United States. WIREs Forensic Sci. 2021, 3, e1424. [Google Scholar] [CrossRef]
- Weidner, L.M.; Nigoghosian, G.; Yoho, K.C.; Parrott, J.J.; Hans, K.R. An Analysis of Forensically Important Insects Associated with Human Remains in Indiana, U.S.A. J. Forensic Sci. 2020, 65, 2036–2041. [Google Scholar] [CrossRef]
- Weidner, L.M.; Tomberlin, J.K.; Hamilton, G.C. Development of Lucilia coeruleiviridis (Diptera: Calliphoridae) in New Jersey, USA. Fla. Entomol. 2014, 97, 849–851. [Google Scholar] [CrossRef]

{kind=link}
| Animal Model | Author, Year | Location | Season |
|---|---|---|---|
| Alligator | Watson and Carlton, 2003 [37] | Louisiana | Spring |
| Watson and Carlton, 2005 [38] | Louisiana | Winter, Fall | |
| Nelder et al., 2009 [39] | Alabama | Spring | |
| Bear | Watson and Carlton, 2003 [37] | Louisiana | Spring |
| Watson and Carlton, 2005 [38] | Louisiana | Winter, Fall | |
| Swiger et al., 2014 [40] | Florida | Summer | |
| Bird | Lord and Burger, 1984 [41] | New Hampshire | Spring, Summer, Fall |
| Bennett and Whitworth, 1991 [42] | Ontario, Canada | Summer | |
| Sawyer et al., 2022 [43] | Texas | Winter, Summer | |
| Cat | Johnson, 1975 [34] | Illinois | Spring, Summer, Fall |
| Early and Goff, 1986 [35] | Hawaii | Winter, Summer | |
| Sanford, 2015 [36] | Texas | Spring | |
| Bobcat | Richards et al., 2015 [44] | Florida | Fall |
| Chipmunk | Payne, 1965 [33] | South Carolina | Spring, Summer |
| Deer | De Jong, 1994 [45] | Colorado | |
| Watson and Carlton, 2003 [37] | Louisiana | Spring | |
| Watson and Carlton, 2005 [38] | Louisiana | Winter, Fall | |
| Cammack and Nelder, 2010 [46] | South Carolina | Fall | |
| Dog | Reed, 1958 [47] | Tennessee | Spring |
| De Jong, 1994 [45] | Colorado | ||
| Sanford, 2015 [36] | Texas | Spring | |
| Coyote | Richards et al., 2015 [44] | Florida | Winter |
| Frog/Toad | Payne, 1965 [33] | South Carolina | Spring, Summer |
| Bolek and Coggins, 2002 [48] | Wisconsin | Spring, Summer | |
| Bolek and Janovy, 2004 [49] | Wisconsin | Summer | |
| Eaton et al., 2008 [50] | Alberta, Canada | Spring, Sumer | |
| Mouse/Rat | Payne, 1965 [33] | South Carolina | Spring, Summer |
| Greenberg, 1990 [51] | Illinois | Summer | |
| De Jong, 1994 [45] | Colorado | ||
| Patrician and Vaidyanathan, 1995 [52] | New York | Fall | |
| Tomberlin and Adler, 1998 [53] | South Carolina | Winter, Summer | |
| De Jong and Hoback, 2006 [54] | Colorado | Summer | |
| Sawyer et al., 2022 [43] | Texas | Winter, Summer | |
| Opossum | Johnson, 1975 [34] | Illinois | Spring, Summer, Fall |
| Richards et al., 2015 [44] | Florida | Summer | |
| Rabbit | Dautaras et al., 2018 [29] | Tennessee | Winter, Spring, Summer |
| Denno and Cothram, 1975 [55] | California | Winter, Spring, Summer, Fall | |
| Johnson, 1975 [34] | Illinois | Spring, Summer, Fall | |
| De Jong and Chadwich, 1999 [56] | Colorado | Summer | |
| Racoon | De Jong, 1994 [45] | Colorado | |
| Joy et al., 2002 [57] | West Virginia | Spring | |
| Shrew | Payne, 1965 [33] | South Carolina | Spring, Summer |
| Skunk | De Jong, 1994 [45] | Colorado | |
| Swine | Payne, 1965 [33] | South Carolina | Spring, Summer |
| Watson and Carlton, 2003 [37] | Louisiana | Spring | |
| Watson and Carlton, 2005 [38] | Louisiana | Winter, Fall | |
| Slone and Gruner, 2007 [58] | Indiana | Summer, Fall | |
| Slone and Gruner, 2007 [58] | Florida | Winter, Spring | |
| Bugajski et al., 2011 [59] | Indiana | Summer | |
| Bugajski and Tolle, 2014 [60] | Indiana | Fall | |
| Mohr and Tomberlin, 2014 [31] | Texas | Winter, Summer | |
| Zurawski et al., 2014 [61] | Michigan | Summer | |
| Weidner et al., 2016 [62] | New Jersey | Summer | |
| Dautaras et al., 2018 [29] | Tennessee | Winter, Spring, Summer | |
| Mañas-Jordá et al., 2018 [63] | Mexico | Summer, Fall | |
| Matuszewski et al., 2019 [30] | |||
| Turtle | Ambercrombie, 1977 [64] | Maryland | Fall |
| Abell et al., 1982 [65] | Massachusetts | Summer | |
| De Jong, 1994 [45] | Colorado |
| Lucilia silvarum 1 | Lucilia bufonivora 1 |
|---|---|
| Subcostal sclerite without setae | |
| Black basicosta | |
| Palp black or brown | |
| Two postsutural intra-alar setae | |
| Presutural intra-alar seta present | |
| Male upper calypter pale, lower calypter tan | Male upper and lower calypter pale |
| Three postsutural acrostichal bristles | Two postsutural acrostichal bristles |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bagsby, K.; Hans, K. Lucilia silvarum Meigen (Diptera: Calliphoridae) Is a Primary Colonizer of Domestic Cats (Felis catus). Insects 2024, 15, 32. https://doi.org/10.3390/insects15010032
Bagsby K, Hans K. Lucilia silvarum Meigen (Diptera: Calliphoridae) Is a Primary Colonizer of Domestic Cats (Felis catus). Insects. 2024; 15(1):32. https://doi.org/10.3390/insects15010032
Chicago/Turabian StyleBagsby, Kelly, and Krystal Hans. 2024. "Lucilia silvarum Meigen (Diptera: Calliphoridae) Is a Primary Colonizer of Domestic Cats (Felis catus)" Insects 15, no. 1: 32. https://doi.org/10.3390/insects15010032
APA StyleBagsby, K., & Hans, K. (2024). Lucilia silvarum Meigen (Diptera: Calliphoridae) Is a Primary Colonizer of Domestic Cats (Felis catus). Insects, 15(1), 32. https://doi.org/10.3390/insects15010032