Pheromone-Trap Monitoring System for Pea Leaf Weevil, Sitona lineatus: Effects of Trap Type, Lure Type and Trap Placement within Fields


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sites
2.2. Pheromone Lures
2.3. Experiment #1: Trap Type
2.4. Experiment #2: Lure Type
2.5. Experiment #3: Trap Placement
2.6. Statistical Analysis
3. Results
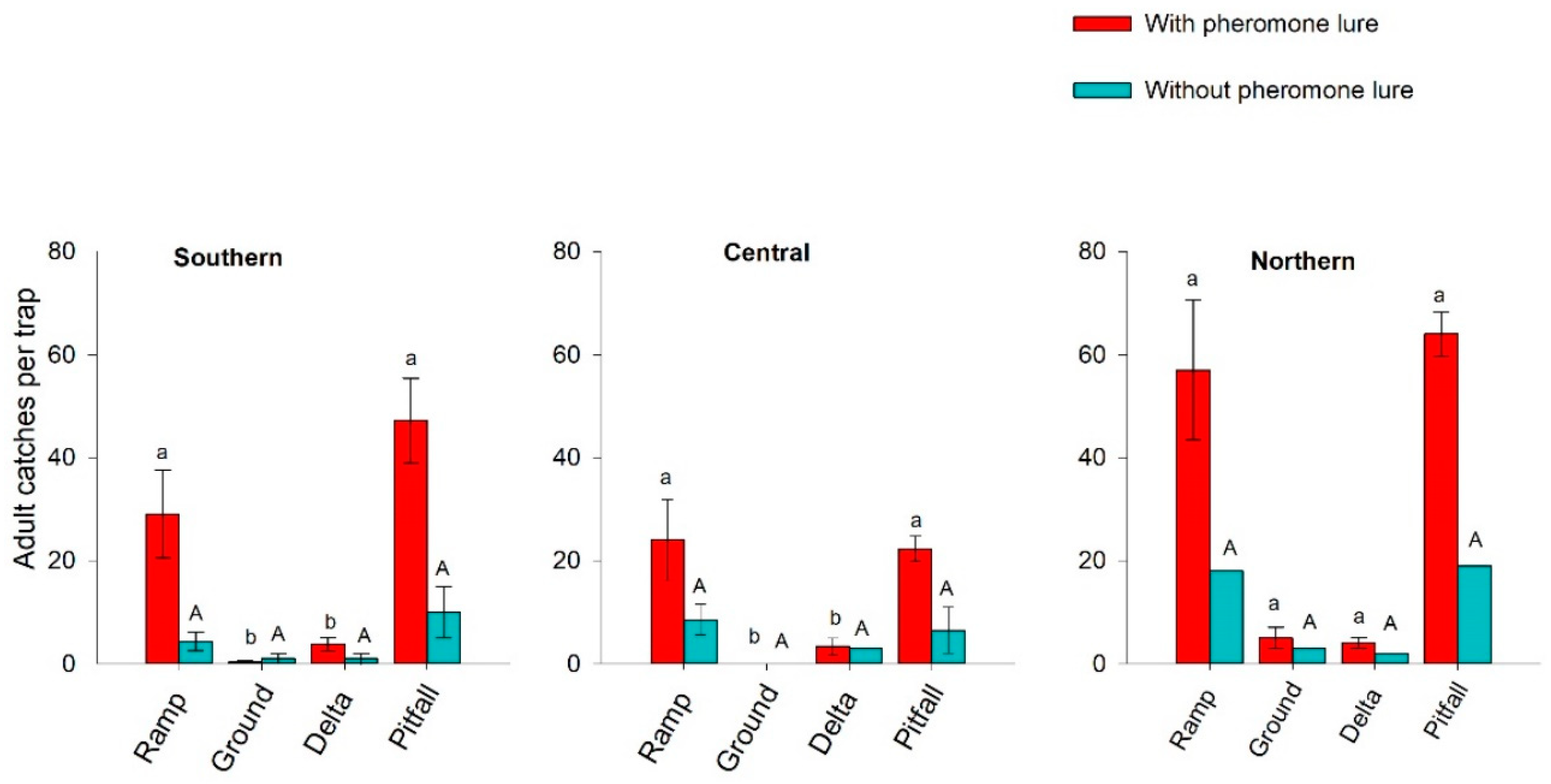
3.1. Experiment #1 Trap Type
3.2. Experiment #2: Lure Type
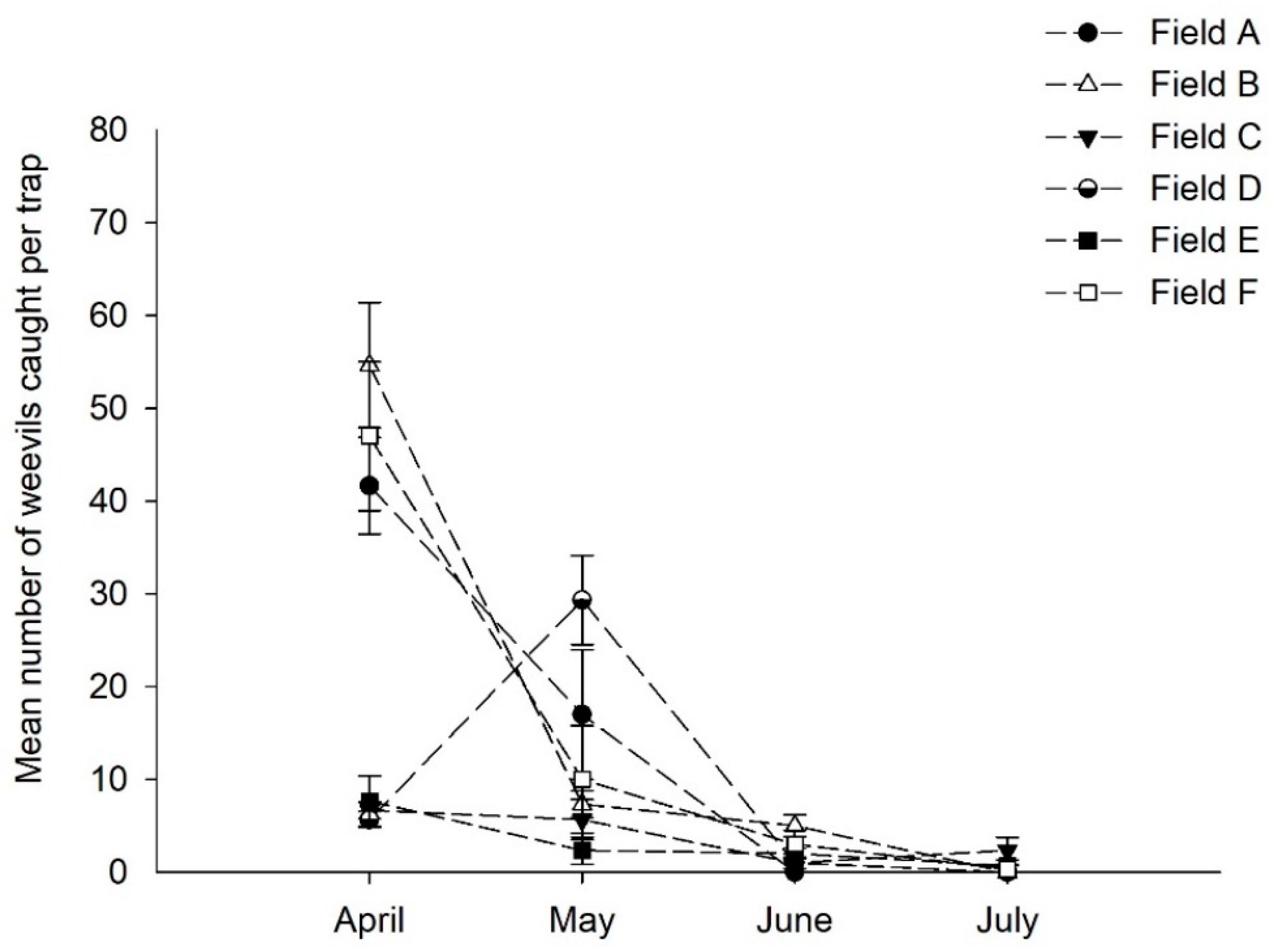
3.3. Experiment #3: Trap Placement
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- USDA NASS. U.S. Crop Production National Agricultural Statistic Service. Available online: http://usda.mannlib.cornell.edu/MannUsda/viewDocumentInfo.do;jsessionid=9F3E2707 A44E1E371AA16E777C7F8362?documentID=1046 (accessed on 21 December 2017).
- Montana Agricultural Statistics; US Department of Agriculture. National Agricultural Statistics Services. Volume LIV. Available online: https://wwwnassusdagov/Statistics (accessed on 7 March 2018).
- Foyer, C.H.; Lam, H.-M.; Nguyen, H.T.; Siddique, K.H.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M. Neglecting legumes has compromised human health and sustainable food production. Nat. Plants 2016, 2, 16112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, B.S. Yield responses of Vicia faba in relation to infestation levels of Sitona lineatus L. (Col.: Curculionidae). J. Appl. Entomol. 1990, 110, 398–407. [Google Scholar] [CrossRef]
- Hoebeke, E.R.; Wheeler, A.G.J. Sitona lineatus (L.), the pea leaf weevil: First records in eastern North America (Coleoptera: Curculionidae). Proc. Entomol. Soc. Wash. 1985, 87, 216–220. [Google Scholar]
- Bloem, S.; Mizell, R.F.; O’Brien, C.W. Old traps for new weevils: New records for curculionids (Coleoptera: Curculionidae), brentids (Coleoptera: Brentidae) and anthribids (Coleoptera: Anthribidae) from Jefferson Co., Florida. Fla. Entomol. 2002, 85, 632–644. [Google Scholar] [CrossRef]
- Vankosky, M.; Dosdall, L.; Cárcamo, H. Distribution, biology and integrated management of the pea leaf weevil, Sitona lineatus L. (Coleoptera: Curculionidae), with an analysis of research needs. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Res. 2009, 4, 1–18. [Google Scholar] [CrossRef]
- Downes, W. The occurrence of Sitona lineatus L. in British Columbia. Can. Entomol. 1938, 70, 320–322. [Google Scholar] [CrossRef]
- Wanner, K.W. Pea Leaf Weevil. MontGuide; Montana State University: Bozeman, MT, USA, 2016; pp. 1–2. [Google Scholar]
- Vankosky, M.; Cárcamo, H.; Dosdall, L. Identification of potential natural enemies of the pea leaf weevil, Sitona lineatus L. in western Canada. J. Appl. Entomol. 2011, 135, 293–301. [Google Scholar] [CrossRef]
- Hamon, N.; Bardner, R.; Allen-Williams, L.; Lee, J. Flight periodicity and infestation size of Sitona lineatus. Ann. Appl. Biol. 1987, 111, 271–284. [Google Scholar] [CrossRef]
- Jackson, D.J.; Macdougall, K.S. Bionomics of weevils of the genus Sitones injurious to leguminous crops in Britain. Ann. Appl. Biol. 1920, 7, 269–298. [Google Scholar] [CrossRef]
- Cárcamo, H.A.; Herle, C.E.; Lupwayi, N.Z. Sitona lineatus (Coleoptera: Curculionidae) larval feeding on Pisum sativum L. affects soil and plant nitrogen. J. Insect Sci. 2015, 15, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Cárcamo, H.A.; Vankosky, M.; Wijerathna, A.; Olfert, O.; Meers, S.; Evenden, M.L. Progress toward integrated pest management of pea leaf weevil: A review. Ann. Entomol. Soc. Am. 2018, in press. [Google Scholar]
- El-Dessouki, S. The effect of some Sitona spp. larvae on leguminous plants. J. Appl. Entomol. 1971, 67, 411–431. [Google Scholar]
- Landona, F.; Levieux, J.; Huignard, J.; Rougon, D.; Taupin, P. Feeding activity of Sitona lineatus L.(Col., Curculionidae) on Pisum sativum L.(Leguminosae) during its imaginal life. J. Appl. Entomol. 1995, 119, 515–522. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Guerrero, A. Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci. 2004, 9, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Evenden, M.; Whitehouse, C.; St. Onge, A.; Vanderark, L.; Lafontaine, J.-P.; Meers, S.; Cárcamo, H. Potential for semiochemical-based monitoring of the pea leaf weevil (Coleoptera: Curculionidae) on field pea (Fabaceae) in the Canadian prairie provinces. Can. Entomol. 2016, 148, 595–602. [Google Scholar] [CrossRef]
- Evenden, M.L. Semiochemical-based management of the pea leaf weevil, Sitona lineatus (Coleoptera: Curculionidae). Ann. Entomol. Soc. Am. 2018, in press. [Google Scholar]
- Nielsen, B.; Jensen, T. Spring dispersal of Sitona lineatus: The use of aggregation pheromone traps for monitoring. Entomol. Exp. Appl. 1993, 66, 21–30. [Google Scholar] [CrossRef]
- St. Onge, A.; Cárcamo, H.A.; Evenden, M.L. Evaluation of semiochemical-baited traps for monitoring the pea leaf weevil, Sitona lineatus (Coleoptera: Curculionidae) in field pea crops. Environ. Entomol. 2017, 47, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Blight, M.M.; Pickett, J.A.; Smith, M.C.; Wadhams, L.J. An aggregation pheromone of Sitona lineatus. Naturwissenschaften 1984, 71, 480. [Google Scholar] [CrossRef]
- Blight, M.M.; Dawson, G.; Pickett, J.A.; Wadhams, L.J. The identification and biological activity of the aggregation pheromone of Sitona lineatus. Asp. Appl. Biol. 1991, 27, 137–142. [Google Scholar]
- Landon, F.; Ferary, S.; Pierre, D.; Auger, J.; Biemont, J.; Levieux, J.; Pouzat, J. Sitona lineatus host-plant odors and their components: Effect on locomotor behavior and peripheral sensitivity variations. J. Chem. Ecol. 1997, 23, 2161–2173. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Cruz, Z.; Guerrero, A. Development of an efficient pheromone-based trapping method for the banana root borer Cosmopolites sordidus. J. Chem. Ecol. 2009, 35, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.V.P.; Fettköther, R.; Noldt, U.; Dettner, K. Capture of female Hylotrupes bajulus as influenced by trap type and pheromone blend. J. Chem. Ecol. 2005, 31, 2169–2177. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.V.P.; Gadi, N.; Taianao, A.J. Efficient sex pheromone trapping: Catching the sweetpotato weevil, Cylas formicarius. J. Chem. Ecol. 2012, 38, 846–853. [Google Scholar] [CrossRef] [PubMed]
- McVay, K.; Burrows, M.; Menalled, F.; Jones, C.; Wanner, K.; O’Neill, R. Montana Cool-Season Pulse. Production Guide; EB0210, Montana State University Extension Service; Montana State University: Bozeman, MT, USA, 2013; pp. 1–28. [Google Scholar]
- Alberta Agriculture Center. Pea Leaf Weevil Fact Sheet. Available online: http://www1.agric.gov.ab.ca/$department/deptdocs.nsf/all/agdex14738 (accessed on 14 June 2018).
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. Available online: http://www.Rproject.org (accessed on 14 January 2018).
- Biddle, A.; Smart, L.; Blight, M.; Lane, A. A monitoring system for the pea and bean weevil (Sitona lineatus). In Proceedings of the 1996 Brighton Crop Protection Conference—Pests and Diseases, Brighton, UK, 18–21 November 1996; pp. 173–178. [Google Scholar]
- Quinn, M.A.; Bezdicek, D.F.; Smart, L.E.; Martin, J. An aggregation pheromone system for monitoring pea leaf weevil (Coleoptera: Curculionidae) in the Pacific Northwest. J. Kans. Entomol. Soc. 1999, 72, 315–321. [Google Scholar]
- Valles, S.; Capinera, J.; Teal, P. Evaluation of pheromone trap design, height, and efficiency for capture of male Diaphania nitidalis (Lepidoptera: Pyralidae) in a field cage. Environ. Entomol. 1991, 20, 1274–1278. [Google Scholar] [CrossRef]
- Fisher, J.R.; O’Keefe, L.E. Seasonal migration and flight of the pea leaf weevil, Sitona lineatus (Coleoptera: Curculionidae) in northern Idaho and eastern Washington. Entomol. Exp. Appl. 1979, 26, 189–196. [Google Scholar] [CrossRef]
- I-lussain, A.; Phillips, T.W.; AliNiazee, M.T. Responses of Tribolium castaneum to different pheromone lures and traps in the laboratory. In Proceedings of the 6th International Working Conference on Stored-Product Protection, Canberra, Australia, 17–23 April 1994; pp. 406–409. [Google Scholar]
- Malo, E.A.; Cruz-Lopez, L.; Valle-Mora, J.; Virgen, A.; Sanchez, J.A.; Rojas, J.C. Evaluation of commercial pheromone lures and traps for monitoring male fall armyworm (Lepidoptera: Noctuidae) in the coastal region of Chiapas, Mexico. Fla. Entomol. 2001, 84, 659–664. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Kavallieratos, N.G.; Mazomenos, B.E. Effect of trap type, trap color, trapping location, and pheromone dispenser on captures of male Palpita unionalis (Lepidoptera: Pyralidae). J. Econ. Entomol. 2004, 97, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Lobos, E.; Occhionero, M.; Werenitzky, D.; Fernandez, J.; Gonzalez, L.M.; Rodriguez, C.; Calvo, C.; Lopez, G.; Oehlschlager, A.C. Optimization of a trap for Tuta absoluta Meyrick (Lepidoptera: Gelechiidae) and trials to determine the effectiveness of mass trapping. Neotrop. Entomol. 2013, 42, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.V.P.; Guerrero, A. New pheromones and insect control strategies. Vitam. Horm. 2010, 83, 493–519. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiments | Field Site | Trap Deployed Date | Geographical Coordinates | Total Cultivated Area (Hectares) | Golden Triangle Area | |
|---|---|---|---|---|---|---|
| Latitude | Longitude | |||||
| Trap type | Field A | 4 April 2016 | N 48°06.325′ | W 111°56.060′ | 53 | Central |
| Field B | 6 April 2016 | N 48°18.463′ | W112°12.086′ | 55 | Central | |
| Field C | 4 April 2016 | N 48°11.398′ | W 111°40.952′ | 259 | Central | |
| Field D | 11 April 2016 | N48°06.351′ | W111°19.477′ | 18 | Southern | |
| Field E | 18 April 2016 | N 47°59.500′ | W 112°05.436′ | 132 | Southern | |
| Field F | 16 May 2016 | N 48°50.347′ | W 111°51.713′ | 61 | Northern | |
| Lure type | Pea | |||||
| Field A | 4 April 2016 | N 48°06.325′ | W111°56.060′ | 53 | Central | |
| Field B | 4 April 2016 | N 48°11.398′ | W 111°40.952′ | 259 | Central | |
| Field C | 18 April 2016 | N 47°59.500′ | W 112°05.436′ | 132 | Southern | |
| Field D | 11 April 2016 | N 48°06.351′ | W111°19.477′ | 18 | Southern | |
| Field E | 11 April 2016 | N 47°59.220′ | W 111°28.076′ | 259 | Southern | |
| Field F | 19 April 2016 | N 47°50.518′ | W 111°41.286′ | 16 | Southern | |
| Field G | 14 April 2016 | N 48°39.628′ | W 111°08.545′ | 216 | Northern | |
| Field H | 22 April 2016 | N 48°50.347′ | W 111°51.713′ | 61 | Northern | |
| Field I | 16 April 2016 | N 48°50.475′ | W 111°47.769′ | 97 | Northern | |
| Field J | 22 April 2016 | N 48°18.458′ | W 111°55.529′ | 129 | Northern | |
| Lentil | ||||||
| Field A | 11 April 2016 | N 48°06.831′ | W 111°55.475′ | 32 | Central | |
| Field B | 2 May 2016 | N 48°46.131′ | W 110°54.192′ | 55 | Central | |
| Field C | 4 April 2016 | N 48°01.785′ | W 111°42.553′ | 20 | Southern | |
| Field D | 9 April 2016 | N 47°57.360′ | W 111°54.560′ | 129 | Southern | |
| Field E | 13 April 2016 | N 47°56.0541′ | W 112°01.069′ | 65 | Southern | |
| Field F | 11 May 2016 | N 47°48.452′ | W 111°30.321′ | 111 | Southern | |
| Trap placement | Field A | 4 April 2016 | N 48°06.325′ | W 111°56.060′ | 53 | Central |
| Field B | 6 April 2016 | N 48°18.463′ | W 112°12.086′ | 55 | Central | |
| Field C | 10 April 2016 | N 48°11.398′ | W 111°40.952′ | 259 | Central | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reddy, G.V.P.; Shrestha, G.; Miller, D.A.; Oehlschlager, A.C. Pheromone-Trap Monitoring System for Pea Leaf Weevil, Sitona lineatus: Effects of Trap Type, Lure Type and Trap Placement within Fields. Insects 2018, 9, 75. https://doi.org/10.3390/insects9030075
Reddy GVP, Shrestha G, Miller DA, Oehlschlager AC. Pheromone-Trap Monitoring System for Pea Leaf Weevil, Sitona lineatus: Effects of Trap Type, Lure Type and Trap Placement within Fields. Insects. 2018; 9(3):75. https://doi.org/10.3390/insects9030075
Chicago/Turabian StyleReddy, Gadi V. P., Govinda Shrestha, Debra A. Miller, and A. Cameron Oehlschlager. 2018. "Pheromone-Trap Monitoring System for Pea Leaf Weevil, Sitona lineatus: Effects of Trap Type, Lure Type and Trap Placement within Fields" Insects 9, no. 3: 75. https://doi.org/10.3390/insects9030075
APA StyleReddy, G. V. P., Shrestha, G., Miller, D. A., & Oehlschlager, A. C. (2018). Pheromone-Trap Monitoring System for Pea Leaf Weevil, Sitona lineatus: Effects of Trap Type, Lure Type and Trap Placement within Fields. Insects, 9(3), 75. https://doi.org/10.3390/insects9030075