Elevated CO2 Concentrations Impact the Semiochemistry of Aphid Honeydew without Having a Cascade Effect on an Aphid Predator




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plants and Insects
2.2. Honeydew Collection
2.3. Quantification of Honeydew
2.4. Identification and Estimated Quantity of Honeydew Semiochemicals
2.5. Behavioral Experiment
2.6. Statistical Analyses
3. Results
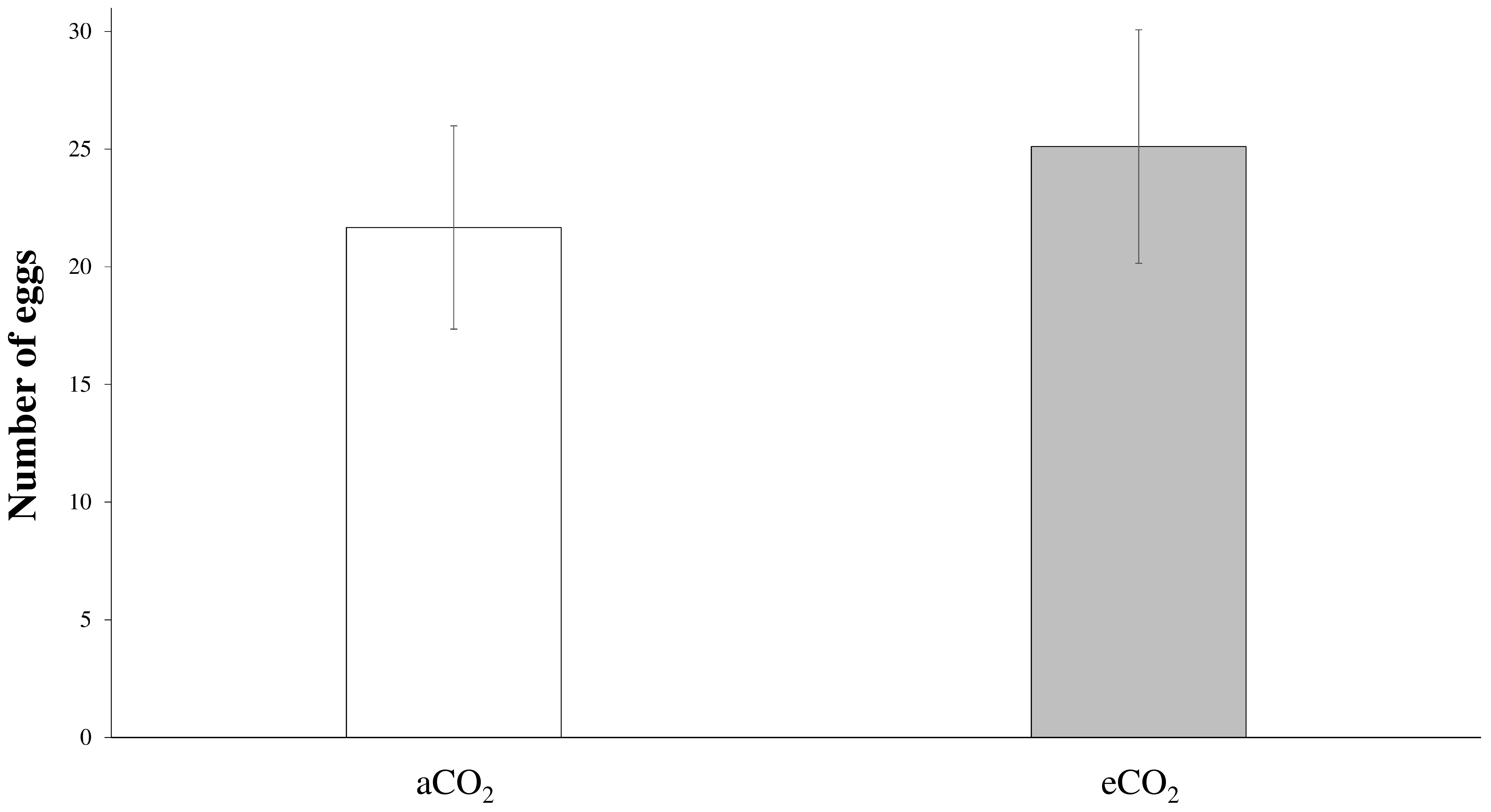
3.1. Quantification of Honeydew
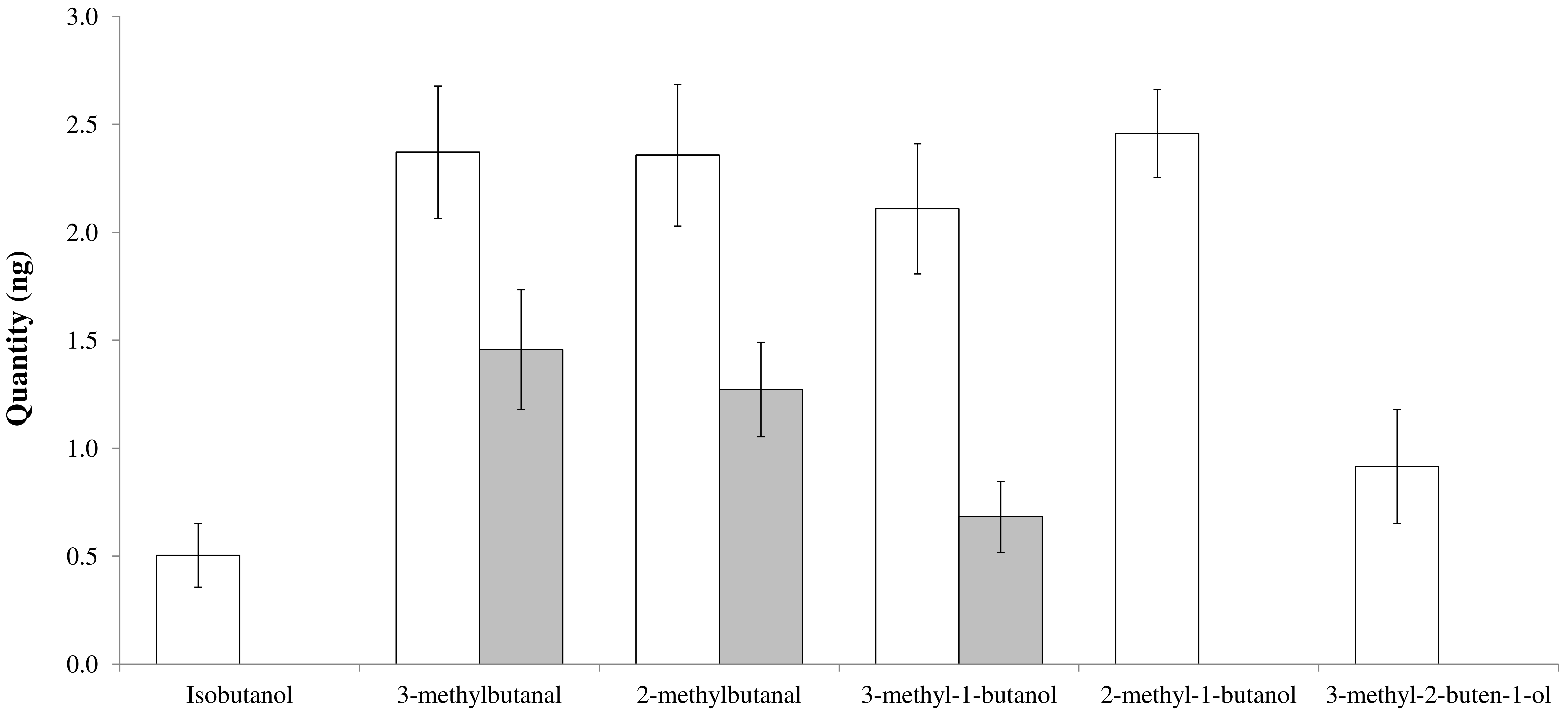
3.2. Honeydew Semiochemical Analysis
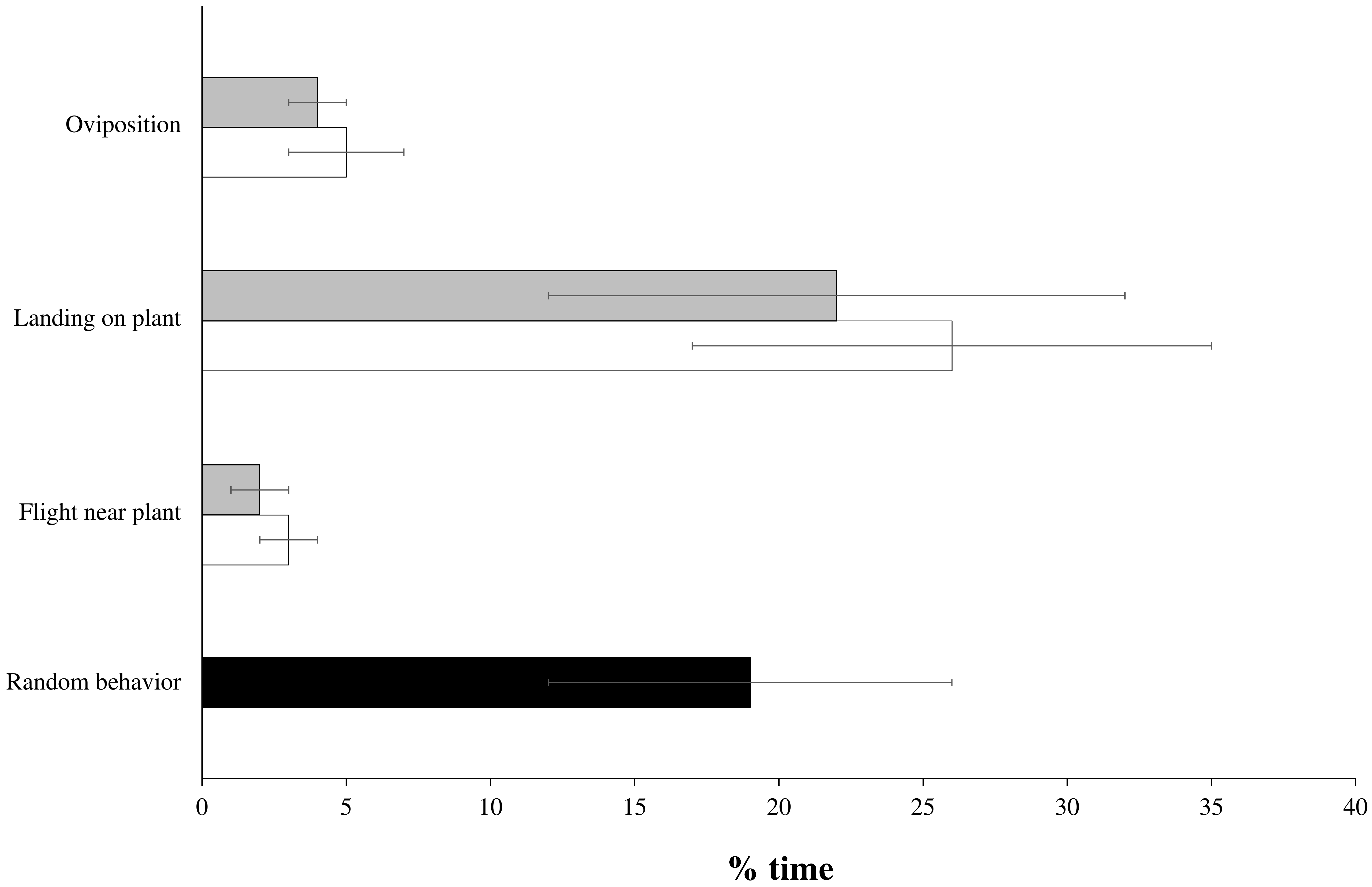
3.3. Behavioral Experiments
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hoffmann, K.H. Aphid honeydew: Rubbish or signaler. In Biology and Ecology of Aphids; Vilcinskas, A., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 199–220. [Google Scholar]
- Leroy, P.D.; Heuskin, S.; Sabri, A.; Verheggen, F.J.; Farmakidis, J.; Lognay, G.; Thonard, P.; Wathelet, J.P.; Brosteaux, Y.; Haubruge, E. Honeydew volatile emission acts as a kairomonal message for the Asian lady beetle Harmonia axyridis (Coleoptera: Coccinellidae). Insect Sci. 2012, 19, 498–506. [Google Scholar] [CrossRef]
- Budenberg, W.J. Honeydew as a contact kairomone for aphid parasitoids. Entomol. Exp. Appl. 1990, 55, 139–148. [Google Scholar] [CrossRef]
- Fischer, M.; Völkl, W.; Hoffmann, K.H. Honeydew production and honeydew sugar composition of polyphagous black bean aphid, Aphis fabae (Hemiptera: Aphididae) on various host plants and implications for ant-attendance. Eur. J. Entomol. 2005, 102, 155–160. [Google Scholar] [CrossRef]
- Auclair, J. Aphid feeding and nutrition. Annu. Rev. Entomol. 1963, 8, 439–490. [Google Scholar] [CrossRef]
- Völkl, W.; Woodring, J.; Fischer, M.; Lorenz, M.; Hoffmann, K.H. Ant-aphid mutualisms: The impact of honeydew production and honeydew sugar composition on ant preferences. Oecologia 1999, 118, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Leroy, P.D.; Wathelet, B.; Sabri, A.; Francis, F.; Verheggen, F.J.; Capella, Q.; Thonard, P.; Haubruge, E. Aphid-host plant interactions: Does aphid honeydew exactly reflect the host plant amino acid composition? Arthropod-Plant Interact. 2011, 5, 193–199. [Google Scholar] [CrossRef]
- Wilkinson, T.; Ashford, D.; Pritchard, J.; Douglas, A. Honeydew sugars and osmoregulation in the pea aphid Acyrthosiphon pisum. J. Exp. Biol. 1997, 200, 2137–2143. [Google Scholar] [PubMed]
- Leroy, P.D.; Sabri, A.; Heuskin, S.; Thonart, P.; Lognay, G.; Verheggen, F.J.; Francis, F.; Brosteaux, Y.; Felton, G.W.; Haubruge, E. Microorganisms from aphid honeydew attract and enhance the efficacy of natural enemies. Nat. Commun. 2011, 2, 348. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Lognay, G.; Detrain, C.; Heil, M.; Grigorescu, A.; Sabri, A.; Thonard, P.; Haubruge, E.; Verheggen, F.J. Bacteria may enhance species association in an ant-aphid mutualistic relationship. Chemoecology 2015, 25, 223–232. [Google Scholar] [CrossRef]
- Scholz, D.; Poehling, H.M. Oviposition site selection of Episyrphus balteatus. Entomol. Exp. Appl. 2000, 94, 149–158. [Google Scholar] [CrossRef]
- Robinson, E.; Ryan, G.; Newman, J. A meta-analytical review of the effects of elevated CO2 on plant—Arthropod interactions highlights the importance of interacting environmental and biological variables. New Phytol. 2012, 194, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Guiney, D. Regulation of bacterial virulence gene expression by the host environment. J. Clin. Investig. 1997, 99, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Boullis, A.; Francis, F.; Verheggen, F.J. Climate change and tritrophic interactions: Will modifications to greenhouse gas emissions increase the vulnerability of herbivorous insects to natural enemies? Environ. Entomol. 2015, 44, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Boullis, A.; Fassotte, B.; Sarles, L.; Lognay, G.; Heuskin, S.; Vanderplanck, M.; Bartram, S.; Haubruge, E.; Francis, F.; Verheggen, F.J. Elevated carbon dioxide concentration reduces alarm signaling in aphids. J. Chem. Ecol. 2017, 43, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Leroy, P.D.; Almohamad, R.; Attia, S.; Capella, Q.; Verheggen, F.J.; Haubruge, E.; Francis, F. Aphid honeydew: An arrestant and a contact kairomone for Episyrphus balteatus (Diptera: Syrphidae) larvae and adults. Eur. J. Entomol. 2014, 111, 237–242. [Google Scholar] [CrossRef]
- Vosteen, I.; Gershenzon, J.; Kunert, G. Hoverfly preference for high honeydew amounts creates enemy-free space for aphids colonizing novel host plants. J. Anim. Ecol. 2016, 85, 1286–1297. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 1 May 2013).
- Schulz, S.; Dickschat, J.S. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef] [PubMed]
- Bidart-Bouzat, M.; Imeh-Nathaniel, A. Global change effects on plant chemical defenses against insect herbivores. J. Integr. Plant Biol. 2008, 50, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.N.; Ryalls, J.; Karley, A. Global climate change and crop resistance to aphids: Contrasting responses of lucerne genotypes to elevated atmospheric carbon dioxide. Ann. Appl. Biol. 2014, 165, 62–72. [Google Scholar] [CrossRef]
- Sun, Y.; Jing, B.; Ge, F. Response of amino acid changes in Aphis gossypii (Glover) to elevated CO2 levels. J. Appl. Entomol. 2009, 133, 189–197. [Google Scholar] [CrossRef]
- Wang, N.; Nobel, P. Phloem exudate collected via scale insect stylets for the CAM species Opuntia-ficus-indica under current and doubled CO2 concentrations. Ann. Bot. 1995, 75, 525–532. [Google Scholar] [CrossRef]
- Ryalls, J.; Moore, B.; Riegler, M.; Bromfield, L.; Hall, A.; Johnson, S.N. Climate and atmospheric change impacts on sap-feeding herbivores: A mechanistic explanation based on functional groups of primary metabolites. Funct. Ecol. 2017, 31, 161–171. [Google Scholar] [CrossRef]
- Oehme, V.; Högy, P.; Zebitz, C.; Fangmeier, A. Effects of elevated atmospheric CO2 concentrations on phloem sap composition of spring crops and aphid performance. J. Plant Interact. 2013, 8, 74–84. [Google Scholar] [CrossRef]
- Ryan, G.; Sylvester, E.; Shelp, B.; Newman, J. Towards an understanding of how phloem amino acid composition shapes elevated CO2-induced changes in aphid population dynamics. Ecol. Entomol. 2015, 40, 247–257. [Google Scholar] [CrossRef]
- Budenberg, W.J.; Powell, W. The role of honeydew as an ovipositional stimulant for two species of syrphids. Entomol. Exp. Appl. 1992, 64, 57–61. [Google Scholar] [CrossRef]
- Fu, X.; Ye, L.; Kang, L.; Ge, F. Elevated CO2 shifts the focus of tobacco plant defences from cucumber mosaic virus to the green peach aphid. Plant Cell Environ. 2010, 33, 2056–2064. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | CAS # | Retention Time (min) | aCO2 | eCO2 | P |
|---|---|---|---|---|---|
| Acetaldehyde | 75-07-0 | 3.406 | D | - | - |
| Ethanol | 64-17-5 | 3.592 | D | D | - |
| 2-propanone | 407-25-0 | 3.756 | D | D | - |
| 2-thiapropane | 75-18-3 | 3.939 | D | D | - |
| Isobutanol | 78-83-1 | 4.958 | 0.504 ± 0.147 | - | - |
| 3-methylbutanal | 590-86-3 | 5.391 | 2.371 ± 0.306 | 1.456 ± 0.277 | 0.197 |
| 2-methylbutanal | 96-17-3 | 5.557 | 2.357 ± 0.328 | 1.272 ± 0.219 | 0.081 |
| 3-methyl-1-butanol | 123-51-3 | 7.038 | 2.108 ± 0.301 | 0.682 ± 0.164 | 0.009 |
| 2-methyl-1-butanol | 137-32-6 | 7.077 | 2.458 ± 0.203 | - | - |
| 3-methyl-2-buten-1-ol | 556-82-1 | 8.201 | 0.916 ± 0.264 | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boullis, A.; Blanchard, S.; Francis, F.; Verheggen, F. Elevated CO2 Concentrations Impact the Semiochemistry of Aphid Honeydew without Having a Cascade Effect on an Aphid Predator. Insects 2018, 9, 47. https://doi.org/10.3390/insects9020047
Boullis A, Blanchard S, Francis F, Verheggen F. Elevated CO2 Concentrations Impact the Semiochemistry of Aphid Honeydew without Having a Cascade Effect on an Aphid Predator. Insects. 2018; 9(2):47. https://doi.org/10.3390/insects9020047
Chicago/Turabian StyleBoullis, Antoine, Solène Blanchard, Frédéric Francis, and François Verheggen. 2018. "Elevated CO2 Concentrations Impact the Semiochemistry of Aphid Honeydew without Having a Cascade Effect on an Aphid Predator" Insects 9, no. 2: 47. https://doi.org/10.3390/insects9020047
APA StyleBoullis, A., Blanchard, S., Francis, F., & Verheggen, F. (2018). Elevated CO2 Concentrations Impact the Semiochemistry of Aphid Honeydew without Having a Cascade Effect on an Aphid Predator. Insects, 9(2), 47. https://doi.org/10.3390/insects9020047