
Interactions between the Multicolored Asian Lady Beetle Harmonia axyridis and the Parasitoid Dinocampus coccinellae


 , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Survey
2.2. Insect Rearing
2.3. Effect of Exposure Time on the Development and Yields of D. Coccinellae in H. Axyridis
2.4. Effects of Exposure to D. Coccinellae on H. Axyridis Longevity, Oviposition and Egg Fertility
2.4.1. Male Exposure
2.4.2. Female Exposure
2.5. Effects of D. Coccinellae on H. Axyridis vs. A. Bipunctata
2.5.1. Acceptance and Suitability
2.5.2. Effects on Longevity, Oviposition and Egg Fertility
2.6. Data Analysis
3. Results
3.1. Field Survey
3.2. Effect of Exposure Time on the Development and Yields of D. Coccinellae in H. Axyridis
3.3. Effects of Exposure to D. coccinellae on H. axyridis Longevity, Oviposition and Egg Fertility
3.3.1. Male Exposure
3.3.2. Female Exposure
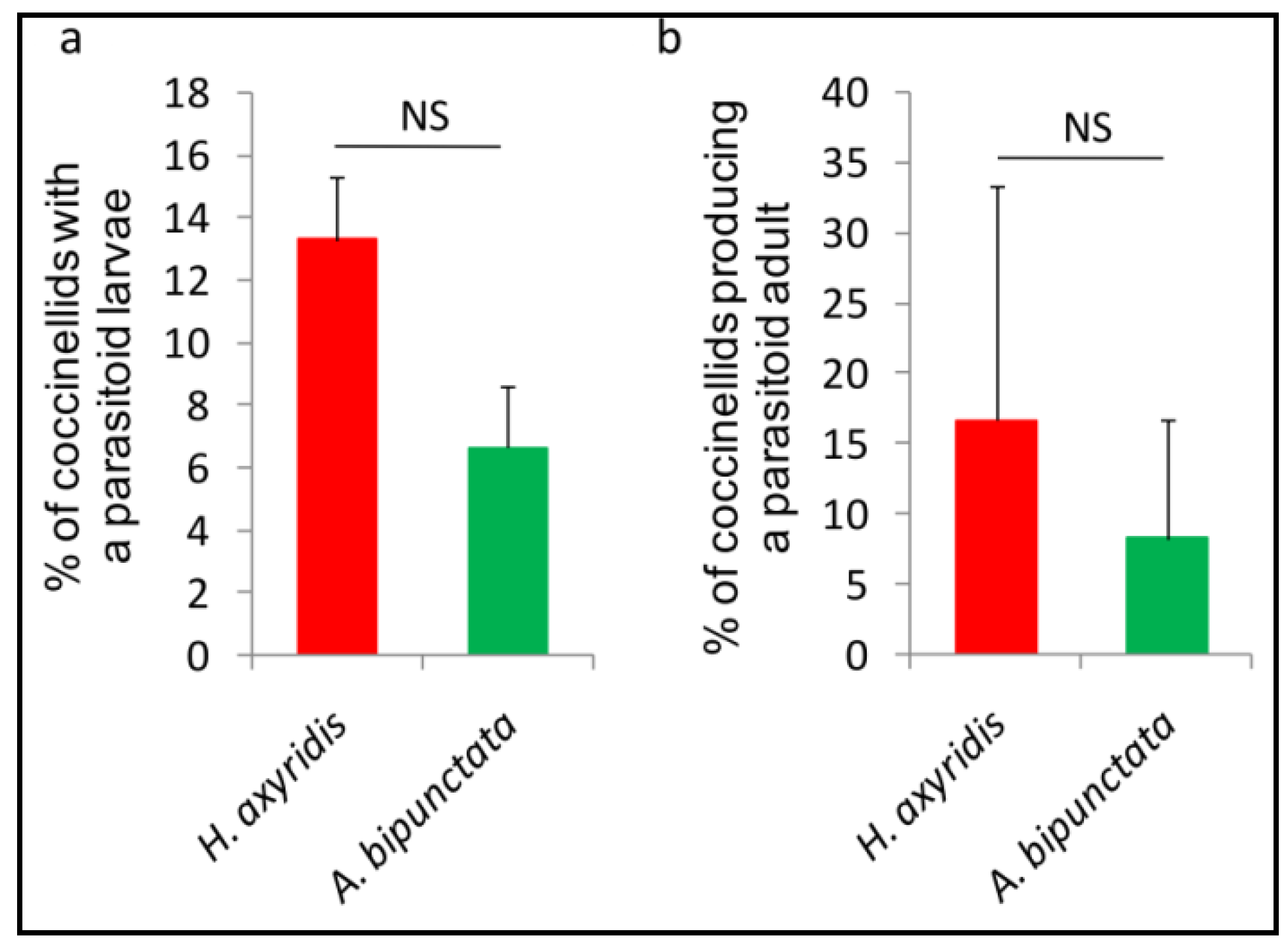
3.4. Effects of D. Coccinellae on H. Axyridis vs. A. Bipunctata
3.4.1. Acceptance and Suitability
3.4.2. Effects on Female Longevity, Oviposition and Egg Fertility
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koch, R.L. The multicolored Asian lady beetle, Harmonia axyridis: A review of its biology, uses in biological control, and non-target impacts. J. Insect Sci. 2003, 3, 16–32. [Google Scholar] [CrossRef]
- Brown, P.M.; Thomas, C.E.; Lombaert, E.; Jeffries, D.L.; Estoup, A.; Handley, L.-J.L. The global spread of Harmonia axyridis (Coleoptera: Coccinellidae): Distribution, dispersal and routes of invasion. BioControl 2011, 56, 623–641. [Google Scholar]
- Roy, H.E.; Brown, P.M.J.; Adriaens, T.; Berkvens, N.; Borges, I.; Clusella-Trullas, S.; Comont, R.F.; de Clercq, P.; Eschen, R.; Estoup, A.; et al. The harlequin ladybird, Harmonia axyridis: Global perspectives on invasion history and ecology. Biol. Invasions 2016, 18, 997–1044. [Google Scholar] [CrossRef]
- Hodek, I.; Evans, E.W. Food relationships. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honěk, A., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2012; pp. 142–243. [Google Scholar]
- Nedvěd, O.; Honěk, A. Life history and development. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honěk, A., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2012; pp. 55–97. [Google Scholar]
- Burgio, G.; Santi, F.; Lanzoni, A.; Masetti, A.; de Luigi, V.; Melandri, M.; Reggiani, A.; Ricci, C.; Loomans, A.J.M.; Maini, S. Harmonia axyridis recordings in northern Italy. Bull. Insectol. 2008, 61, 361–364. [Google Scholar]
- Burgio, G.; Santi, F.; Maini, S. On intra-guild predation and cannibalism in Harmonia axyridis (Pallas) and Adalia bipunctata L. (Coleoptera: Coccinellidae). Biol. Control 2002, 24, 110–116. [Google Scholar] [CrossRef]
- Roy, H.E.; De Clercq, P.; Handley, L.-J.; Sloggett, J.J.; Poland, R.L.; Wajnberg, E. Special issue: Invasive alien arthropod predators and parasitoids: An ecological approach. BioControl 2011, 56, 375–382. [Google Scholar]
- Glemser, E.J.; Dowling, L.; Inglis, D.; Pickering, G.J.; McFadden-Smith, W.; Sears, M.K.; Hallett, R.H. A novel method for controlling multicolored Asian lady beetle (Coleoptera: Coccinellidae) in vineyards. Environ. Entomol. 2012, 41, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Disney, R.H. A new species of Phoridae (Diptera) that parasitises a wide-spread Asian ladybird beetle (Coleoptera: Coccinellidae). Entomologist 1997, 116, 163–168. [Google Scholar]
- Park, H.; Park, Y.C.; Hong, O.K.; Cho, S.Y. Parasitoids of the aphidophagous ladybeetles, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae) in Chuncheon areas, Korea. Korean J. Entomol. 1996, 26, 143–147. [Google Scholar]
- Katsoyannos, P.; Aliniazee, M.T. First record of Strongygaster triangulifera (Loew) (Diptera: Tachinidae) as a parasitoid of Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae) in western North America. Can. Entomol. 1998, 130, 905–906. [Google Scholar] [CrossRef]
- Nelepa, C.A.; Kidd, K.A. Parasitism of the multicolored Asian lady beetle (Coleoptera: Coccinellidae) by Strongygaster triangulifera (Diptera: Tachinidae) in North Carolina. J. Entomol. Sci. 2002, 37, 124–127. [Google Scholar]
- Ware, R.; Michie, L.-J.; Otani, T.; Rhule, E.; Hall, R. Adaptation of native parasitoids to a novel host: The invasive coccinellid Harmonia axyridis. IOBC/WPRS Bull. 2010, 58, 175–182. [Google Scholar]
- Francati, S. Native preimaginal parasitism of Harmonia axyridis: New record of association with Phalacotrophora fasciata in Italy. Bull. Insectol. 2015, 68, 3–6. [Google Scholar]
- Ceryngier, P.; Roy, H.E.; Poland, R.L. Natural enemies of ladybird beetles. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honěk, A., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2012; pp. 375–444. [Google Scholar]
- Maeta, Y. Some biological studies on the natural enemies of some coccinellid beetles. Tohoku Konchu Kenyu 1969, 4, 1–6. [Google Scholar]
- Firlej, A.; Lucas, E.; Coderre, D.; Boivin, G. Impact of host behavioral defenses on parasitization efficacy of a larval and adult parasitoid. BioControl 2010, 55, 339–348. [Google Scholar]
- Triltsch, H. On the parasitization of the ladybird Coccinella septempunctata L. (Col, Coccinellidae). J. Appl. Entomol. 1996, 120, 375–378. [Google Scholar] [CrossRef]
- Berkvens, N.; Moens, J.; Berkvens, D.; Samih, M.A.; Tirry, L.; DeClercq, P. Dinocampus coccinellae as a parasitoid of the invasive ladybird Harmonia axyridis in Europe. Biol. Control 2010, 53, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Maure, F.; Doyon, J.; Thomas, F.; Brodeur, J. Host behaviour manipulation as an evolutionary route towards attenuation of parasitoid virulence. J. Evolution. Biol. 2014, 27, 2871–2875. [Google Scholar] [CrossRef] [PubMed]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Burgio, G.; Lanzoni, A.; Accinelli, G.; Maini, S. Estimation of mortality by entomophages on exotic Harmonia axyridis versus Adalia bipunctata in semi-field conditions in northern Italy. BioControl 2008, 53, 277–287. [Google Scholar] [CrossRef]
- Majerus, M.E.N.; Kearns, P. Ladybirds (Naturalists’ Handbook Series 10); Richmond Publishing: Slough, UK, 1989. [Google Scholar]
- Lanzoni, A.; Accinelli, G.; Bazzocchi, G.G.; Burgio, G. Biological traits and life table of the exotic Harmonia axyridis compared with Hippodamia variegata and Adalia bipunctata (Col., Coccinellidae). J. Appl. Entomol. 2004, 128, 298–306. [Google Scholar] [CrossRef]
- Goidanich, A. Materiali per lo studio degli imenotteri braconidi. Boll. Lab. Entomol. R. Ist. Super. Agric. Bol. 1933, 6, 33–50. [Google Scholar]
- McCornack, B.P.; Koch, R.L.; Ragsdale, D.W. A simple method for in-field sex determination of the multicolored Asian lady beetle Harmonia axyridis. J. Insect Sci. 2007, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sighinolfi, L.; Febvay, G.; Dindo, M.L.; Rey, M.; Pageaux, J.-F.; Baronio, P.; Grenier, S. Biological and biochemical characteristics for quality control of Harmonia axyridis (Pallas) (Coleoptera, Coccinellidae) reared on a liver-based diet. Arch. Insect Biochem. 2008, 68, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.W.; Stevenson, A.T.; Richards, D.R. Essential versus alternative foods of insect predators: Benefits of a mixed diet. Oecologia 1999, 121, 107–112. [Google Scholar] [CrossRef]
- Sighinolfi, L.; Febvay, G.; Dindo, M.L.; Rey, M.; Pageaux, J.F.; Grenier, S. Biochemical content in fatty acids and biological parameters of Harmonia axyridis reared on artificial diet. Bull. Insectol. 2013, 66, 283–290. [Google Scholar]
- Obrycki, J.J.; Tauber, M.J.; Tauber, C.A. Perilitus coccinellae (Hymenoptera: Braconidae): Parasitization and development in relation to host-stage attacked. Ann. Entomol. Soc. Am. 1985, 78, 852–854. [Google Scholar] [CrossRef]
- Kadono-Okuda, K.; Sakurai, H.; Takeda, S.; Okuda, T. Synchronous growth of parasitoid, Perilitus coccinellae, and teratocytes with the development of the host, Coccinella septempunctata. Entomol. Exp. Appl. 1995, 75, 145–149. [Google Scholar] [CrossRef]
- StatSoft, Inc. STATISTICA (Data Analysis Software System), Version 10. 2011. Available online: www.statsoft.com (accessed on 19 October 2016).
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Englewood Cliffs, NJ, USA, 1984. [Google Scholar]
- Kenis, M.; Roy, H.E.; Majerus, M.E.N. Current and potential management strategies against Harmonia axyridis. BioControl 2008, 53, 235–252. [Google Scholar] [CrossRef] [Green Version]
- Hoogendoorn, M.; Heimpel, G.E. Indirect interactions between an introduced and a native ladybird beetle species mediated by a shared parasitoid. Biol. Control 2002, 25, 224–230. [Google Scholar] [CrossRef]
- Firlej, A.; Boivin, G.; Lucas, E.; Coderre, D. First report of Harmonia axyridis Pallas being attacked by Dinocampus coccinellae Schrank in Canada. Biol. Invasions 2005, 7, 553–556. [Google Scholar] [CrossRef]
- Browdrey, J.; Mabbott, P. The multivariate Asian ladybird Harmonia axyridis Pallas in Essex. Essex Nat. 2004, 22, 42–44. [Google Scholar]
- Koyama, S.; Majerus, M.E.N. Interactions between the parasitoid wasp Dinocampus coccinellae and two species of coccinellid from Japan and Britain. In From Biological Control to Invasion: The Ladybird Harmonia axyridis as a Model Species; Roy, H.E., Wajnberg, E., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 253–264. [Google Scholar]
- Richerson, J.V.; Deloach, C.J. Some aspects of host selection by Perilitus coccinellae (Hymenoptera Braconidae). Ann. Entomol. Soc. Am. 1972, 65, 834–839. [Google Scholar] [CrossRef]
- Firlej, A.; Lucas, E.; Coderre, D.; Boivin, G. Teratocytes growth pattern reflects host suitability in a host-parasitoid assemblage. Physiol. Entomol. 2007, 32, 181–187. [Google Scholar] [CrossRef]
- Burling, J.M.; Remy, L.W.; Handley, L.L. Is Harmonia axyridis a marginal host for the parasitoid wasp Dinocampus coccinellae? Morphological and molecular genetic analyses. IOBC/WPRS Bull. 2010, 58, 23–25. [Google Scholar]
- De Castro-Guedes, C.; de Almeida, L. Laboratory investigations reveal that Harmonia axyridis (Coleoptera: Coccinellidae) is a poor host for Dinocampus coccinellae (Hymenoptera: Braconidae) in Brazil. J. Insect Sci. 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Ceryngier, P. Host discrimination in Dinocampus coccinellae (Hymenoptera: Braconidae), a solitary parasitoid of coccinellid beetles. Appl. Entomol. Zool. 2000, 35, 535–539. [Google Scholar] [CrossRef]
- Balduf, W.V. The bionomics of Dinocampus coccinellae Schrank. Ann. Entomol. Soc. Am. 1926, 19, 465–498. [Google Scholar] [CrossRef]
- Sluss, R. Behavioral and anatomical responses of the convergent lady beetle to parasitism by Perilitus coccinellae (Schrank) (Hymenoptera: Braconidae). J. Invertebr. Pathol. 1968, 10, 9–27. [Google Scholar] [CrossRef]
- Harvey, J.A.; Harvey, I.F.; Thompson, D.J. The effect of superparasitism on development of the solitary parasitoid wasp, Venturia canescens (Hymenoptera: Ichneumonidae). Ecol. Entomol. 1993, 18, 203–208. [Google Scholar] [CrossRef]
- Tunca, H.; Klncer, N. Effect of superparasitism on the development of the solitary parasitoid Chelonus oculator Panzer (Hymenoptera: Braconidae). Turk. J. Agric. For. 2009, 33, 463–468. [Google Scholar]
- Hodek, I.; Honěk, A. Ecology of Coccinellidae; Kluwer Academic Publishers: Boston, MA, USA, 1996. [Google Scholar]
- Riddick, E.W.; Cottrell, T.E.; Kidd, K.A. Natural enemies of the Coccinellidae: Parasites, pathogens, and parasitoids. Biol. Control 2009, 51, 306–312. [Google Scholar] [CrossRef]
- Riddick, E.W.; Cottrell, T.E. Is the prevalence and intensity of the ectoparasitic fungus Hesperomyces virescens related to the abundance of entomophagous coccinellids? Bull. Insectol. 2010, 63, 71–78. [Google Scholar]
- Rondoni, G.; Athey, K.J.; Hardwood, J.D.; Conti, E.; Ricci, C.; Obrycki, J.J. Development and application of molecular gut-content analysis to detect aphid and coccinellid predation by Harmonia axyridis (Coleoptera: Coccinellidae) in Italy. Insect Sci. 2015, 22, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.W.; Miller, S.S. Coccinellidae (Coleoptera) in apple orchards of eastern West Virginia and the impact of invasion by Harmonia axyridis. ENT News 1998, 109, 136–142. [Google Scholar]
- Honěk, A.; Martinkova, Z.; Kindlmann, P.; Ameixa, O.M.C.C.; Dixon, A.F.G. Long-term trends in the composition of aphidophagous coccinellid communities in Central Europe. Insect Conserv. Divers. 2013. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Exposure Time | Parasitoid Cocoons Detected (1) | χ2 (3) | p | Parasitoid Adults Emerged (2) | χ2 (3) | p |
|---|---|---|---|---|---|---|
| 5 min | 18 ± 0.04 | 8.22 | 0.004 | 89 ± 0.08 | 0.21 | 0.647 |
| 1 h | 36 ± 0.05 | 89 ± 0.05 | ||||
| 5 min | 18 ± 0.04 | 3.37 | 0.067 | 89 ± 0.08 | 0.20 | 0.653 |
| 24 h | 29 ± 0.05 | 79 ± 0.08 | ||||
| 1 h | 36 ± 0.05 | 1.12 | 0.291 | 89 ± 0.05 | 0.52 | 0.473 |
| 24 h | 29 ± 0.05 | 79 ± 0.08 |
| Exposure Time | Time from Exposure to Cocoon | Time from Cocoon to Adult | Time from Exposure to Adult |
|---|---|---|---|
| 5 min | 17.44 ± 1.34 a | 7.44 ± 0.96 | 24.68 ± 1.20 a |
| 1 h | 17.08 ± 0.77 a | 7.25 ± 0.76 | 24.38 ± 0.87 a |
| 24 h | 19.41 ± 3.21 b | 7.48 ± 1.2 | 26.70 ± 3.59 b |
| H (N) = 23.66 (83) | F (df) = 0.43 (2,64) | H (N) = 11.34 (71) | |
| p | <0.001 | 0.65 | 0.035 |
| Male Parasitization Status | Pre-Oviposition (Days) | Oviposition (Days) | Eggs Laid in 10 Days (E10) | Fertility (% Hatched Eggs) |
|---|---|---|---|---|
| Not exposed (controls) (n = 20) | 5.25 ± 0.33 | 20.4 ± 1 | 306.65 ± 23.32 | 62.79 ± 3.99 a |
| Successfully exposed (n = 7) | 5.86 ± 0.56 | 19 ± 1.69 | 310.57 ± 39.31 | 48.61 ± 6.75 a,b |
| Unsuccessfully exposed (n = 13) | 4.85 ± 0.41 | 20.77 ± 1.24 | 266.54 ± 28.92 | 35.06 ± 4.95 b |
| F (df = 2,37) | 1.05 | 0.37 | 0.68 | 9.26 |
| p | 0.36 | 0.69 | 0.51 | <0.001 |
| Female Parasitization Status | Pre-Oviposition (Days) | Oviposition (Days) | Eggs Laid in 10 Days (E10) | Fertility (% Hatched Eggs) |
|---|---|---|---|---|
| Not exposed (control) (n = 20) | 7.7 ± 0.47 | 16.3 ± 0.49 a | 359.7 ± 21.22 a | 49.92 ± 3.09 a |
| Successfully exposed (n = 3) | 8 ± 1.53 | 8 ± 3.61 b | 31 ± 13.08 b | 6.06 ± 6.06 b |
| Unsuccessfully exposed (n = 17) | 6.59 ± 0.74 | 11.82 ± 1.51 b | 175.53 ± 28.79 b | 21.28 ± 4.69 b |
| F (2,37) = 0.98 | H (40) = 10.76 | F (2,37) = 2.19 | H (40) = 0.92 | |
| p | 0.38 | 0.005 | <0.001 | <0.001 |
| Parameters | Coccinellid Species | Exposure to D. coccinellae | Source of Variation | |||
|---|---|---|---|---|---|---|
| Yes | No | Species Effect | Exposure Effect | Interaction Species × Exposure | ||
| Longevity (days) | H. axyridis | (16) 20.1 ± 0.9 | (20) 22.2 ± 0.9 | |||
| F = 0.02 | F = 2.97 | F = 0.23 | ||||
| df = 1,72 | df = 1,72 | df = 1,72 | ||||
| p = 0.9 | p = 0.09 | p = 0.63 | ||||
| A. bipunctata | (20) 20.7 ± 1.2 | (20) 21.9 ± 0.8 | ||||
| Pre-oviposition (days) | H. axyridis | (14) 5.93 ± 0.97 | (19) 5.11 ± 0.4 | |||
| F = 8.99 | F = 1.3 | F = 0.34 | ||||
| df = 1,67 | df = 1,67 | df = 1,67 | ||||
| p = 0.004 | p = 0.26 | p = 0.56 | ||||
| A. bipunctata | (19) 4.21 ± 0.3 | (19) 3.95 ± 0.3 | ||||
| Oviposition (days) | H. axyridis | (14) 10.1 ± 1.7 | (19) 16.6 ± 0.72 | |||
| F = 3.36 | F = 14.4 | F = 0.36 | ||||
| df = 1,67 | df = 1,67 | df = 1,67 | ||||
| p = 0.07 | p = 0.0003 | p = 0.56 | ||||
| A. bipunctata | (19) 12.9 ± 1.5 | (19) 17.1 ± 0.9 | ||||
| Fertility (% hatched eggs) | H. axyridis | (14) 22 ± 5.4 | (19) 35.4 ± 4.1 | |||
| F = 7.36 | F = 6.75 | F = 0.39 | ||||
| df = 1,67 | df = 1,67 | df = 1,67 | ||||
| p = 0.008 | p = 0.012 | p = 0.54 | ||||
| A. bipunctata | (19) 13.7 ± 3.3 | (19) 20.6 ± 3.7 | ||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dindo, M.L.; Francati, S.; Lanzoni, A.; Di Vitantonio, C.; Marchetti, E.; Burgio, G.; Maini, S. Interactions between the Multicolored Asian Lady Beetle Harmonia axyridis and the Parasitoid Dinocampus coccinellae. Insects 2016, 7, 67. https://doi.org/10.3390/insects7040067
Dindo ML, Francati S, Lanzoni A, Di Vitantonio C, Marchetti E, Burgio G, Maini S. Interactions between the Multicolored Asian Lady Beetle Harmonia axyridis and the Parasitoid Dinocampus coccinellae. Insects. 2016; 7(4):67. https://doi.org/10.3390/insects7040067
Chicago/Turabian StyleDindo, Maria Luisa, Santolo Francati, Alberto Lanzoni, Cinzia Di Vitantonio, Elisa Marchetti, Giovanni Burgio, and Stefano Maini. 2016. "Interactions between the Multicolored Asian Lady Beetle Harmonia axyridis and the Parasitoid Dinocampus coccinellae" Insects 7, no. 4: 67. https://doi.org/10.3390/insects7040067
APA StyleDindo, M. L., Francati, S., Lanzoni, A., Di Vitantonio, C., Marchetti, E., Burgio, G., & Maini, S. (2016). Interactions between the Multicolored Asian Lady Beetle Harmonia axyridis and the Parasitoid Dinocampus coccinellae. Insects, 7(4), 67. https://doi.org/10.3390/insects7040067