Monitoring and Biosurveillance Tools for the Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae)

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. 2016 Trap and Lure Comparisons
2.2. 2017 Lure Comparison using Clear Sticky Traps
2.3. Statistical Analyses
3. Results
3.1. Relative Population Density Classifications
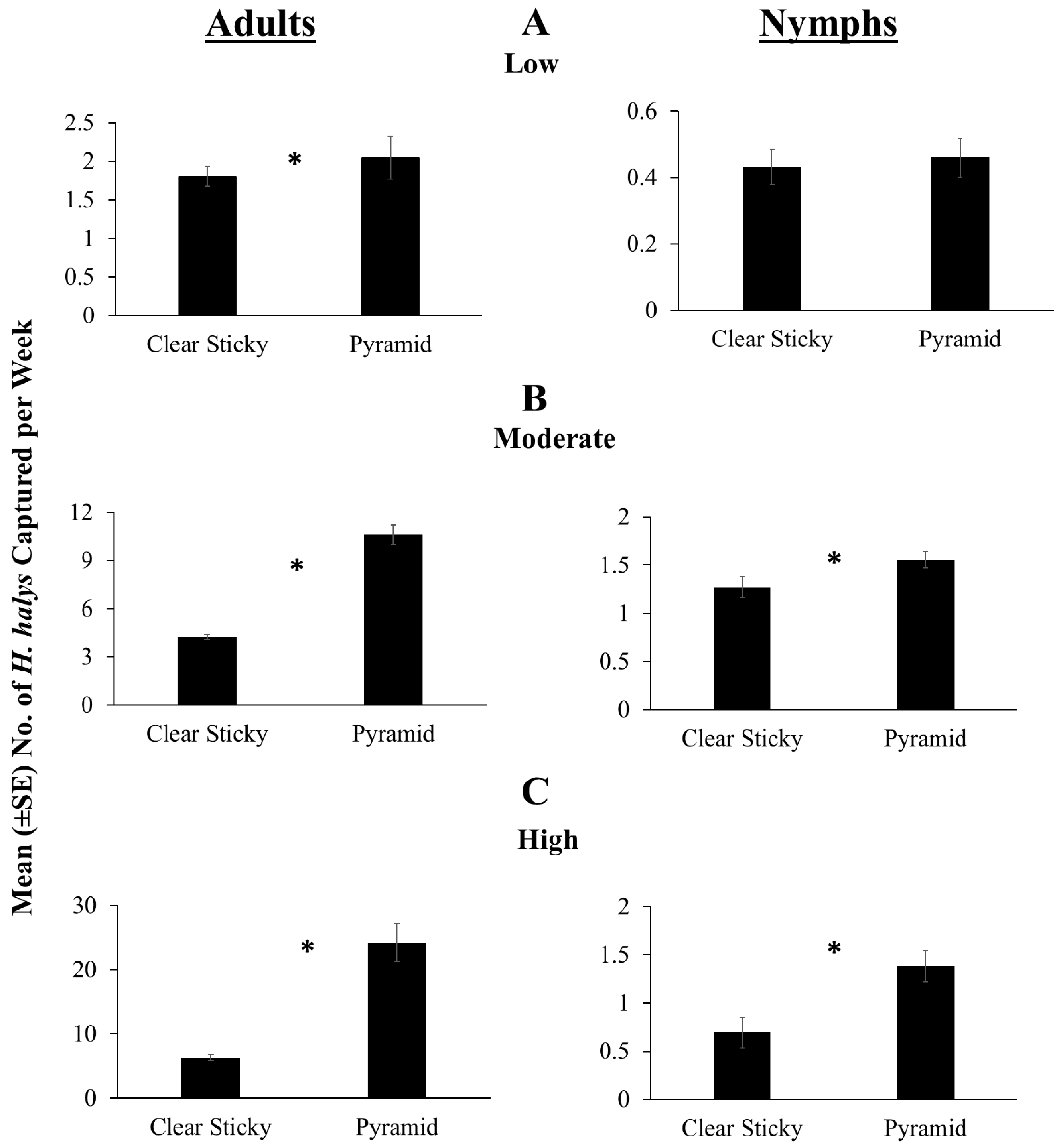
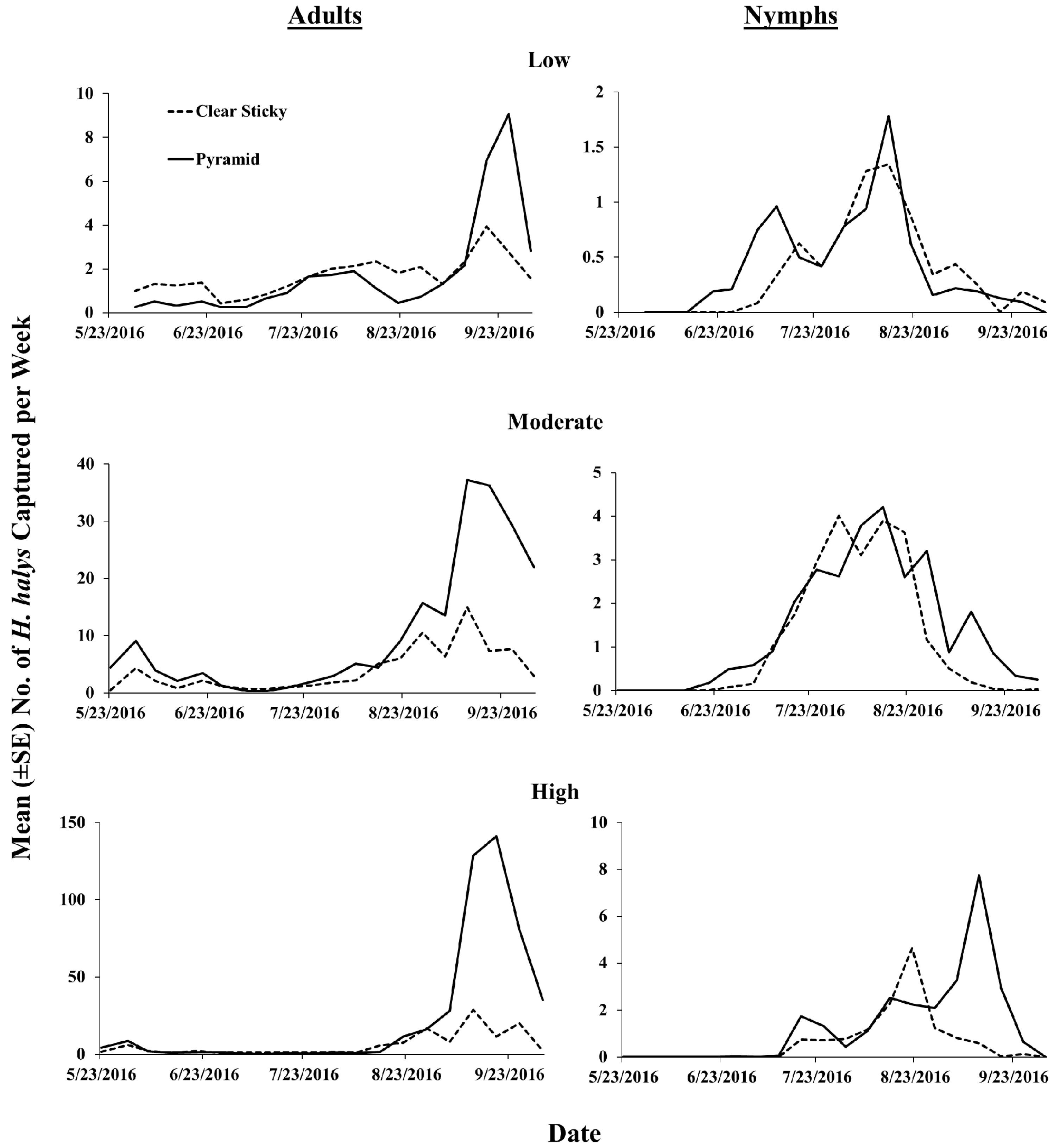
3.2. 2016 Trap Design Comparison
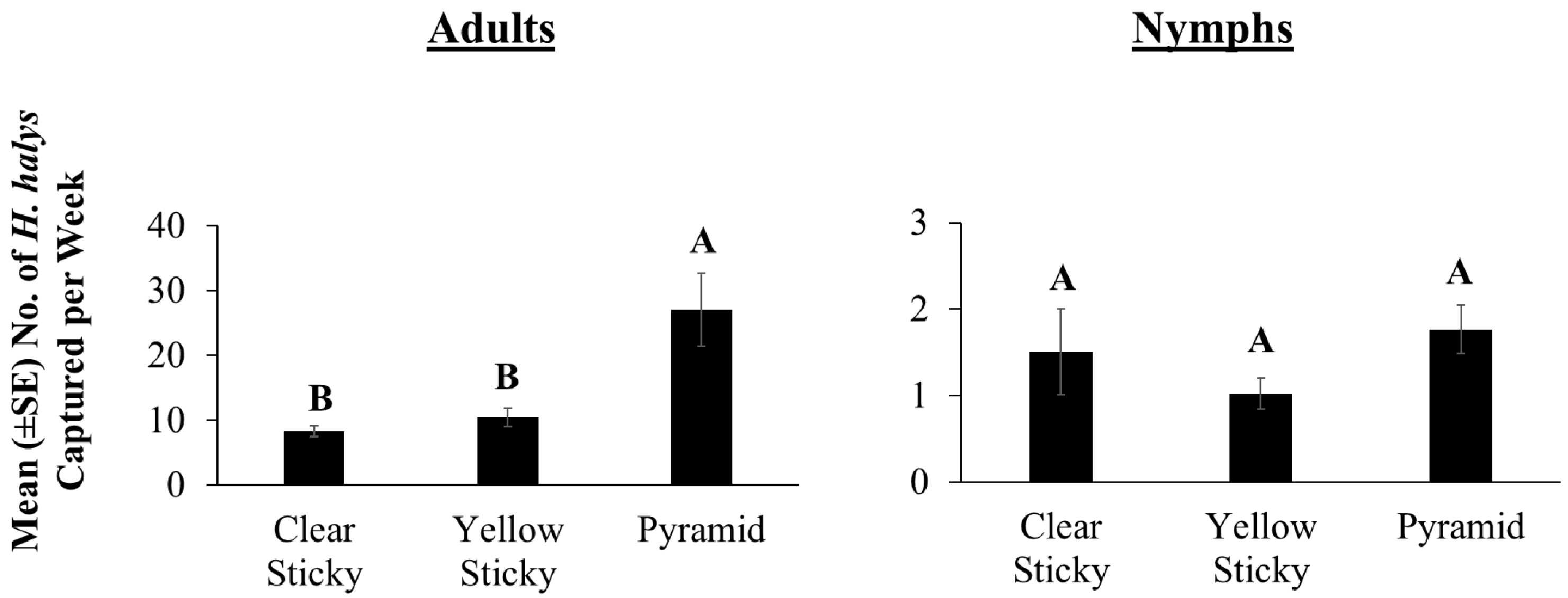
3.3. 2016 Lure Comparison
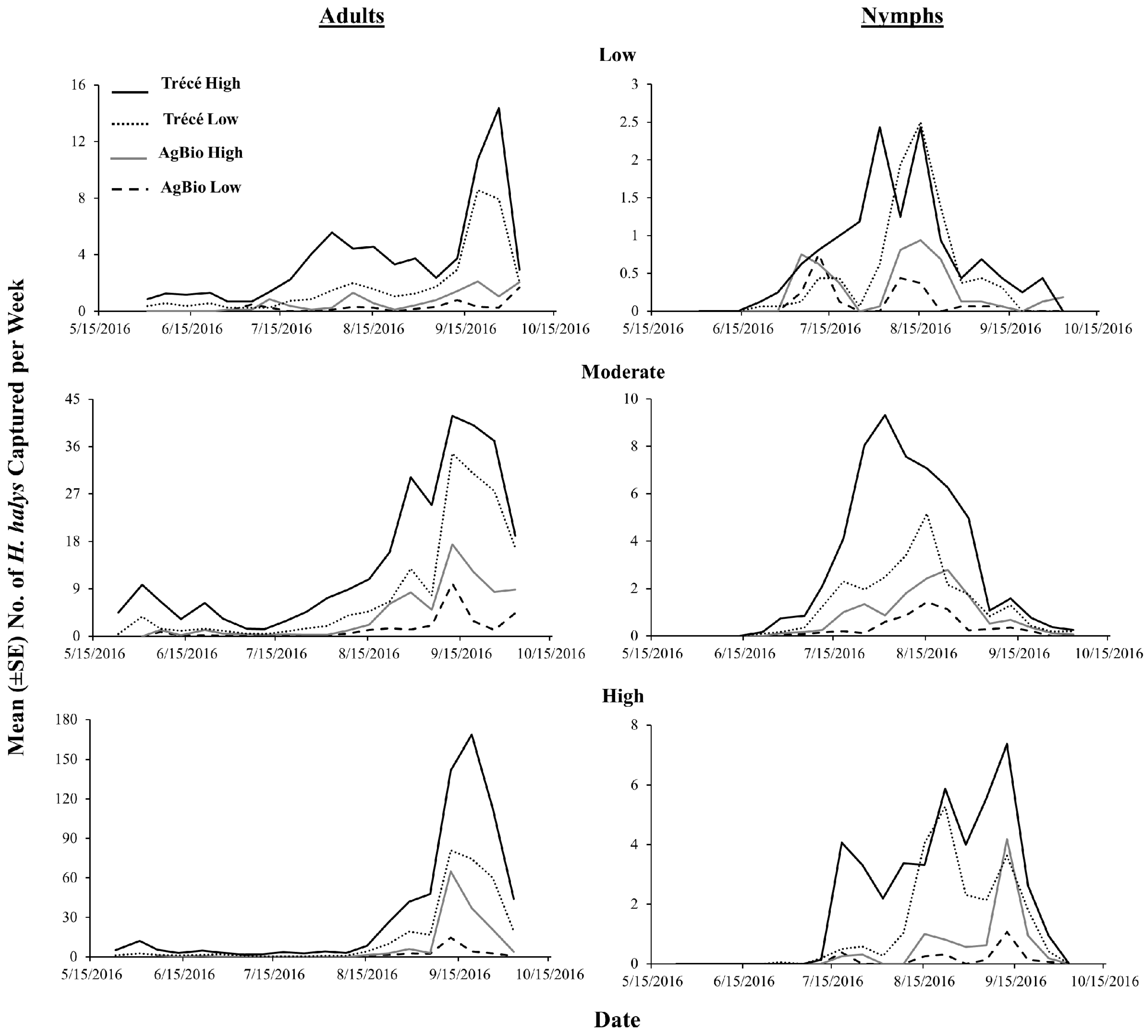
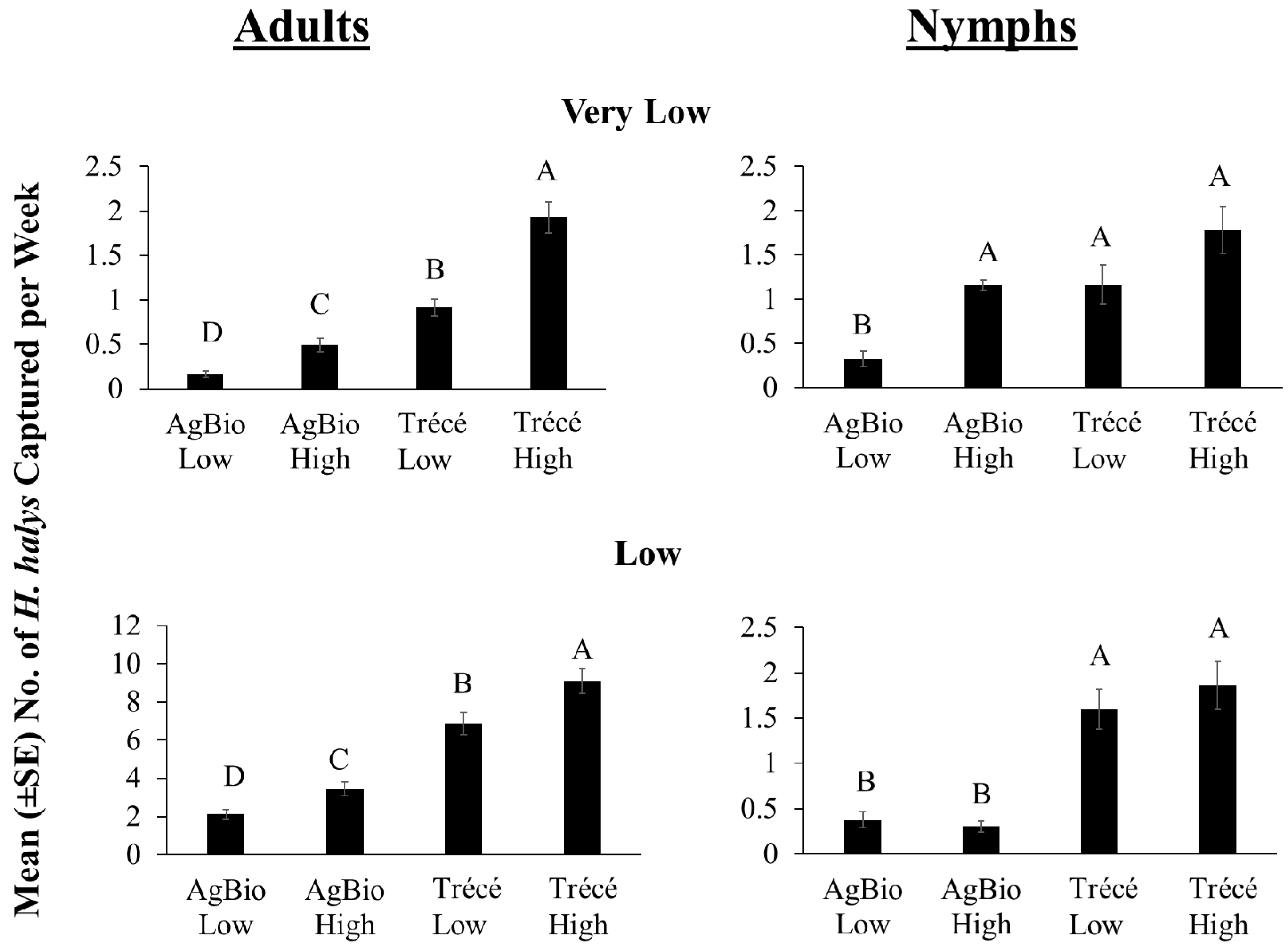
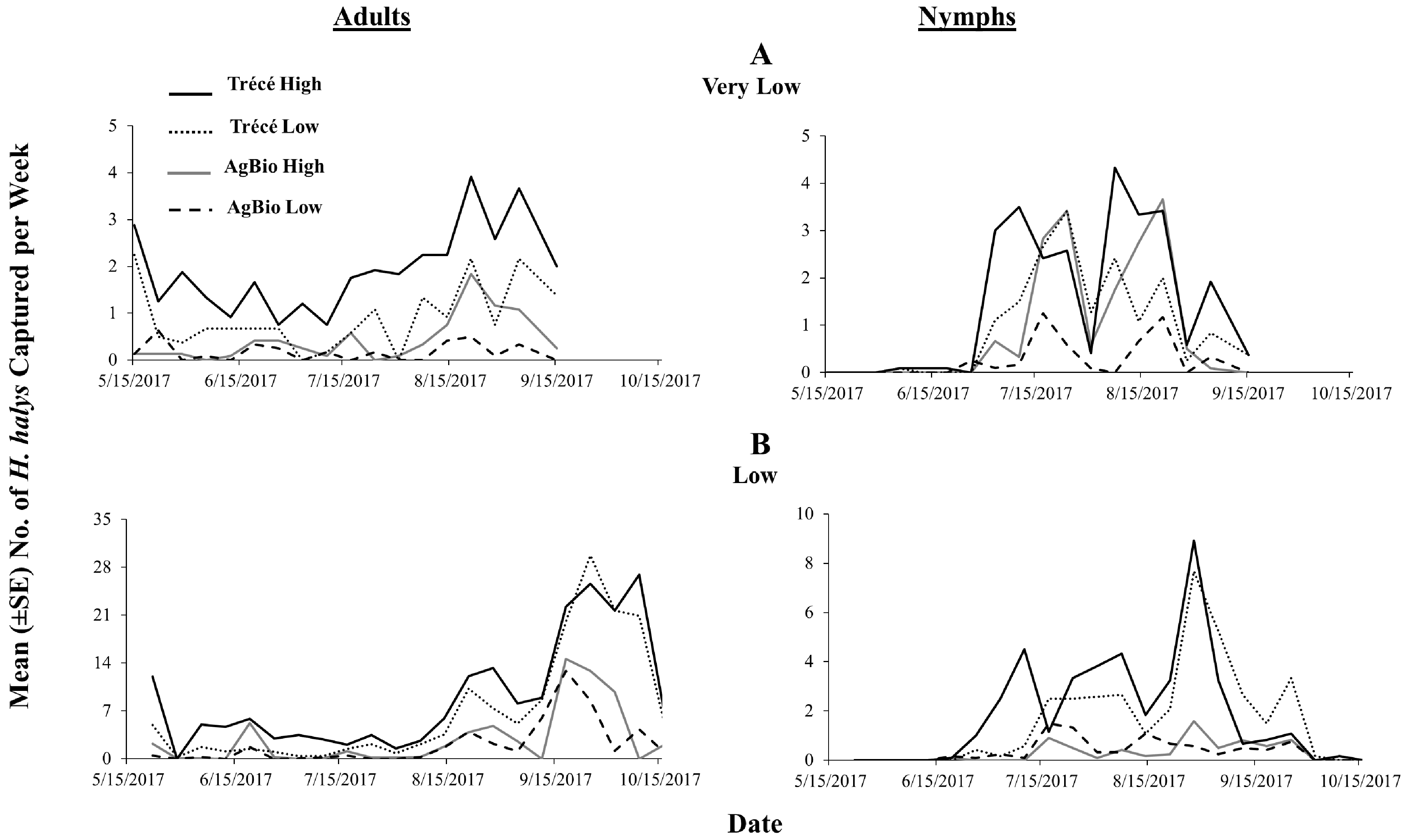
3.4. 2017 Lure Comparisons with Sticky Traps
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dent, D. Insect Pest Management; CABI: Wallingford, UK, 2000. [Google Scholar]
- Jarrad, F.; Low-Choy, S.; Mengersen, K. Biosecurity Surveillance: Quantitative Approaches; CABI: Wallingford, UK, 2015. [Google Scholar]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: history, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [PubMed]
- Fogain, R.; Graff, S. First records of the invasive pest, Halyomorpha halys (Hemiptera: Pentatomidae), in Ontario and Quebec. Proc. Entomol. Soc. Ont. 2011, 142, 45–48. [Google Scholar]
- Wermelinger, B.; Wyniger, D.; Forster, B. First records of an invasive bug in Europe: Halyomorpha halys Stal (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Mitt. Schweiz. Entomol. Ges. 2008, 81, 1–8. [Google Scholar]
- Maistrello, L.; Dioli, P.; Bariselli, M.; Mazzoli, G.L.; Giacalone-Forini, I. Citizen science and early detection of invasive species: phenology of first occurrences of Halyomorpha halys in Southern Europe. Biol. Invasions 2016, 18, 3109–3116. [Google Scholar] [CrossRef]
- Gapon, D. First records of the brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Heteroptera, Pentatomidae) in Russia, Abkhazia, and Georgia. Entomol. Rev. 2016, 96, 1086–1088. [Google Scholar] [CrossRef]
- Macavei, L.I.; Bâeţan, R.; Oltean, I.; Florian, T.; Varga, M.; Costi, E.; Maistrello, L. First detection of Halyomorpha halys Stål, a new invasive species with a high potential of damage on agricultural crops in Romania. Lucrări Ştiinţifice Ser. Agron. 2015, 58, 105–108. [Google Scholar]
- Faúndez, E.; Rider, D. The brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae) in Chile. Arq. Entomol. 2017, 17, 305–307. [Google Scholar]
- Zhu, G.; Bu, W.; Gao, Y.; Liu, G. Potential geographic distribution of brown marmorated stink bug invasion (Halyomorpha halys). PLoS ONE 2012, 7, e31246. [Google Scholar] [CrossRef] [PubMed]
- Kriticos, D.J.; Kean, J.M.; Phillips, C.B.; Senay, S.D.; Acosta, H.; Haye, T. The potential global distribution of the brown marmorated stink bug, Halyomorpha halys, a critical threat to plant biosecurity. J. Pest Sci. 2017, 90, 1033–1043. [Google Scholar] [CrossRef]
- Ministry for Primary Industries, Protection and Response. Available online: https://www.mpi.govt.nz/protection-and-response/responding/alerts/brown-marmorated-stink-bug/ (accessed on 23 April 2018).
- Leskey, T.C.; Wright, S.E.; Short, B.D.; Khrimian, A. Development of behaviorally based monitoring tools for the brown marmorated stink bug, Halyomorpha halys (Stål) (Heteroptera: Pentatomidae) in commercial tree fruit orchards. J. Entomol. Sci. 2012, 47, 76–85. [Google Scholar] [CrossRef]
- Joseph, S.V.; Bergh, J.C.; Wright, S.E.; Leskey, T.C. Factors affecting captures of brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae), in baited pyramid traps. J. Entomol. Sci. 2013, 48, 43–51. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Holmstrom, K.; Hamilton, G.C.; Cambridge, J.; Ingerson-Mahar, J. Use of black light traps to monitor the abundance, spread, and flight behavior of Halyomorpha halys (Hemiptera: Pentatomidae). J. Econ. Entomol. 2013, 106, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Morrison, W.R.I.; Cullum, J.P.; Leskey, T.C. Evaluation of trap designs and deployment strategies for capturing Halyomorpha halys (Hemiptera: Pentatomidae). J. Econ. Entomol. 2015, 108, 1683–1692. [Google Scholar] [CrossRef] [PubMed]
- Rice, K.B.; Morrison, W.R.I.; Short, B.D.; Acebes-Doria, A.L.; Bergh, C.J.; Leskey, T.C. Improved Trap Designs and Retention Mechanisms for Halyomorpha halys (Hemiptera: Pentatomidae). J. Econ. Entomol. 2018. [Google Scholar] [CrossRef]
- Weber, D.C.; Morrison, W.R., III; Khrimian, A.; Rice, K.B.; Leskey, T.C.; Rodriguez-Saona, C.; Nielsen, A.L.; Blaauw, B.R. Chemical ecology of Halyomorpha halys: discoveries and applications. J. Pest Sci. 2017, 90, 989–1008. [Google Scholar] [CrossRef]
- Khrimian, A.; Zhang, A.; Weber, D.C.; Ho, H.-Y.; Aldrich, J.R.; Vermillion, K.E.; Siegler, M.A.; Shirali, S.; Guzman, F.; Leskey, T.C. Discovery of the aggregation pheromone of the brown marmorated stink bug (Halyomorpha halys) through the creation of stereoisomeric libraries of 1-bisabolen-3-ols. J. Nat. Prod. 2014, 77, 1708–1717. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.C.; Leskey, T.C.; Walsh, G.C.; Khrimian, A. Synergy of aggregation pheromone with methyl (E,E,Z)-2, 4, 6-decatrienoate in attraction of Halyomorpha halys (Hemiptera: Pentatomidae). J. Econ. Entomol. 2014, 107, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Leskey, T.C.; Agnello, A.; Bergh, J.C.; Dively, G.P.; Hamilton, G.C.; Jentsch, P.; Khrimian, A.; Krawczyk, G.; Kuhar, T.P.; Lee, D.-H.; et al. Attraction of the invasive Halyomorpha halys (Hemiptera: Pentatomidae) to traps baited with semiochemical stimuli across the United States. Environ. Entomol. 2015, 44, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Short, B.D.; Khrimian, A.; Leskey, T.C. Pheromone-based decision support tools for management of Halyomorpha halys in apple orchards: Development of a trap-based treatment threshold. J. Pest Sci. 2017, 90, 1191–1204. [Google Scholar] [CrossRef]
- Toyama, M.; Kishimoto, H.; Mishiro, K.; Nakano, R.; Ihara, F. Sticky traps baited with synthetic aggregation pheromone predict fruit orchard infestations of Plautia stali (Hemiptera: Pentatomidae). J. Econ. Entomol. 2015, 108, 2366–2372. [Google Scholar] [CrossRef] [PubMed]
- Prokopy, R.J.; Owens, E.D. Visual detection of plants by herbivorous insects. Annu. Rev. Entomol. 1983, 28, 337–364. [Google Scholar] [CrossRef]
- O’hara, R.B.; Kotze, D.J. Do not log-transform count data. Methods Ecol. Evol. 2010, 1, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Nixon, L.J.; Morrison, W.R.; Rice, K.B.; Brockerhoff, E.G.; Leskey, T.C.; Guzman, F.; Khrimian, A.; Goldson, S.; Rostás, M. Identification of volatiles released by diapausing brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). PLoS ONE 2018, 13, e0191223. [Google Scholar] [CrossRef] [PubMed]
- Morrison, W.R.; Acebes-Doria, A.; Ogburn, E.; Kuhar, T.P.; Walgenbach, J.F.; Bergh, J.C.; Nottingham, L.; Dimeglio, A.; Hipkins, P.; Leskey, T.C. Behavioral response of the brown marmorated stink bug (Hemiptera: Pentatomidae) to semiochemicals deployed inside and outside anthropogenic structures during the overwintering period. J. Econ. Entomol. 2017, 110, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- Morrison, W.R.; Milonas, P.; Kapantaidaki, D.E.; Cesari, M.; di Bella, E.; Guidetti, R.; Haye, T.; Maistrello, L.; Moraglio, S.T.; Piemontese, L.; et al. Attraction of Halyomorpha halys (Hemiptera: Pentatomidae) haplotypes in North America and Europe to baited traps. Sci. Rep. 2017, 7, 16941. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.; Storer, A.; Fraser, I.; Mastro, V. Efficacy of trap and lure types for detection of Agrilus planipennis (Col., Buprestidae) at low density. J. Appl. Entomol. 2010, 134, 296–302. [Google Scholar] [CrossRef]
- Ryall, K.L.; Fidgen, J.G.; Turgeon, J.J. Detectability of the emerald ash borer (Coleoptera: Buprestidae) in asymptomatic urban trees by using branch samples. Environ. Entomol. 2011, 40, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Valentin, R.E.; Fonseca, D.M.; Nielsen, A.L.; Leskey, T.C.; Lockwood, J.L. Early detection of invasive exotic insect infestations using eDNA from crop surfaces. Front. Ecol. Environ. 2018, 265–270. [Google Scholar] [CrossRef]
- Acebes-Doria, A.L.; Leskey, T.C.; Bergh, C.J. Development and comparison of trunk traps to monitor movement of Halyomorpha halys nymphs on host trees. Entomol. Exp. Appl. 2016, 158, 44–53. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Morrison, W.R.; Mathews, C.; Leskey, T.C.; Nielsen, A.L. Measuring host plant selection and retention of Halyomorpha halys by a trap crop. Environ. Entomol. 2017, 163, 197–208. [Google Scholar] [CrossRef]
- Morrison, W.R.; Lee, D.H.; Short, B.D.; Khrimian, A.; Leskey, T.C. Establishing the behavioral basis for an attract-and-kill strategy to manage the invasive Halyomorpha halys in apple orchards. J. Pest Sci. 2016, 89, 81–96. [Google Scholar] [CrossRef]
- Hoelmer, K.A.; Simmons, A.M. Yellow sticky trap catches of parasitoids of Bemisia tabaci (Hemiptera: Aleyrodidae) in vegetable crops and their relationship to in-field populations. Environ. Entomol. 2008, 37, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Böckmann, E.; Meyhöfer, R. Sticky trap monitoring of a pest–predator system in glasshouse tomato crops: are available trap colours sufficient? J. Appl. Entomol. 2017, 141, 339–351. [Google Scholar] [CrossRef]
- Leskey, T.C.; Khrimian, A.; Weber, J.D.; Aldrich, C.; Short, B.D.; Lee, D.-H.; Morrison, W.R., III. Behavioral responses of the invasive Halyomorpha halys (Stål) to traps baited with stereoisomeric mixtures of 10, 11-epoxy-1-bisabolen-3-OL. J. Chem. Ecol. 2015, 41, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Rice, K.B.; Bedoukian, R.H.; Hamilton, G.C.; Jentsch, P.; Khrimian, A.; MacLean, P.; Morrison, W.R.; Short, B.D.; Shrewsbury, P.; Weber, D.C. Enhanced response of Halyomorpha halys (Hemiptera: Pentatomidae) to its aggregation pheromone with ethyl decatrienoate. J. Econ. Entomol. 2017, 11, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Morrison, W.R.; Allen, M.; Leskey, T.C. Behavioural response of the invasive Halyomorpha halys (Hemiptera: Pentatomidae) to host plant stimuli augmented with semiochemicals in the field. Agric. For. Entomol. 2018, 20, 62–72. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locations | GPS Coordinates | Trapping Period (Relative Population Density) | |
|---|---|---|---|
| West Virginia | 2016 Study | 2017 Study | |
| Jefferson | 39°22′52.41” N 77°52′49.81” W | 23 May–3 October (M) | 22 May–16 October (L) |
| Burns | 39°14′49.30” N 77°55′5.59” W | 23 May–3 October (M) | - |
| USGS * | 39°21′16.89” N 77°54′39.89” W | 23 May–3 October (H) | 22 May–16 October (L) |
| Link * | 39°20′31.12” N 77°53′45.25” W | 23 May–3 October (M) | - |
| Tabb | 39°20′41.52” N 77°55′27.78” W | 23 May–3 October (M) | - |
| Twin Ridge * | 39°22′42.59” N 77°50′42.81” W | 23 May–3 October (H) | 22 May–16 October (L) |
| Virginia | |||
| Eastern Shore | 37°35′23.44′′ N 75°49′11.82′′ W | 31 May–3 October (L) | 7 Jun–5 September (VL) |
| 37°38′21.18′′ N 77°58′36.93′′ W | 6 June–3 October (M) | - | |
| Goochland 2 | 37°41′49.65′′ N 78°01′8.89′′ W | 6 June–3 October(L) | - |
| Goochland 3 | 37°40′45.97′′ N 77°53′32.29′′ W | 6 June–3 October (M) | - |
| VA Site 1 | 37°12′20.49′′ N 80°33′53.60′′ W | 31 May–26 September (M) | 17 May–13 September (VL) |
| VA Site 2 | 37°11′28.07′′ N 80°34′42.73′′ W | 31 May–26 September (M) | 17 May–13 September (VL) |
| Lure Type & Population Density | Adults | Nymphs | ||||
|---|---|---|---|---|---|---|
| r | df | p | r | df | p | |
| Trécé Low | ||||||
| Low | 0.817 | 37 | 0.0001 | 0.832 | 37 | 0.0001 |
| Moderate | 0.739 | 152 | 0.0001 | 0.544 | 152 | 0.0001 |
| High | 0.803 | 40 | 0.0001 | 0.559 | 40 | 0.0002 |
| Trécé High | ||||||
| Low | 0.782 | 37 | 0.0001 | 0.661 | 37 | 0.0001 |
| Moderate | 0.636 | 152 | 0.0001 | 0.695 | 152 | 0.0001 |
| High | 0.801 | 40 | 0.0001 | 0.615 | 40 | 0.0001 |
| AgBio Low | ||||||
| Low | 0.822 | 25 | 0.0001 | 0.433 | 25 | 0.0305 |
| Moderate | 0.759 | 130 | 0.0001 | 0.467 | 130 | 0.0001 |
| High | 0.752 | 36 | 0.0001 | 0.114 | 36 | 0.507 |
| AgBio High | ||||||
| Low | 0.820 | 25 | 0.0001 | 0.741 | 25 | 0.0001 |
| Moderate | 0.719 | 130 | 0.0001 | 0.451 | 130 | 0.0001 |
| High | 0.726 | 36 | 0.0001 | 0.287 | 36 | 0.090 |
| Lure Type & Population Density | 2016 | Lure Type & Population Density | 2017 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adults | Nymphs | Adults | Nymphs | ||||||||||
| r | df | r | df | r | df | r | df | p | |||||
| Trécé lures | Trécé lures | ||||||||||||
| Low | 0.812 | 37 | 0.0001 | 0.696 | 37 | 0.0001 | Very Low | 0.598 | 58 | 0.0001 | 0.825 | 50 | 0.0001 |
| Moderate | 0.87 | 152 | 0.0001 | 0.736 | 152 | 0.0001 | Low | 0.889 | 66 | 0.0001 | 0.788 | 66 | 0.0001 |
| High | 0.889 | 40 | 0.0001 | 0.647 | 40 | 0.0001 | - | - | - | - | - | - | - |
| AgBio lures | |||||||||||||
| Low | 0.618 | 25 | 0.001 | 0.741 | 25 | 0.0001 | Very Low | 0.386 | 50 | 0.0056 | 0.68 | 50 | 0.0001 |
| Moderate | 0.787 | 130 | 0.0001 | 0.719 | 130 | 0.0001 | Low | 0.855 | 66 | 0.0001 | 0.553 | 66 | 0.0001 |
| High | 0.931 | 36 | 0.0001 | 0.147 | 36 | 0.3934 | - | - | - | - | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acebes-Doria, A.L.; Morrison, W.R., III; Short, B.D.; Rice, K.B.; Bush, H.G.; Kuhar, T.P.; Duthie, C.; Leskey, T.C. Monitoring and Biosurveillance Tools for the Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae). Insects 2018, 9, 82. https://doi.org/10.3390/insects9030082
Acebes-Doria AL, Morrison WR III, Short BD, Rice KB, Bush HG, Kuhar TP, Duthie C, Leskey TC. Monitoring and Biosurveillance Tools for the Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae). Insects. 2018; 9(3):82. https://doi.org/10.3390/insects9030082
Chicago/Turabian StyleAcebes-Doria, Angelita L., William R. Morrison, III, Brent D. Short, Kevin B. Rice, Hayley G. Bush, Thomas P. Kuhar, Catherine Duthie, and Tracy C. Leskey. 2018. "Monitoring and Biosurveillance Tools for the Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae)" Insects 9, no. 3: 82. https://doi.org/10.3390/insects9030082
APA StyleAcebes-Doria, A. L., Morrison, W. R., III, Short, B. D., Rice, K. B., Bush, H. G., Kuhar, T. P., Duthie, C., & Leskey, T. C. (2018). Monitoring and Biosurveillance Tools for the Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae). Insects, 9(3), 82. https://doi.org/10.3390/insects9030082