Virology, Epidemiology and Pathology of Glossina Hytrosavirus, and Its Control Prospects in Laboratory Colonies of the Tsetse Fly, Glossina pallidipes (Diptera; Glossinidae)



Abstract
:
1. Introduction

2. An Overview of Tsetse Fly Control Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Investigator(s) | Year | Major contribution(s) | Ref. |
|---|---|---|---|
| Whitnall | 1932, 34 | First published record of SGH Glossina spp. | [55,56] |
| Burtt | 1945 | Suggested that SGH is sex-linked | [57] |
| Jenni et al. | 1973, 74, 76 | Described virus particles in G. morsitans and G. fuscipes fuscipes; suggested Golgi-ER viral assembly | [58,59,60,61] |
| Lyon | 1973 | First published record of SGH in M. equestris | [62] |
| Jaenson | 1978 | First clear association of viral particles with SGH | [63] |
| Amargier et al. | 1979 | Reported SGH in M. equestris | [64] |
| Otieno et al. | 1980 | Reported SGH as common feature in wild G. pallidipes | [65] |
| Opiyo | 1983 | Reported poor productivity of G. pallidipes colony at Kenya Trypanosomosis Research Institute (KETRI), Kenya | [66] |
| Odindo et al. | 1981, 83, 86 | Demonstrated that viral particles are infectious per os; First report that Glossina virus has dsDNA genome | [67,68,69,70] |
| Jaenson | 1986 | First report on reduced insemination rates, fecundity and lifespan in laboratory colonies of G. pallidipes | [71] |
| Ellis et al. | 1987 | Reported SGH in Zimbabwe and Ivory Coast | [72,73] |
| International Atomic Energy Agency | 1987, 89 | Reported poor productivity of G. pallidipes colonies at IPCL, Seibersdorf, Austria | |
| Odindo | 1988 | Proposed Glossina virus as a bio-control agent | [74] |
| Jura et al. | 1988, 89, 92, 93 | Demonstrated transmission of Glossina virus after artificial infection | [75,76,77,78] |
| Kokwaro et al. | 1990–1991 | Cytopathology of virus particles in tsetse salivary glands | [79,80] |
| Shaw | 1993 | Reported SGH in G. m. swyenatoni and G. brevipalpis | [81] |
| Coler et al. | 1993 | First published record of SGH in M. domestica | [82] |
| Sang | 1996–1999 | Reported SGHV in tsetse milk glands, mid-gut and male accessory reproductive glands | [83,84,85,86] |
| International Atomic Energy Agency | 2002 | Collapse of an Ethiopian-derived G. pallidipes colony at IPCL, Seibersdorf, Austria | |
| Kokwaro | 2006 | Reported viral particles in male accessory reproductive glands of G. m. morsitans Westwood | [87] |
| Abd-Alla et al.; Garcia-Maruniak et al. | 2008 | G. pallidipes and M. domestica SGHVs genome sequenced | [26,27] |
| Abd-Alla et al. | 2009 | Establishment Hytrosaviridae family | [27] |
| Salem et al. | 2009 | Transcription analysis of M. domestica SGHV | [88] |
| Kariithi et al. | 2010–2013 | Described proteome and morphogenesis of Glossina SGHV | [89,90] |
| Prompiboon et al. | 2010 | Reported wild-wide distribution of SGHV in M. domestica | [91] |
| Luo and Zheng | 2010 | SGHV-like virus described in accessory gland filaments of the parasitic braconid wasp, D. longicuadata | [92] |
| Boucias et al. | 2013 | Described the role of endosymbionts on trans generational trans mission of SGHV in G. pallidipes | [93] |
| Abd-Alla et al. | 2013 | Reported successful management of Glossina hytrosavirus and eradication of SGH in G. pallidipes colonies at IPCL, Seibersdorf | [94] |
3. A Historic Chronology of the Discovery and Biogeography of SGH
3.1. SGH in Dipteran Insects
3.2. Possible Hytrosaviruses in other Insect Species
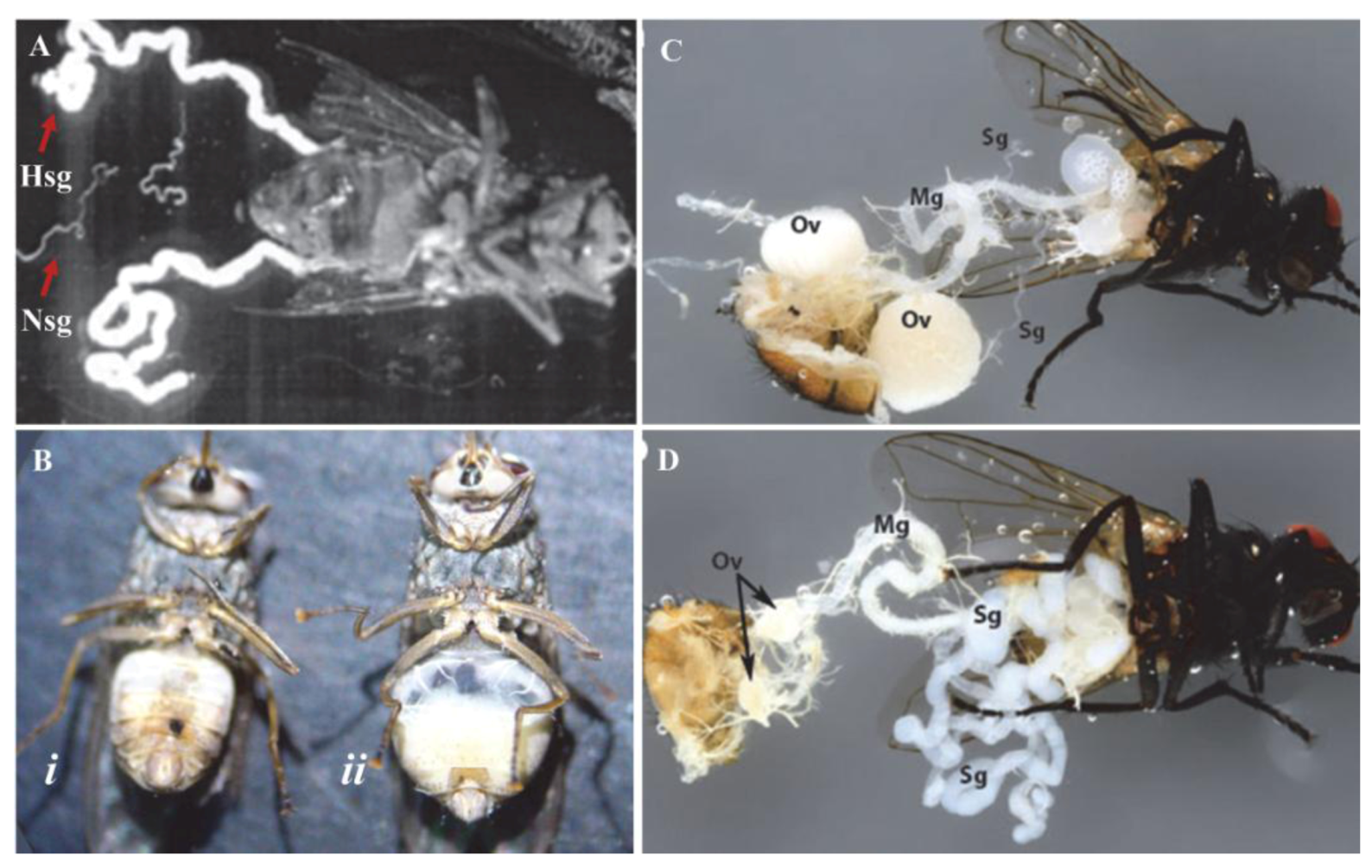
4. Pathology of Hytrosaviruses

5. Virology of the Hytrosaviruses
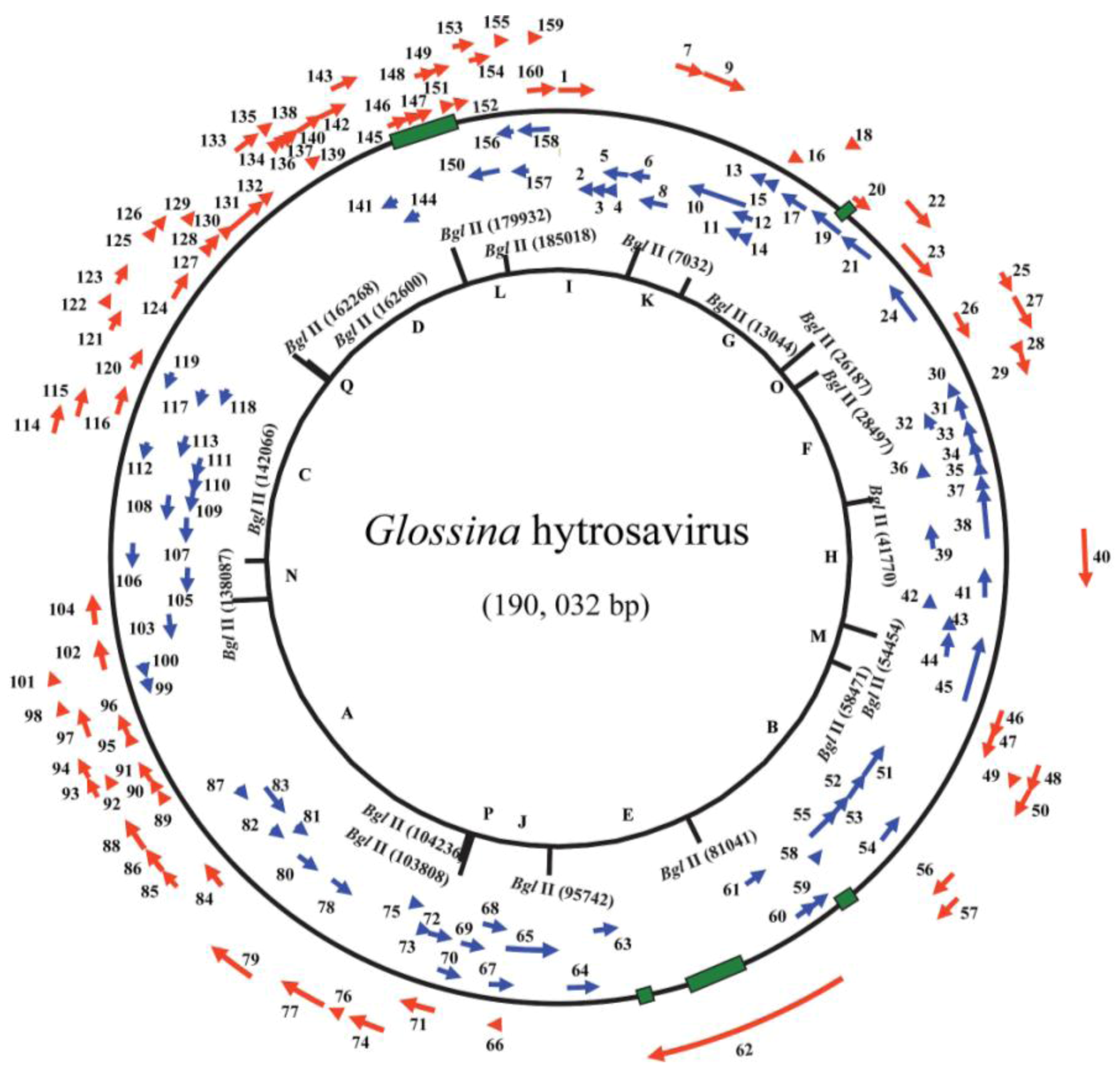
5.1. Genomics of Hytrosaviruses

5.2. Classification and Phylogeny of Hytrosaviruses
| Key characteristics | GpSGHV | MdSGHV | Ref. | |
|---|---|---|---|---|
| Biological | Replication site(s) | Salivary glands, milk glands | Salivary glands | [75,82,83,104,130] |
| Infection phenotype | Symptomatic; asymptomatic | symptomatic | [28,93] | |
| Symptoms besides SGH | Male/female gonadal abnormities | Under-developed ovaries | [75,82,93,105] | |
| Vertical (trans-generational) transmission | Milk glands, trans-ovarian | No evid ence available to-date | [52,75,82,93,105] | |
| Horizontal transmission | Oral (salivary) secretions | Oral (salivary) secretions and excreta | [25,52,131] | |
| Sterilizing agent | Male and female infertility | Female infertility | [86,105] | |
| Impact on host behavior | Impaired feeding | Mating disruption | [84,105,132] | |
| Morphogenesis | Cytoplasmic envelopment, egress by disintegration or rapture of the plasma membrane | Cytoplasmic, egress via budding on the plasma membrane | [93,133,134] | |
| Structural | Virion size | 50 × 1000 nm | 65 × 550 nm | [120,134] |
| Ultra-structure | Nucleocapsid, tegument, envelop, outer surface projections | Nucleocapsid, envelop | [89] | |
| Virion topography | Helical surface projections | Braided, bead-like surface | [89,120] | |
| Molecular | Genome size | 190,032 bp | 124,279 bp | [26,120] |
| G + C content (%) | 28 | 44 | [26,120] | |
| No. of RFs | 160 | 108 | [26,120] | |
| Shared ORFs between GpSGHV and MdSGHV | 41 | 37 | [90,128] | |
| ORFs homologs in other large dsDNA viruses | Nudivirus (11), whispovirus (4), baculovirus (12) | Nudivirus (17), whispovirus (6), baculovirus (12) | [26,120] | |
5.3. Proteomics and Interactomics of Hytrosaviruses
5.4. Latency of Hytrosaviruses
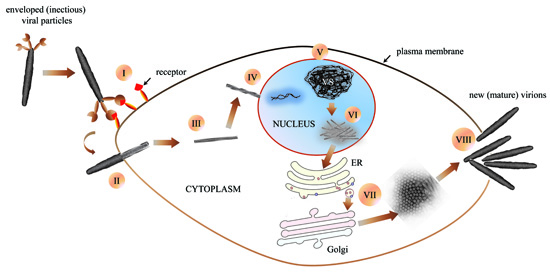
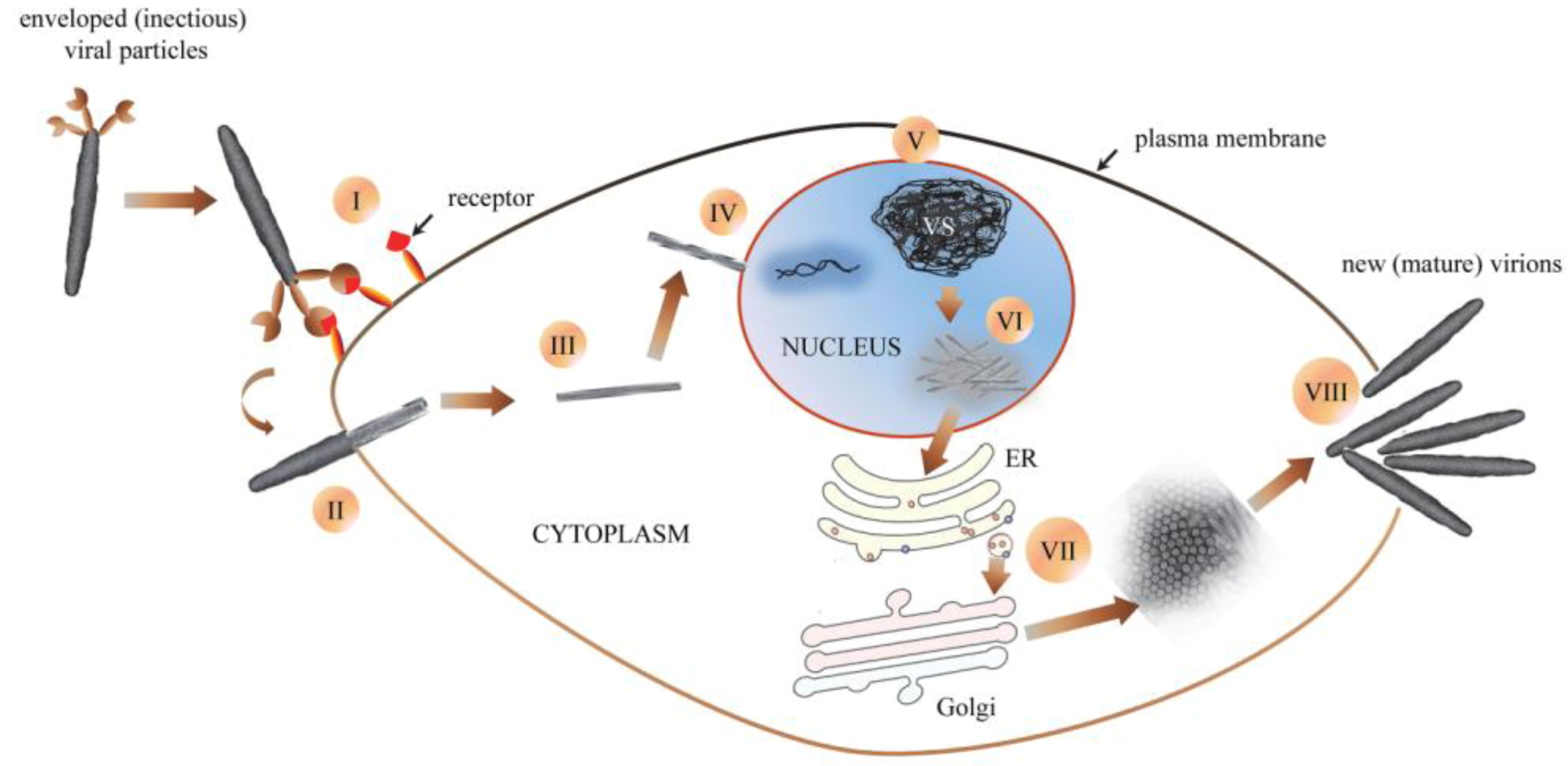
5.5. Replication, Morphology and Morphogenesis of Hytrosaviruses

6. Epidemiology of Hytrosaviruses
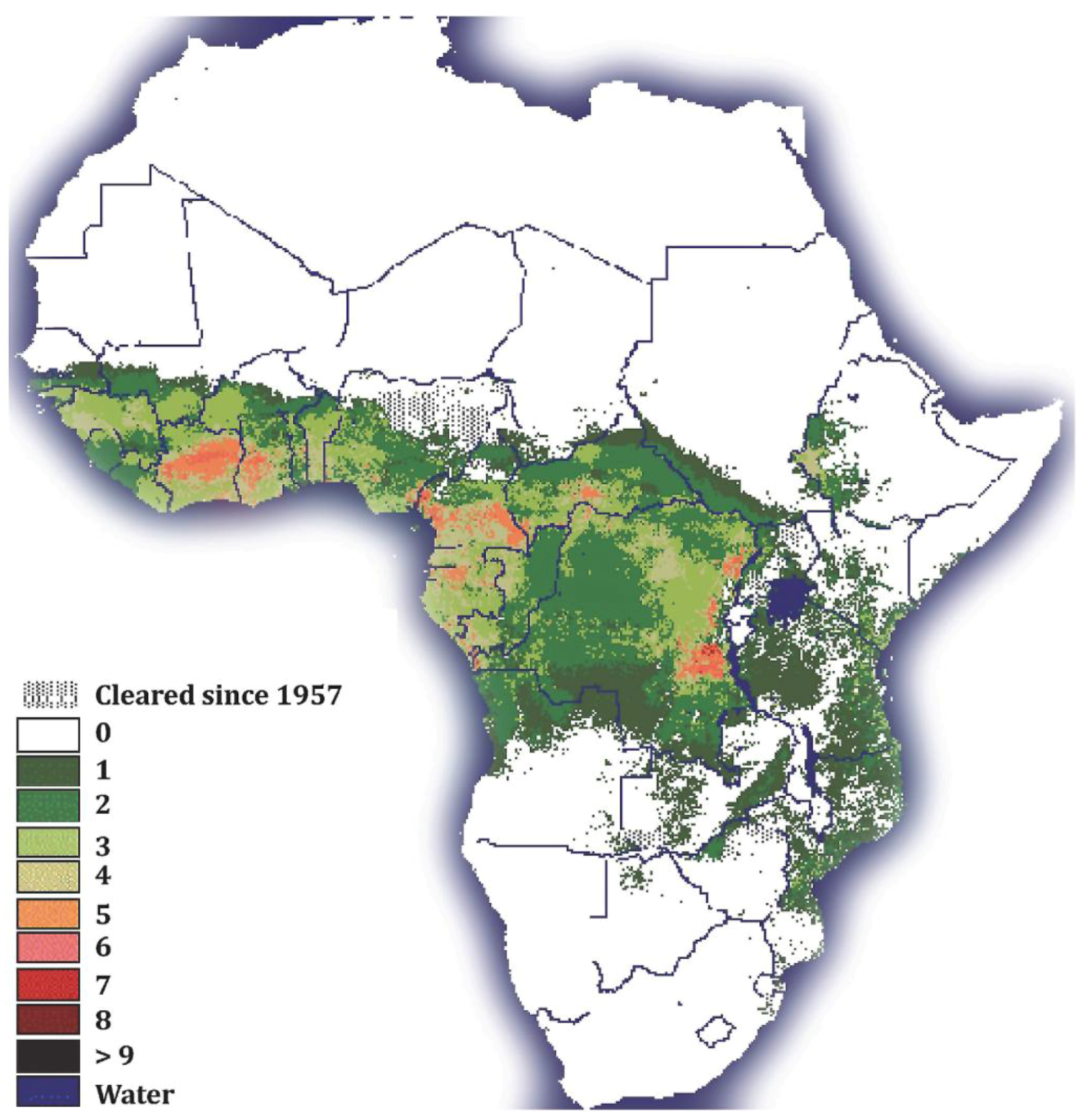
6.1. Prevalence and Ecogeography of Hytrosaviruses
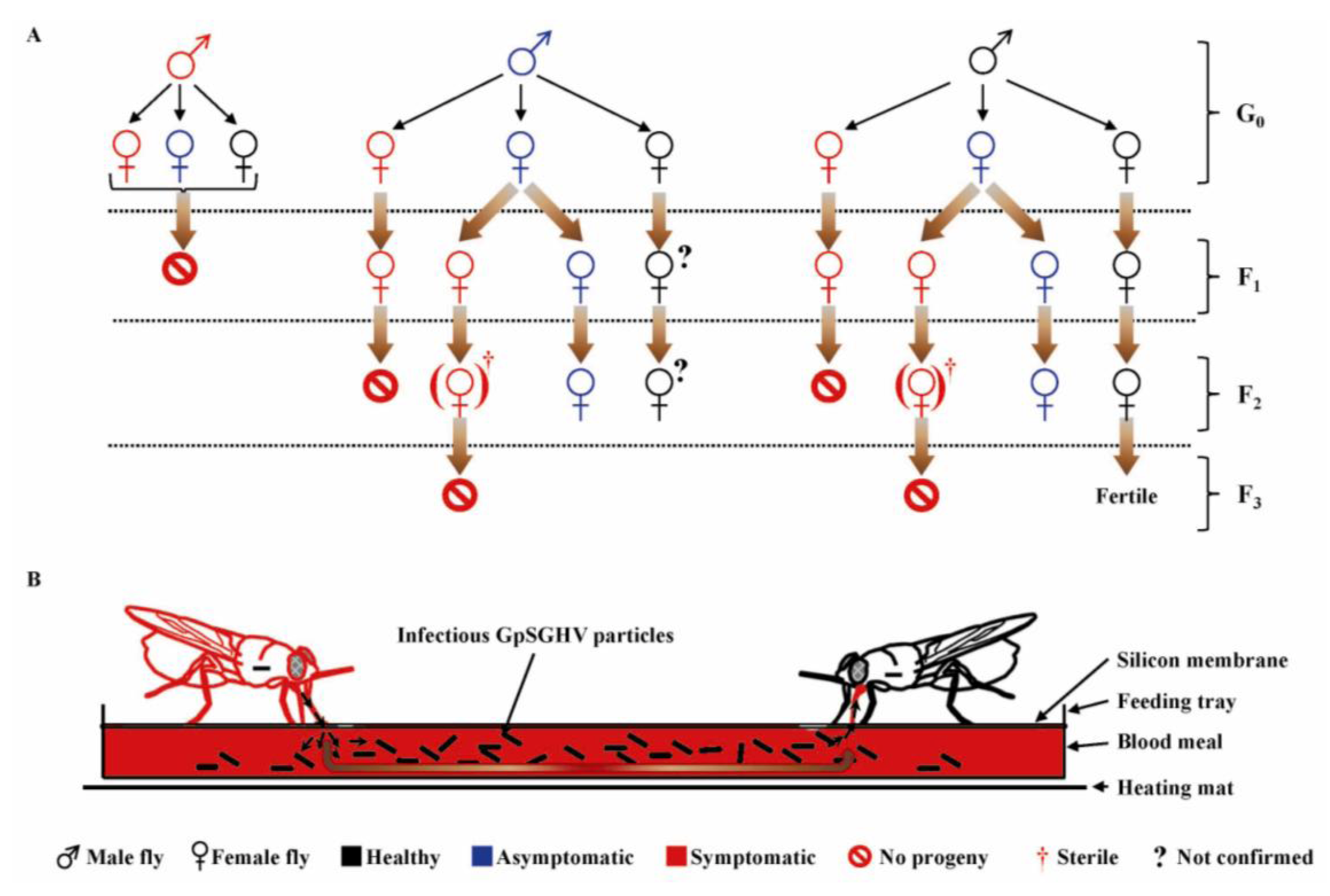
6.2. Transmission Dynamics of GpSGHV in the Laboratory Fly Colonies

7. Potential of Hytrosaviruses as Bio-Pesticides
8. Strategies to Control GpSGHV in Laboratory Colonies of G. pallidipes
8.1. Immune-Intervention Strategies
8.2. Modifications of in vitro Membrane Feeding Regime
8.3. Implications of Successful Control of GpSGHV Infections in Colonized G. pallidipes
9. Concluding Remarks
Supplementary Files
Supplementary File 1Acknowledgments
Conflict of Interest
References and Notes
- Gooding, R.H.; Krafsur, E.S. Tsetse genetics: Contributions to biology, systematics, and control of tsetse flies. Annu. Rev. Entomol. 2005, 50, 101–123. [Google Scholar] [CrossRef]
- Mattioli, R.C.; Feldmann, U.; Hendrickx, G.; Wint, W.; Jannin, J.; Slingenbergh, J. Tsetse and trypanosomiasis intervention policies supporting sustainable animal-agricultural development. J. Food Agric. Environ. 2004, 2, 310–314. [Google Scholar]
- Mamoudou, A.; Zoli, A.; Hamadama, H.; Abah, S.; Geerts, S.; Clausen, P.H.; Zessin, K.H.; Kyule, M.; van den Bossche, P. Seasonal distribution and abundance of tsetse flies (Glossina spp.) in the Faro and Deo Division of the Adamaoua Plateau in Cameroon. Med. Vet. Entomol. 2008, 22, 32–36. [Google Scholar] [CrossRef]
- Smith, D.H.; Pepin, J.; Stich, A.H. Human African trypanosomiasis: An emerging public health crisis. Br. Med. Bull. 1998, 54, 341–355. [Google Scholar] [CrossRef]
- Cockerell, T.D.A. A fossil tsetse-fly in Colorado. Nature 1907, 76, 414. [Google Scholar] [CrossRef]
- Elsen, P.; Amoudi, M.A.; Leclercq, M. First record of Glossina fuscipes fuscipes Newstead, 1910 and Glossina morsitans submorsitans Newstead, 1910 in southwestern Saudi Arabia. Ann. Soc. Bel. Med. Trop. 1990, 70, 281–287. [Google Scholar]
- Hotez, P.J.; Kamath, A. Neglected tropical diseases in sub-saharan Africa: Review of their prevalence, distribution, and disease burden. PLoS Negl. Trop. Dis. 2009, 3, e412. [Google Scholar] [CrossRef]
- Hotez, P.J.; Molyneux, D.H.; Fenwick, A.; Kumaresan, J.; Sachs, S.E.; Sachs, J.D.; Savioli, L. Control of neglected tropical diseases. N. Engl. J. Med. 2007, 357, 1018–1027. [Google Scholar] [CrossRef]
- Cecchi, G.; Mattioli, R.C.; Slingenbergh, J.; de la Rocque, S. Land cover and tsetse fly distributions in sub-Saharan Africa. Med. Vet. Entomol. 2008, 22, 364–373. [Google Scholar] [CrossRef]
- Vreysen, M.J.B. Prospects for area-wide integrated control of tsetse flies (Diptera: Glossinidae) and trypanosomosis in sub-Saharan Africa. Rev. Soc. Entomol. Argent 2006, 65, 1–21. [Google Scholar]
- Otte, M.J.; Chilonda, P. Classification of cattle and small ruminant production systems in sub-Saharan Africa. Outlook Agr. 2003, 32, 183–190. [Google Scholar] [CrossRef]
- Budd, L.T. DFID-funded Tsetse and Trypanosomosis Research and Development Since 1980; Department of International Development: Aylesford, UK, 1999; Volume 2. [Google Scholar]
- Matovu, E.; Seebeck, T.; Enyaru, J.C.K.; Kaminsky, R. Drug resistance in Trypanosoma brucei spp., the causative agents of sleeping sickness in man and nagana in cattle. Microb. Infect. 2001, 3, 763–770. [Google Scholar] [CrossRef]
- Oberholzer, M.; Lopez, M.A.; McLelland, B.T.; Hill, K.L. Social motility in african trypanosomes. PLoS Pathog. 2010, 6, e1000739. [Google Scholar]
- Vickerman, K. Developmental cycles and biology of pathogenic trypanosomes. Br. Med. Bull. 1985, 41, 105–114. [Google Scholar]
- Brun, R.; Blum, J.; Chappuis, F.; Burri, C. Human African trypanosomiasis. Lancet 2010, 375, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Chappuis, F.; Loutan, L.; Simarro, P.; Lejon, V.; Büscher, P. Options for field diagnosis of human african trypanosomiasis. Clin. Microbiol. Rev. 2005, 18, 133–146. [Google Scholar] [CrossRef]
- Friedheim, E.A.H. Mel B in the treatment of human trypanosomiasis. Am. J. Trop. Med. Hyg. 1949, 29, 173–180. [Google Scholar]
- Burri, C.; Nkunku, S.; Merolle, A.; Smith, T.; Blum, J.; Brun, R. Efficacy of new, concise schedule for melarsoprol in treatment of sleeping sickness caused by Trypanosoma brucei gambiense: A randomised trial. Lancet 2000, 355, 1419–1425. [Google Scholar] [CrossRef]
- Burri, C. Chemotherapy against human African trypanosomiasis: Is there a road to success? Parasitology 2010, 137, 1987–1994. [Google Scholar] [CrossRef]
- Barrett, M.P.; Vincent, I.M.; Burchmore, R.J.; Kazibwe, A.J.; Matovu, E. Drug resistance in human African trypanosomiasis. Future Microbiol. 2011, 6, 1037–1047. [Google Scholar] [CrossRef]
- Geerts, S.; Holmes, P.H. Drug Management and Parasite Resistance in Bovine Trypanosomiasis in Africa; FAO: Rome, Italy, 1998. [Google Scholar]
- Schofield, C.J.; Kabayo, J.P. Trypanosomiasis vector control in Africa and Latin America. Parasit Vectors 2008, 1, 24. [Google Scholar] [CrossRef]
- Vreysen, M.J.B.; Seck, M.T.; Sall, B.; Bouyer, J. Tsetse flies: Their biology and control using area-wide integrated pest management approaches. J. Invertebr. Pathol. 2013, 112, S15–S25. [Google Scholar] [CrossRef]
- Abd-Alla, A.; Cousserans, F.; Parker, A.; Parker, N.; Robinson, A.; Bergoin, M. Characterization of a Novel virus from the Mass-reared Tsetse Fly Glossina pallidipes. In Proceedings of 11th meeting of the Working Group Arthropod Mass Rearing and Quality Control, Montreal, Canada, 28 October–1 November 2007; pp. 1–4.
- Abd-Alla, A.M.M.; Cousserans, F.; Parker, A.G.; Jehle, J.A.; Parker, N.J.; Vlak, J.M.; Robinson, A.S.; Bergoin, M. Genome analysis of a Glossina pallidipes salivary gland hypertrophy virus reveals a novel large double-stranded circular DNA virus. J. Virol. 2008, 82, 4595–4611. [Google Scholar] [CrossRef]
- Abd-Alla, A.M.M.; Vlak, J.M.; Bergoin, M.; Maruniak, J.E.; Parker, A.G.; Burand, J.P.; Jehle, J.A.; Boucias, D.G. Hytrosavirus Study Group of the ICTV Hytrosaviridae: A proposal for classification and nomenclature of a new insect virus family. Arch. Virol. 2009, 154, 909–918. [Google Scholar] [CrossRef]
- Abd-Alla, A.; Bossin, H.; Cousserans, F.; Parker, A.; Bergoin, M.; Robinson, A. Development of a non-destructive PCR method for detection of the salivary gland hypertrophy virus (SGHV) in tsetse flies. J. Virol. Methods 2007, 139, 143–149. [Google Scholar] [CrossRef]
- Leak, S.G.A. Tsetse Biology and Ecology: Their Role in the Epidemiology and Control of Trypanosomosis; CABI Publishing: Wallingford, CT, USA, 1998. [Google Scholar]
- Matthews, E.G.; Clyne, D.; Kitching, R.L. Insect Ecology, 2nd ed.; University of Queensland Press: St. Lucia, Qld, Australia, 1984; Volume 6. [Google Scholar]
- Dujardin, J.-P.; Schofield, C.J. Triatominae: Systematics, morphology and population biology. In The Trypanosomiases; Maudlin, I., Holmes, P.H., Miles, M.A., Eds.; CABI Publishing: Wallingford, CT, USA, 2004; pp. 181–201. [Google Scholar]
- Allsopp, R. Options for vector control against trypanonomiasis in Africa. Trends Parasitol. 2001, 17, 15–19. [Google Scholar] [CrossRef]
- Brightwell, B.; Dransfield, R. Odour attractants for tsetse: Glossina austeni, G. brevipalpis and G. swynnertoni. Med. Vet. Entomol. 1997, 11, 297–299. [Google Scholar] [CrossRef]
- Thomson, J.W.; Wilson, A. The Control of Tsetse Flies and Trypanosomiasis by the Application of Deltamethrin to Cattle. In proceedings of the 20th Meeting of the International Scientific Coubcil for the Trypanosomiasis Research and Control (ISCTRC), Mombasa, Kenya, 10–14 April 1989; pp. 450–454.
- Brightwell, B.; Dransfield, R.D.; Stevenson, P.; Williams, B. Changes over twelve years in populations of Glossina pallidipes and Glossina longipennis (Diptera: Glossinidae) subject to varying trapping pressure at Nguruman, sourth-west Kenya. Bull. Entomol. Res. 1997, 87, 349–370. [Google Scholar] [CrossRef]
- Knipling, E.F. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–469. [Google Scholar]
- Knipling, E.F. Sterile-male method of population control. Science 1959, 130, 902–904. [Google Scholar]
- Robinson, A.S. Mutations and their use in insect control. Mutat. Res. 2002, 511, 113–132. [Google Scholar] [CrossRef]
- Robinson, A.S. Genetic Basis of the Sterile Insect Technique. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 95–114. [Google Scholar]
- Abila, P.P.; Kiendrebeogo, M.; Mutika, G.N.; Parker, A.G.; Robinson, A.S. The effect of age on the mating competitiveness of male Glossina fuscipes fuscipes and G. palpalis palpalis. J. Insect Sci. 2003, 3, 13. [Google Scholar]
- Enkerlin, W.R. Impact of Fruit Fly Control Programmes Using the Sterile Insect Technique. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 651–676. [Google Scholar]
- Franz, G. Genetic Sexing Strains in Mediterranean Fruit Fly, An Example for Other Species Amenable to Large-scale Rearing as Required for the Sterile Insect Technique. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 427–451. [Google Scholar]
- Dyck, V.A.; Graham, S.H.; Bloem, K.A. Implementation of the Sterile Insect Release Programme to Eradicate the Codling Moth, Cydia pomonella (L.) (Lepidoptera: Olethreutidae), in British Columbia, Canada. In Proceedings of Management of Insect Pests: Nuclear and Related Molecular and Genetic Techniques, Vienna, Austria, 19–23 October 1992; pp. 285–297.
- Carpenter, J.; Bloem, S.; Hofmeyr, H. Area-wide Control Tactics for the False Codling Moth Thaumatotibia leucotreta in South Africa: A Potential Invasive Species. In Area-wide Control of Insect Pests: From Research to Field Implementation, 1st ed.; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 351–359. [Google Scholar]
- Daly, J.M.; Barrington, A.M.; Hackett, J.K.; Suckling, D.M. Sterilisation of painted apple moth Teia anartoides (Lepidoptera: Lymantridae) by irradiation. N. Z. Plant Prot. 2002, 55, 7–11. [Google Scholar]
- Koyama, J.; Kakinohana, H.; Miyatake, T. Eradication of the melon fly, Bactrocera cucurbitae, in Japan: Importance of behaviour, ecology, genetics and evolution. Ann. Rev. Entomol. 2004, 49, 331–349. [Google Scholar] [CrossRef]
- Vreysen, M.J.; Saleh, K.M.; Ali, M.Y.; Abdulla, A.M.; Zhu, Z.R.; Juma, K.G.; Dyck, V.A.; Msangi, A.R.; Mkonyi, P.A.; Feldmann, H.U. Glossina austeni (Diptera: Glossinidae) eradicated on the island of Unguja, Zanzibar, using the sterile insect technique. J. Econ. Entomol. 2000, 93, 123–135. [Google Scholar] [CrossRef]
- Vreysen, M.J.; Saleh, K.M.; Lancelot, R.; Bouyer, J. Factory tsetse flies must behave like wild flies: A prerequisite for the sterile insect technique. PLoS Negl. Trop. Dis. 2011, 5, e907. [Google Scholar] [CrossRef] [Green Version]
- Politzar, H.; Merot, P.; Brandl, F.E. Experimental aerial release of sterile males of Glossina palpalis gambiensis and Glossina tachinoides in a biological control operation. Rev. Élev. Méd. Vét. Pays Trop. 1984, 37, 198–202. [Google Scholar]
- Olandunmade, M.A.; Feldmann, U.; Takken, W.; Tanabe, S.O.; Hamann, H.J.; Onah, J.; Dengwat, L.; Vloedt, A.M.V.V.D.; Gingrich, R.E. Eradication of Glossina palpalis palpalis (Robineau-Desvoidy) (Diptera: Glossinidae) from agropastoral land in central Nigeria by means of the sterile insect technique. In Proceedings of the final research co-ordination meeting, Vom, Plateau State, Nigeria, 6–10 June 1988; pp. 5–23.
- Alemu, T.; Kapitano, B.; Mekonnen, S.; Aboset, G.; Kiflom, M.; Bancha, B.; Woldeyes, G.; Bekele, K.; Feldmann, U. Area-wide Control of Tsetse and Trypanosomosis: Ethiopian Experience in the Southern Rift Valley. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 325–335. [Google Scholar]
- Abd-Alla, A.M.M.; Kariithi, H.M.; Parker, A.G.; Robinson, A.S.; Kiflom, M.; Bergoin, M.; Vreysen, M.J.B. Dynamics of the salivary gland hypertrophy virus in laboratory colonies of Glossina pallidipes (Diptera: Glossinidae). Virus Res. 2010, 150, 103–110. [Google Scholar] [CrossRef]
- Abd-Alla, A.M.M.; Parker, A.G.; Vreysen, M.J.B.; Bergoin, M. Tsetse salivary gland hypertrophy virus: Hope or hindrance for tsetse control? PLoS Negl. Trop. Dis. 2011, 5, e1220. [Google Scholar] [CrossRef]
- Kariithi, H.M.; Ahmadi, M.; Parker, A.G.; Franz, G.; Ros, V.I.D.; Haq, I.; Elashry, A.M.; Vlak, J.M.; Bergoin, M.; Vreysen, M.J.B.; et al. Prevalence and genetic variation of salivary gland hypertrophy virus in wild populations of the tsetse fly Glossina pallidipes from southern and eastern Africa. J. Invertebr. Pathol. 2013, 112, S123–S132. [Google Scholar] [CrossRef]
- Whitnall, A.B.M. The trypanosome infections of Glossina pallidipes in the Umfolosi Game Reserve, Zululand. Onderstepoort J. Vet. Sci. Anim. Ind. 1934, 2, 2–21. [Google Scholar]
- Whitnall, A.B.M. The trypanosome infections of Glossina pallidipes in the Umfolosi Game Reserve, Zululand. (Preliminary Report); In 18th Report of the Director of Veterinary Services and Animal Industry; Union of South Africa: Onderstepoort, South Africa, 1932; pp. 7–21. [Google Scholar]
- Burtt, E. Hypertrophied salivary glands in Glossina: Evidence that G. pallidipes with this abnormality is particularly suited to trypanosome infection. Ann. Trop. Med. Parasitol. 1945, 39, 11–13. [Google Scholar]
- Jenni, L. Virus like particles in a strain of G. morsitans centralis, Machado 1970. Trans. R. Soc. Trop. Med. Hyg. 1973, 67, 295. [Google Scholar] [CrossRef]
- Jenni, L.; Steiger, R.F. Virus like particles in the tsetse fly, Glossina morsitans sspp. Preliminary results. Rev. Suisse Zool. 1974, 81, 663–666. [Google Scholar]
- Jenni, L.; Steiger, R. Virus like particles of Glossina fuscipes fuscipes Newst. 1910. Acta Trop. 1974, 31, 177–180. [Google Scholar]
- Jenni, L.; Böhringer, S. Nuclear coat and virus like particles in the midgut epithelium of Glossina morsitans sspp. Acta Trop. 1976, 33, 380–389. [Google Scholar]
- Lyon, J.P. La mouche des Narcisses (Merodon equestris F., Diptere Syrphidae). I. Identification de l'insecte et de ses degats et biologie dans le sud-est de la France. Rev. Zool. Agr. Pathol. Veg. 1973, 72, 65–92. [Google Scholar]
- Jaenson, T.G.T. Virus-like rods associated with salivary gland hyperplasia in tsetse, Glossina pallidipes. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 234–238. [Google Scholar] [CrossRef]
- Amargier, A.; Lyon, J.P.; Vago, C.; Meynadier, G.; Veyrunes, J.C. Mise en evidence et purification d'un virus dans la proliferation monstrueuse glandulaire d'insectes. Etude sur Merodon equestris F. (Diptere: Syrphidae). C. R. Acad. Sci. D 1979, 289, 481–484. [Google Scholar]
- Otieno, L.H.; Kokwaro, E.D.; Chimtawi, M.; Onyango, P. Prevalence of enlarged salivary glands in wild populations of Glossina pallidipes in Kenya, with a note on the ultrastructure of the affected organ. J. Invertebr. Pathol. 1980, 36, 113–118. [Google Scholar] [CrossRef]
- Opiyo, E.A.; Okumu, I. The KETRI Glossina pallidipes Colony: Further Observations. In Diseases of the Tropics; Tukei, P.M., Njogu, A.R., Eds.; Africa Book Services: Nairobi, Kenya, 1983; pp. 52–54. [Google Scholar]
- Odindo, M.O.; Sabwa, D.M.; Amutalla, P.A.; Otieno, W.A. Preliminary tests on the transmission of virus-like particles to the tsetse Glossina pallidipes. Insect Sci. Appl. 1981, 2, 219–221. [Google Scholar]
- Odindo, M.O. Incidence of salivary gland hypertrophy in field populations of the tsetse Glossina pallidipes on the South Kenya coast. Insect Sci. Appl. 1982, 3, 59–64. [Google Scholar]
- Odindo, M.O.; Payne, C.C.; Crook, N.E.; Jarret, P. Properties of a novel DNA virus from the tsetse fly, Glossina pallidipes. J. Gen. Virol. 1986, 67, 527–536. [Google Scholar] [CrossRef]
- Odindo, M.O.; Amutalla, P.A. Distribution pattern of the virus of Glossina pallidipes Austen in a forest ecosystem. Insect Sci. Appl. 1986, 7, 79–84. [Google Scholar]
- Jaenson, T.G.T. Sex ratio distortion and reduced lifespan of Glossina pallidipes infected with the virus causing salivary gland hyperplasia. Entomol. Exp. Appl. 1986, 41, 256–271. [Google Scholar]
- Ellis, D.S.; Maudlin, I. Salivary gland hyperplasia in wild caught tsetse from Zimbabwe. Entomol. Exp. Appl. 1987, 45, 167–173. [Google Scholar] [CrossRef]
- Gouteux, J.P. Prevalence of enlarged salivary glands in Glossina palpalis, G. pallicera, and G. nigrofusca (Diptera: Glossinidae) from the Vavoua area, Ivory Coast. J. Med. Entomol. 1987, 24, 268. [Google Scholar]
- Odindo, M.O. Glossina pallidipes virus: Its potential for use in biological control of tsetse. Insect Sci. Appl. 1988, 9, 399–403. [Google Scholar]
- Jura, W.G.Z.O.; Odhiambo, T.R.; Otieno, L.H.; Tabu, N.O. Gonadal lesions in virus-infected male and female tsetse, Glossina pallidipes (Diptera: Glossinidae). J. Invertebr. Pathol. 1988, 52, 1–8. [Google Scholar] [CrossRef]
- Jura, W.G.Z.O.; Zdarek, J.; Otieno, L.H. A simple method for artificial infection of tsetse, Glossina morsitans morsitans larvae with the DNA virus of G. pallidipes. Int. J. Trop. Insect Sci. 1993, 14, 383–387. [Google Scholar] [CrossRef]
- Jura, W.G.Z.O.; Davies-Cole, J.O.A. Some aspects of mating behavior of Glossina morsitans morsitans males infected with a DNA virus. Biol. Control 1992, 2, 188–192. [Google Scholar] [CrossRef]
- Jura, W.; Otieno, L.; Chimtawi, M. Ultrastructural evidence for trans-ovum transmission of the DNA virus of tsetse, Glossina pallidipes (Diptera: Glossinidae). Curr. Microbiol. 1989, 18, 1–4. [Google Scholar] [CrossRef]
- Kokwaro, E.D.; Nyindo, M.; Chimtawi, M. Ultrastructural changes in salivary glands of tsetse, Glossina morsitans morsitans, infected with virus and rickettsia-like organisms. J. Invertebr. Pathol. 1990, 56, 337–346. [Google Scholar] [CrossRef]
- Kokwaro, E.D.; Otieno, L.H.; Chimtawi, M. Salivary glands of the tsetse Glossina pallidipes Austen infected with Trypanosoma brucei and virus particles: Ultrastructural study. Int. J. Trop. Insect Sci. 1991, 12, 661–669. [Google Scholar]
- Shaw, M.K.; Moloo, S.K. Virus-like particles in Rickettsia within the midgut epithelial cells of Glossina morsitans centralis and Glossina brevipalpis. J. Invertebr. Pathol. 1993, 61, 162–166. [Google Scholar] [CrossRef]
- Coler, R.R.; Boucias, D.G.; Frank, J.H.; Maruniak, J.E.; Garcia-Canedo, A.; Pendland, J.C. Characterization and description of a virus causing salivary gland hyperplasia in the housefly, Musca domestica. Med. Vet. Entomol. 1993, 7, 275–282. [Google Scholar] [CrossRef]
- Sang, R.C.; Jura, W.G.Z.O.; Otieno, L.H.; Ogaja, P. Ultrastructural changes in the milk gland of tsetse Glossina morsitans centralis (Diptera; Glissinidae) female infected by a DNA virus. J. Invertebr. Pathol. 1996, 68, 253–259. [Google Scholar] [CrossRef]
- Sang, R.C.; Jura, W.G.Z.O.; Otieno, L.H.; Tukei, P.M.; Mwangi, R.W. Effects of tsetse DNA virus infection on the survival of a host fly Glossina morsitans centralis (Diptera: Glossinidae). J. Invertebr. Pathol. 1997, 69, 253–260. [Google Scholar] [CrossRef]
- Sang, R.C.; Jura, W.G.Z.O.; Otieno, L.H.; Mwangi, R.W. The effects of a DNA virus infection on the reproductive potential of female tsetse flies, Glossina morsitans centralis and Glossina morsitans morsitans (Diptera: Glossinidae). Mem. Inst. Oswaldo Cruz. 1998, 93, 861–864. [Google Scholar] [CrossRef]
- Sang, R.C.; Jura, W.G.Z.O.; Otieno, L.H.; Mwangi, R.W.; Ogaja, P. The effects of a tsetse DNA virus infection on the functions of the male accessory reproductive gland in the host fly Glossina morsitans centralis (Diptera; Glossinidae). Curr. Microbiol. 1999, 38, 349–354. [Google Scholar]
- Kokwaro, E.D. Virus particles in male accessory reproductive glands of tsetse, Glossina morsitans morsitans (Diptera: Glossinidae) and associated tissue changes. Int. J. Trop. Insect Sci. 2006, 26, 266–272. [Google Scholar] [CrossRef]
- Salem, T.Z.; Garcia-Maruniak, A.; Lietze, V.U.; Maruniak, J.E.; Boucias, D.G. Analysis of transcripts from predicted open reading frames of Musca domestica salivary gland hypertrophy virus. J. Gen. Virol. 2009, 90, 1270–1280. [Google Scholar] [CrossRef]
- Kariithi, H.M.; van Lent, J.W.; Boeren, S.; Abd-Alla, A.M.; Ince, I.A.; van Oers, M.M.; Vlak, J.M. Correlation between structure, protein composition, morphogenesis and cytopathology of Glossina pallidipes salivary gland hypertrophy virus. J Gen. Virol. 2013, 94, 193–208. [Google Scholar] [CrossRef]
- Kariithi, H.M.; Ince, I.A.; Boeren, S.; Vervoort, J.; Bergoin, M.; van Oers, M.M.; Abd-Alla, A.M.M.; Vlak, J.M. Proteomic analysis of Glossina pallidipes salivary gland hypertrophy virus virions for immune intervention in tsetse fly colonies. J. Gen. Virol. 2010, 91, 3065–3074. [Google Scholar] [CrossRef]
- Prompiboon, P.; Lietze, V.-U.; Denton, J.S.S.; Geden, C.J.; Steenberg, T.; Boucias, D.G. Musca domestica salivary gland hypertrophy virus, a globally distributed insect virus that infects and sterilizes female houseflies. Appl. Environ. Microbiol. 2010, 76, 994–998. [Google Scholar] [CrossRef]
- Luo, L.; Zeng, L. A new rod-shaped virus from parasitic wasp Diachasmimorpha longicaudata (Hymenoptera: Braconidae). J. Invertebr. Pathol. 2010, 103, 165–169. [Google Scholar] [CrossRef]
- Boucias, D.G.; Kariithi, H.M.; Bourtzis, K.; Schneider, D.I.; Kelley, K.; Miller, W.J.; Parker, A.G.; Abd-Alla, A.M.M. Trans-generational transmission of the Glossina pallidipes hytrosavirus depends on the presence of a functional symbiome. PLoS One 2013, 8, e61150. [Google Scholar]
- Abd-Alla, A.M.M.; Kariithi, H.M.; Mohamed, A.H.; Lapiz, E.; Parker, A.G.; Vreysen, M.J.B. Managing hytrosavirus infections in Glossina pallidipes colonies: Feeding regime affects the prevalence of salivary gland hypertrophy syndrome. PLoS One 2013, 8, e61875. [Google Scholar]
- Howatson, A.; Whitmore, G. The development and structure of vesicular stomatitis virus. Virology 1962, 16, 466–478. [Google Scholar] [CrossRef]
- Mussgay, M.; Weibel, J. Electron microscopic studies on the development oof vesicular stomatis virus in KB cells. J. Cell Biol. 1963, 16, 119–129. [Google Scholar] [CrossRef]
- Chamberlain, R.W. Arboviruses, the arthropod-borne animal viruses. Curr. Top. Microbiol. Immunol. 1968, 42, 38–58. [Google Scholar] [CrossRef]
- Janzen, H.G.; Rhodes, A.J.; Doane, F.W. Chikungunya virus in salivary glands of Aedes aegypti (L.): An electron microscope study. Can. J. Microbiol. 1970, 16, 581–586. [Google Scholar] [CrossRef]
- Lamotte, L.C., Jr. Japanese B encephalitis virus in the organs of infected mosquitoes. Am. J. Epidemiol. 1960, 72, 73–87. [Google Scholar]
- Mims, C.A.; Day, M.F.; Marshall, I.D. Cytopathic effect of Semliki Forest virus in the mosquito, Aedes aegypti. Am. J. Trop. Med. Hyg. 1966, 15, 775–784. [Google Scholar]
- Longdon, B.; Wilfert, L.; Obbard, D.J.; Jiggins, F.M. Rhabdoviruses in two species of Drosophila: Vertical transmission and a recent sweep. Genetics 2011, 188, 141–150. [Google Scholar] [CrossRef]
- L'Heritier, P.H. The Hereditary virus of Drosophila. In Advances in Virus Research; Kenneth, M.S., Lauffer, M.A., Eds.; Academic Press Inc.: New York, NY, USA, 1958; Volume 5, pp. 195–245. [Google Scholar]
- Anderson, R.M.; May, R.M. Infectious diseases and population cycles of forest insects. Science 1980, 210, 658–661. [Google Scholar]
- Geden, C.J.; Lietze, V.U.; Boucias, D.G. Seasonal prevalence and transmission of salivary gland hypertrophy virus of house flies (Diptera: Muscidae). J. Med. Entomol. 2008, 45, 42–51. [Google Scholar] [CrossRef]
- Lietze, V.U.; Geden, C.J.; Blackburn, P.; Boucias, D.G. Effects of salivary gland hypertrophy virus on the reproductive behavior of the housefly, Musca domestica. Appl. Environ. Microbiol. 2007, 73, 6811–6818. [Google Scholar] [CrossRef]
- Geden, C.; Garcia-Maruniak, A.; Lietze, V.U.; Maruniak, J.; Boucias, D.G. Impact of house fly salivary gland hypertrophy virus (MdSGHV) on a heterologous host, Stomoxys calcitrans. J. Med. Entomol. 2011, 48, 1128–1135. [Google Scholar] [CrossRef]
- Chou, M.-Y.; Haymer, D.S.; Feng, H.-Y.; Mau, R.F.L.; Hsu, J.-C. Potential for insecticide resistance in populations of Bactrocera dorsalis in Hawaii: Spinosad susceptibility and molecular characterization of a gene associated with organophosphate resistance. Entomol. Exp. Appl. 2010, 134, 296–303. [Google Scholar]
- Lawrence, P.O.; Akin, D. Virus-like particles from the poison glands of the parasitic wasp Biosteres longicaudatus (Hymenoptera: Braconidae). Can. J. Zool. 1990, 68, 539–546. [Google Scholar] [CrossRef]
- Lawrence, P.O. Morphogenesis and cytopathic effects of the Diachasmimorpha longicaudata entomopoxvirus in host haemocytes. J. Insect Physiol. 2005, 51, 221–233. [Google Scholar] [CrossRef]
- Abd-Alla, A.; Cousserans, F.; Parker, A.; Bergoin, M.; Chiraz, J.; Robinson, A. Quantitative PCR analysis of the salivary gland hypertrophy virus (GpSGHV) in a laboratory colony of Glossina pallidipes. Virus Res. 2009, 139, 48–53. [Google Scholar] [CrossRef]
- Lietze, V.-U.; Abd-Alla, A.; Vreysen, M.; Geden, C.C.; Boucias, D.G. Salivary gland hypertrophy viruses: A novel group of insect pathogenic viruses. Annu. Rev. Entomol. 2011, 56, 63–80. [Google Scholar] [CrossRef]
- Duron, O.; Bouchon, D.; Boutin, S.; Bellamy, L.; Zhou, L.; Engelstadter, J.; Hurst, G.D. The diversity of reproductive parasites among arthropods: Wolbachia do not walk alone. BMC Biol. 2008, 6, 27. [Google Scholar] [CrossRef]
- Engelstadter, J.; Hurst, G.D.D. The ecology and evolution of microbes that manipulate host reproduction. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 127–149. [Google Scholar] [CrossRef]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?—A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef]
- Doudoumis, V.; Tsiamis, G.; Wamwiri, F.; Brelsfoard, C.; Alam, U.; Aksoy, E.; Dalaperas, S.; bd-Alla, A.; Ouma, J.; Takac, P.; et al. Detection and characterization of Wolbachia infections in laboratory and natural populations of different species of tsetse flies (genus Glossina). BMC Microbiol. 2012, 12, S3. [Google Scholar] [CrossRef]
- Alam, U.; Hyseni, C.; Symula, R.E.; Brelsfoard, C.; Wu, Y.; Kruglov, O.; Wang, J.; Echodu, R.; Alioni, V.; Okedi, L.M.; et al. Implications of microfauna-host interactions for trypanosome transmission dynamics in Glossina fuscipes fuscipes in Uganda. Appl. Environ. Microbiol. 2012, 78, 4627–4637. [Google Scholar] [CrossRef]
- Hughes, G.L.; Ren, X.; Ramirez, J.L.; Sakamoto, J.M.; Bailey, J.A.; Jedlicka, A.E.; Rasgon, J.L. Wolbachia infections in Anopheles gambiae cells: Transcriptomic characterization of a novel host-symbiont interaction. PLoS Pathog. 2011, 7, e1001296. [Google Scholar] [CrossRef]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef]
- Song, J.; Wang, R.; Deng, F.; Wang, H.; Hu, Z. Functional studies of per os infectivity factors of Helicoverpa armigera single nucleocapsid nucleopolyhedrovirus. J. Gen. Virol. 2008, 89, 2331–2338. [Google Scholar] [CrossRef]
- Garcia-Maruniak, A.; Maruniak, J.E.; Farmerie, W.; Boucias, D.G. Sequence analysis of a non-classified, non-occluded DNA virus that causes salivary gland hypertrophy of Musca domestica, MdSGHV. Virology 2008, 377, 184–196. [Google Scholar] [CrossRef]
- Cheng, C.H.; Liu, S.M.; Chow, T.Y.; Hsiao, Y.Y.; Wang, D.P.; Huang, J.J.; Chen, H.H. Analysis of the complete genome sequence of the Hz-1 virus suggests that it is related to members of the Baculoviridae. J. Virol. 2002, 76, 9024–9034. [Google Scholar] [CrossRef]
- Wang, Y.; Kleespies, R.G.; Huger, A.M.; Jehle, J.A. The genome of Gryllus bimaculatus nudivirus indicates an ancient diversification of baculovirus-related nonoccluded nudiviruses of insects. J. Virol. 2007, 81, 5395–5406. [Google Scholar] [CrossRef]
- Wang, Y.; van Oers, M.M.; Crawford, A.M.; Vlak, J.M.; Jehle, J.A. Genomic analysis of Oryctes rhinoceros virus reveals genetic relatedness to Heliothis zea virus 1. Arch. Virol. 2006, 152, 519–531. [Google Scholar]
- Wang, Y.; Bininda-Emonds, O.; van Oers, M.; Vlak, J.; Jehle, J. The genome of Oryctes rhinoceros nudivirus provides novel insight into the evolution of nuclear arthropod-specific large circular double-stranded DNA viruses. Virus Genes 2011, 42, 444–456. [Google Scholar] [CrossRef]
- Ohkawa, T.; Washburn, J.O.; Sitapara, R.; Sid, E.; Volkman, L.E. Specific binding of Autographa californica M nucleopolyhedrovirus occlusion-derived virus to midgut cells of Heliothis virescens larvae is mediated by products of pif genes Ac119 and Ac022 but not by Ac115. J. Virol. 2005, 79, 15258–15264. [Google Scholar] [CrossRef]
- Blissard, G.W.; Rohrmann, G.F. Baculovirus diversity and molecular biology. Ann. Rev. Entomol. 1990, 35, 127–155. [Google Scholar] [CrossRef]
- Tidona, C.A.; Darai, G. Iridovirus homologues of cellular genes—Implications for the molecular evolution of large DNA viruses. Virus Genes 2000, 21, 77–81. [Google Scholar] [CrossRef]
- Invertebrate Virus Subcommittee, Hytrosavirus Study Group. Available online: http://www.ictvonline.org/taxonomyHistory.asp?taxnode_id=20125369&taxa_name=Glossinavirus/ (accessed on 19 June 2013).
- Garcia-Maruniak, A.; Abd-Alla, A.M.M.; Salem, T.Z.; Parker, A.G.; van Oers, M.M.; Maruniak, J.E.; Kim, W.; Burand, J.P.; Cousserans, F.; Robinson, A.S.; et al. Two viruses that cause salivary gland hypertrophy in Glossina pallidipes and Musca domestica are related and form a distinct phylogenetic clade. J. Gen. Virol. 2009, 90, 334–346. [Google Scholar] [CrossRef]
- Jaenson, T.G.T. Reproductive Biology of the Tsetse Glossina pallidipes Austen (Diptera, Glossinidae) with Special Reference to Mating Behaviour; Department of entomology, University of Uppsala: Uppsala, Sweden, 1978; p. 39. [Google Scholar]
- Lietze, V.U.; Sims, K.R.; Salem, T.Z.; Geden, C.J.; Boucias, D.G. Transmission of MdSGHV among adult house flies, Musca domestica (Diptera: Muscidae), occurs via oral secretions and excreta. J. Invertebr. Pathol. 2009, 101, 49–55. [Google Scholar] [CrossRef]
- Mutika, G.N.; Marin, C.; Parker, A.G.; Boucias, D.G.; Vreysen, M.J.; Abd-Alla, A.M. Impact of salivary gland hypertrophy virus infection on the mating success of male Glossina pallidipes: Consequences for the sterile insect technique. PLoS One 2012, 7, e42188. [Google Scholar]
- Boucias, D.G.; Deng, F.; Hu, Z.; Garcia-Maruniak, A.; Lietze, V.U. Analysis of the structural proteins from the Musca domestica hytrosavirus with an emphasis on the major envelope protein. J. Invertebr. Pathol. 2013, 112, S44–S52. [Google Scholar] [CrossRef]
- Lietze, V.U.; Abd-Alla, A.M.M.; Boucias, D.G. Two hytrosaviruses, MdSGHV and GpSGHV, induce distinct cytopathologies in their respective host insects. J. Invertebr. Pathol. 2011, 107, 161–163. [Google Scholar] [CrossRef]
- Gao, Y.; Luo, L. Genome-based phylogeny of dsDNA viruses by a novel alignment-free method. Gene 2012, 492, 309–314. [Google Scholar] [CrossRef]
- Wu, G.A.; Jun, S.R.; Sims, G.E.; Kim, S.H. Whole-proteome phylogeny of large dsDNA virus families by an alignment-free method. Proc. Natl. Acad. Sci. USA 2009, 106, 12826–12831. [Google Scholar] [CrossRef]
- Yu, Z.G.; Chu, K.H.; Li, C.P.; Anh, V.; Zhou, L.Q.; Wang, R.W. Whole-proteome phylogeny of large dsDNA viruses and parvoviruses through a composition vector method related to dynamical language model. BMC Evol. Biol. 2010, 10, 192. [Google Scholar] [Green Version]
- Wang, Y.; Jehle, J.A. Nudiviruses and other large, double-stranded circular DNA viruses of invertebrates: New insights on an old topic. J. Invertebr. Pathol. 2009, 101, 187–193. [Google Scholar] [CrossRef]
- Jehle, J.A.; Abd-Alla, A.M.; Wang, Y. Phylogeny and evolution of Hytrosaviridae. J. Invertebr. Pathol. 2013, 112, S62–S67. [Google Scholar] [CrossRef]
- Lodish, H.; Berk, A.; Zipursky, S.L.; Matsudaira, P.; Baltimore, D.; Darnell, J. Mutations: Types and Causes. In Molecular Cell Biology, 4th ed.; W. H. Freeman and Company: New York, NY, USA, 2000. [Google Scholar]
- Joyce, A.R.; Palsson, B.O. The model organism as a system: Integrating “omics” data sets. Nat. Rev. Mol. Cell Biol. 2006, 7, 198–210. [Google Scholar] [CrossRef]
- Kariithi, H.M.; Ince, I.A.; Boeren, S.; Abd-Alla, A.M.M.; Parker, A.G.; Aksoy, S.; Vlak, J.M.; van Oers, M.M. The salivary secretome of the tsetse fly, Glossina pallidipes (Diptera: Glossinidae) infected by salivary gland hypertrophy virus. PLoS Negl. Trop. Dis. 2011, 5, e1371. [Google Scholar] [CrossRef]
- Lee, J.C.; Chen, H.H.; Chao, Y.C. Persistent baculovirus infection results from deletion of the apoptotic suppressor gene p35. J. Virol. 1998, 72, 9157–9165. [Google Scholar]
- Geden, C.J.; Steenberg, T.; Lietze, V.U.; Boucias, D.G. Salivary gland hypertrophy virus of house flies in Denmark: Prevalence, host range, and comparison with a Florida isolate. J. Vector. Ecol. 2011, 36, 231–238. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Weidner, E.H.; Richter, A.R. Polyhedra without virions in a vertically transmitted nuclear polyhedrosis virus. J. Invertebr. Pathol. 1992, 60, 53–58. [Google Scholar]
- Hughes, D.S.; Possee, R.D.; King, L.A. Activation and detection of a latent baculovirus resembling Mamestra brassicae nuclear polyhedrosis virus in M. brassicae insects. Virology 1993, 194, 608–615. [Google Scholar] [CrossRef]
- Hughes, D.S.; Possee, R.D.; King, L.A. Evidence for the presence of a low-level, persistent baculovirus infection of Mamestra brassicae insects. J. Gen. Virol. 1997, 78, 1801–1805. [Google Scholar]
- Murillo, R.; Hussey, M.S.; Possee, R.D. Evidence for covert baculovirus infections in a Spodoptera exigua laboratory culture. J. Gen. Virol. 2011, 92, 1061–1070. [Google Scholar] [CrossRef] [Green Version]
- Bronkhorst, A.W.; van Cleef, K.W.; Vodovar, N.; Ince, I.A.; Blanc, H.; Vlak, J.M.; Saleh, M.C.; van Rij, R.P. The DNA virus invertebrate iridescent virus 6 is a target of the Drosophila RNAi machinery. Proc. Natl. Acad. Sci. USA. 2012, 109, E3604–E3613. [Google Scholar]
- Jayachandran, B.; Hussain, M.; Asgari, S. RNA interference as a cellular defense mechanism against the DNA virus baculovirus. J. Virol. 2012, 86, 13729–13734. [Google Scholar] [CrossRef]
- Wu, Y.L.; Wu, C.P.; Liu, C.Y.; Hsu, P.W.; Wu, E.C.; Chao, Y.C. A non-coding RNA of insect HzNV-1 virus establishes latent viral infection through microRNA. Sci. Rep. 2011, 1, 1–10. [Google Scholar]
- Van Den Abbeele, J.; Bourtzis, K.; Weiss, B.; Cordön-Rosales, C.; Miller, W.; Abd-Alla, A.M.M.; Parker, A. Enhancing tsetse fly refractoriness to trypanosome infection—A new IAEA coordinated research project. J. Invertebr. Pathol. 2013, 112, S142–S147. [Google Scholar] [CrossRef]
- Strand, M.R. Polydnaviruses. In Insect Virology; Asgari, S., Johnson, K.N., Eds.; Caister Academic Press: Norfolk, UK, 2010; pp. 171–197. [Google Scholar]
- Terzian, C.; Pelisson, A.; Bucheton, A. When Drosophila Meets Retrovirology: The Gypsy Case. In Transposons and the Dynamic Genome; Lankenau, D.H., Volff, J.N., Eds.; Springer-Verlag Berlin Heidelberg: London, UK, 2009; pp. 95–107. [Google Scholar]
- Varaldi, J.; Ravallec, M.; Labrosse, C.; Lopez-Ferber, M.; Bouletreau, M.; Fleury, F. Artifical transfer and morphological description of virus particles associated with superparasitism behaviour in a parasitoid wasp. J. Insect Physiol. 2006, 52, 1202–1212. [Google Scholar] [CrossRef]
- Varaldi, J.; Fouillet, P.; Ravallec, M.; Lopez-Ferber, M.; Bouletreau, M.; Fleury, F. Infectious behavior in a parasitoid. Science 2003, 302, 1930. [Google Scholar] [CrossRef]
- Rosen, L. Mechanism of vertical transmission of the dengue virus in mosquitoes. C. R. Acad. Sci. III 1987, 304, 347–350. [Google Scholar]
- Capinera, J.L. Habita. In Encyclopedia of Entomology, 2nd ed.; Capinera, J.L., Ed.; Springer Science: Dordrecht, The Netherlands, 2008; pp. 1761–1906. [Google Scholar]
- Lietze, V.U.; Geden, C.J.; Doyle, M.A.; Boucias, D.G. Disease dynamics and persistence of Musca domestica salivary gland hypertrophy virus infections in laboratory house fly (Musca domestica) populations. Appl. Environ. Microbiol. 2012, 78, 311–317. [Google Scholar] [CrossRef]
- Walshe, D.P.; Lehane, M.J.; Haines, L.R. Post eclosion age predicts the prevalence of midgut trypanosome infections in Glossina. PLoS One 2011, 6, e26984. [Google Scholar] [CrossRef]
- Bignell, D.E. The Arthropod Gut as an Environment for Microorganisms. In Microbial Ecology of the Gut; Clarke, R.T.J., Bauchop, T., Eds.; Academic Press Inc.: New York, NY, USA, 1983; pp. 205–227. [Google Scholar]
- Zhuzhikov, D.P. Function of the peritrophic membrane in Musca domestica L. and Calliphora erythrocephala Meig. J. Insect Physiol. 1964, 10, 273–278. [Google Scholar] [CrossRef]
- Chapman, R.F. Circulatory System, Blood and Immune Systems. In The Insects: Structure and Functions, 4th ed.; Simpson, S.J., Douglas, A.E., Eds.; Cambridge University Press: Cambridge, NY, USA, 1998; pp. 94–127. [Google Scholar]
- Feng, L.C. The role of the peritrophic membrane in Leishmania and Trypanosome infections of sandflies. Peking Nat. Hist. Bull. 1951, 19, 327–334. [Google Scholar]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New insights into peritrophic matrix synthesis, architecture, and function. Annu. Rev. Entomol. 2009, 54, 285–302. [Google Scholar] [CrossRef]
- Blackburn, K.; Wallbanks, K.R.; Molyneux, D.H.; Lavin, D.R.; Winstanley, S.L. The peritrophic membrane of the female sandfly Phlebotomus papatasi. Ann. Trop. Med. Parasitol. 1988, 82, 613–619. [Google Scholar]
- Bertram, D.S.; Bird, R.G. Studies on mosquito-borne viruses in their vector. I. The normal fine structure of the midgut epithelium of the adult female Aedes aegypti (L). and the functional significance of its modifications following a bloodmeal. Trans. R. Soc. Trop. Med. Hyg. 1961, 55, 404–423. [Google Scholar] [CrossRef]
- Chamberlain, R.W.; Sudia, W.D. Mechanism of transmission of viruses by mosquitoes. Annu. Rev. Entomol. 1961, 6, 371–390. [Google Scholar] [CrossRef]
- Gemetchu, T. The morphology and fine structure of the midgut and peritrophic membrane of the adult female, Phlebotomus longipes Parrot and Martin (Diptera: Psychodidae). Ann. Trop. Med. Parasitol. 1974, 68, 111–124. [Google Scholar]
- Mitsuhashi, W.; Kawakita, H.; Murakami, R.; Takemoto, Y.; Saiki, T.; Miyamoto, K.; Wada, S. Spindles of an entomopoxvirus facilitate its infection of the host insect by disrupting the peritrophic membrane. J. Virol. 2007, 81, 4235–4243. [Google Scholar] [CrossRef]
- Wang, P.; Granados, R.R. Molecular structure of the peritrophic membrane (PM): Identification of potential PM target sites for insect control. Arch. Insect Biochem. Physiol. 2001, 47, 110–118. [Google Scholar] [CrossRef]
- Tsai, Y.L.; Hayward, R.E.; Langer, R.C.; Fidock, D.A.; Vinetz, J.M. Disruption of Plasmodium falciparum chitinase markedly impairs parasite invasion of mosquito midgut. Infect. Immun. 2001, 69, 4048–4054. [Google Scholar] [CrossRef]
- Li, F.; Patra, K.P.; Vinetz, J.M. An anti-Chitinase malaria transmission-blocking single-chain antibody as an effector molecule for creating a Plasmodium falciparum-refractory mosquito. J. Infect. Dis. 2005, 192, 878–887. [Google Scholar] [CrossRef]
- Ejezie, G.C.; Davey, K.G. Some effects of mating in female tsetse, Glossina austeni Newst. J. Exp. Zool. 1977, 200, 303–310. [Google Scholar] [CrossRef]
- Harrison, R.; Hoover, K. Baculoviruses and other occluded viruses. In Insect Pathology, 2nd ed.; Vega, E.F., Kaya, H.K., Eds.; Elsevier: New York, NY, USA, 2012; pp. 73–131. [Google Scholar]
- Moscardi, F. Assessment of the application of baculoviruses for control of Lepidoptera. Annu. Rev. Entomol. 1999, 44, 257–289. [Google Scholar] [CrossRef]
- Horton, H.M.; Burand, J.P. Saturable attachment sites for polyhedron-derived baculovirus on insect cells and evidence for entry via direct membrane fusion. J. Virol. 1993, 67, 1860–1868. [Google Scholar]
- Sparks, W.O.; Rohlfing, A.; Bonning, B.C. A peptide with similarity to baculovirus ODV-E66 binds the gut epithelium of Heliothis virescens and impedes infection with Autographa californica multiple nucleopolyhedrovirus. J. Gen. Virol. 2011, 92, 1051–1060. [Google Scholar] [CrossRef]
- Sparks, W.O.; Harrison, R.L.; Bonning, B.C. Autographa californica multiple nucleopolyhedrovirus ODV-E56 is a per os infectivity factor, but is not essential for binding and fusion of occlusion-derived virus to the host midgut. Virology 2011, 409, 69–76. [Google Scholar] [CrossRef]
- Abd-Alla, A.M.; Adun, H.; Parker, A.G.; Vreysen, M.J.; Bergoin, M. The antiviral drug valacyclovir successfully suppresses salivary gland hypertrophy virus (SGHV) in laboratory colonies of Glossina pallidipes. PLoS One 2012, 7, e38417. [Google Scholar]
- Feldmann, U. Guidelines for the rearing of tsetse flies using the membrane feeding technique. In Techniques of Insect Rearing for the Development of Integrated Pest and Vector Management Strategies; Ochieng'-Odero, J.P.R., Ed.; ICIPE Science Press: Nairobi, Kenya, 1994; pp. 449–471. [Google Scholar]
- Enserink, M. Welcome to Ethiopia’s fly factory. Science 2007, 317, 310–313. [Google Scholar] [CrossRef]
- Malele, I.I.; Manangwa, O.; Nyingilili, H.H.; Kitwika, W.A.; Lyaruu, E.A.; Msangi, A.R.; Ouma, J.O.; Nkwangulila, G.; Abd-Alla, A.M.M. Prevalence of SGHV among tsetse species of economic importance in Tanzania and their implication for SIT application. J. Invertebr. Pathol. 2013, 112, S133–S137. [Google Scholar] [CrossRef]
- Abd-Alla, A.M.M.; Arif, B. Foreword. J. Invertebr. Pathol. 2013, 112, S1. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kariithi, H.M.; Van Oers, M.M.; Vlak, J.M.; Vreysen, M.J.B.; Parker, A.G.; Abd-Alla, A.M.M. Virology, Epidemiology and Pathology of Glossina Hytrosavirus, and Its Control Prospects in Laboratory Colonies of the Tsetse Fly, Glossina pallidipes (Diptera; Glossinidae). Insects 2013, 4, 287-319. https://doi.org/10.3390/insects4030287
Kariithi HM, Van Oers MM, Vlak JM, Vreysen MJB, Parker AG, Abd-Alla AMM. Virology, Epidemiology and Pathology of Glossina Hytrosavirus, and Its Control Prospects in Laboratory Colonies of the Tsetse Fly, Glossina pallidipes (Diptera; Glossinidae). Insects. 2013; 4(3):287-319. https://doi.org/10.3390/insects4030287
Chicago/Turabian StyleKariithi, Henry M., Monique M. Van Oers, Just M. Vlak, Marc J. B. Vreysen, Andrew G. Parker, and Adly M. M. Abd-Alla. 2013. "Virology, Epidemiology and Pathology of Glossina Hytrosavirus, and Its Control Prospects in Laboratory Colonies of the Tsetse Fly, Glossina pallidipes (Diptera; Glossinidae)" Insects 4, no. 3: 287-319. https://doi.org/10.3390/insects4030287
APA StyleKariithi, H. M., Van Oers, M. M., Vlak, J. M., Vreysen, M. J. B., Parker, A. G., & Abd-Alla, A. M. M. (2013). Virology, Epidemiology and Pathology of Glossina Hytrosavirus, and Its Control Prospects in Laboratory Colonies of the Tsetse Fly, Glossina pallidipes (Diptera; Glossinidae). Insects, 4(3), 287-319. https://doi.org/10.3390/insects4030287