
PGC-1α Methylation, miR-23a, and miR-30e Expression as Biomarkers for Exercise- and Diet-Induced Mitochondrial Biogenesis in Capillary Blood from Healthy Individuals: A Single-Arm Intervention

 , and
, and 
Abstract
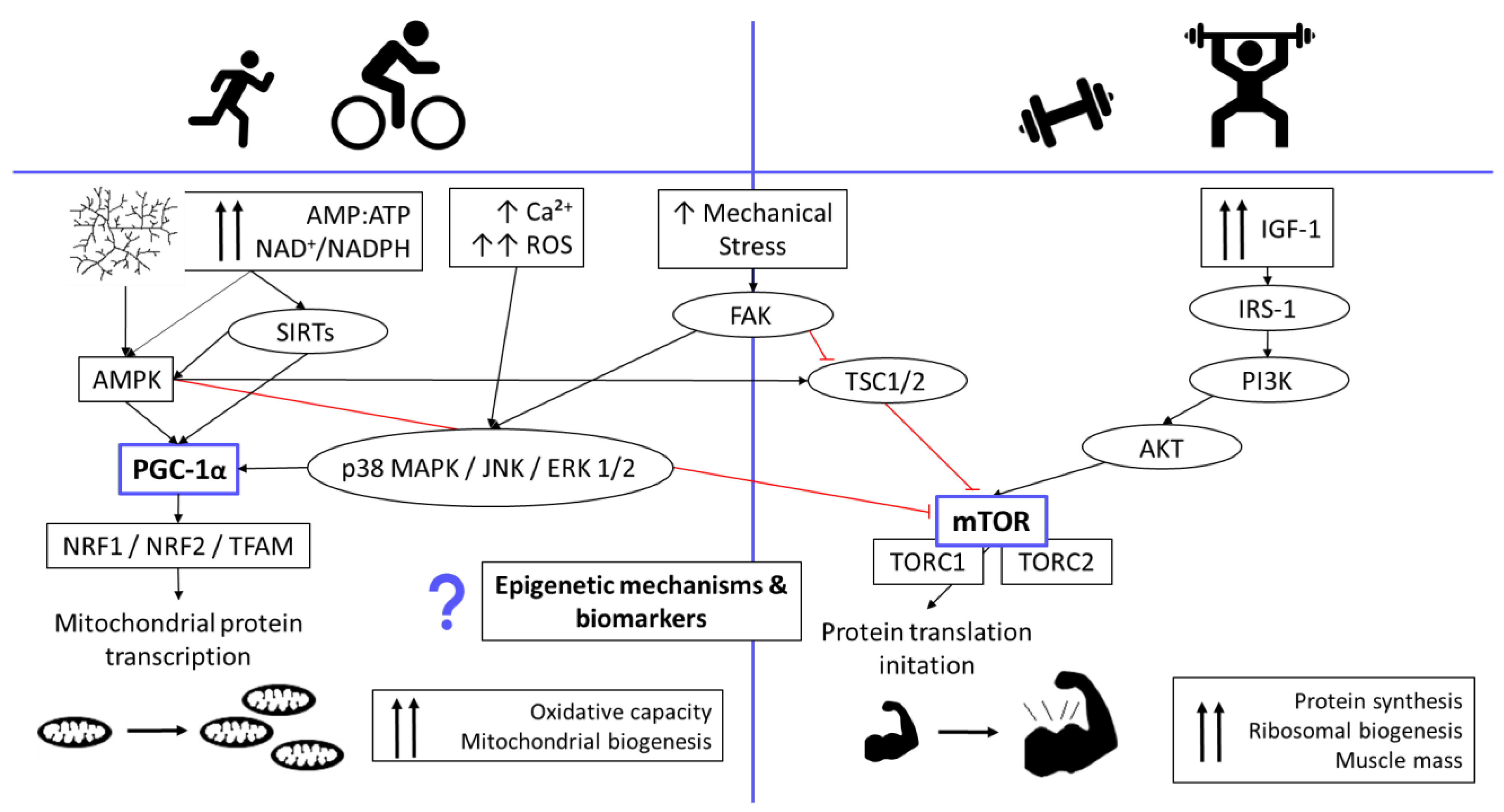
:1. Introduction

2. Materials and Methods
2.1. Subjects and Experimental Design
2.2. Training Plan
2.3. Dried Blood Spot Sample Preparation (RNA and DNA Extraction)
2.4. Bisulfite Conversion and Pyrosequencing
2.5. Real-Time Quantitaive PCR (qPCR)
2.6. Statistical Analysis
3. Results
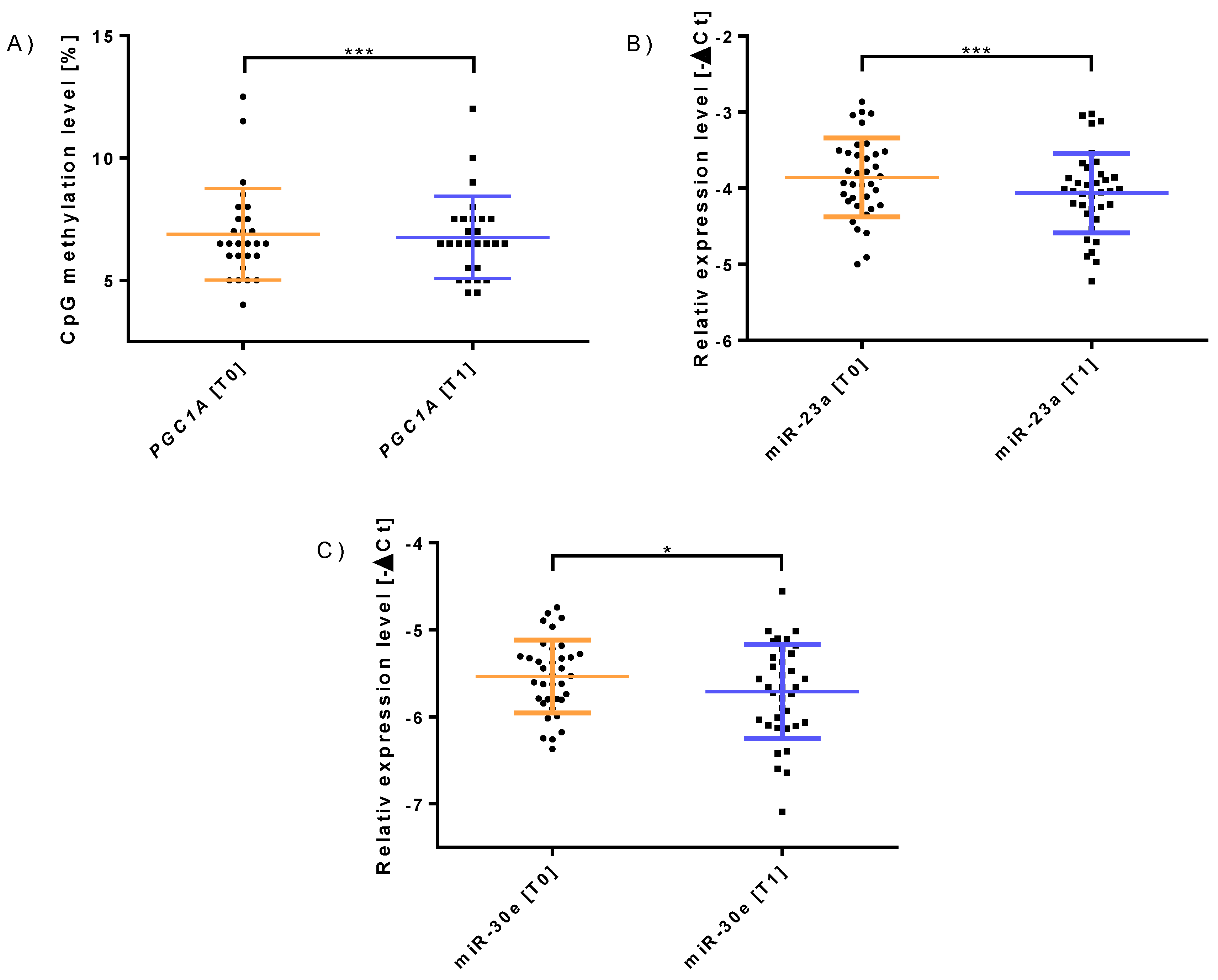
3.1. Molecular and Physiological Changes Following 12 Weeks of Concurrent Training
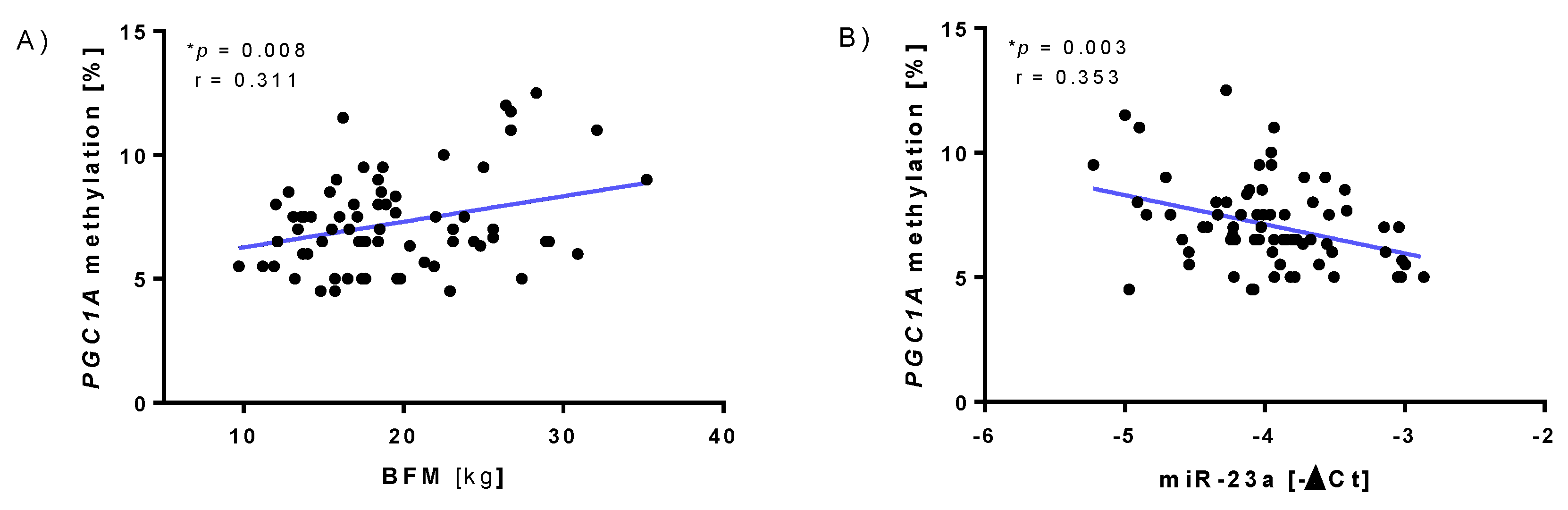
3.2. Correlations between Eating and Lifestyle Habits and Anthropometric Data with Selected Mitochondria- and Exercise-Related Markers Independent of the Intervention
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Methenitis, S. A Brief Review on Concurrent Training: From Laboratory to the Field. Sports 2018, 6, 127. [Google Scholar] [CrossRef] [Green Version]
- Simoes, D.C.M.; Vogiatzis, I. Can Muscle Protein Metabolism Be Specifically Targeted by Exercise Training in COPD? J. Thorac. Dis. 2018, 10, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Jornayvaz, F.R.; Shulman, G.I. Regulation of Mitochondrial Biogenesis. Essays Biochem. 2010, 47, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Valero, T. Mitochondrial Biogenesis: Pharmacological Approaches. Curr. Pharm. Des. 2014, 20, 5507–5509. [Google Scholar] [CrossRef] [PubMed]
- Ristow, M.; Schmeisser, K. Mitohormesis: Promoting Health and Lifespan by Increased Levels of Reactive Oxygen Species (ROS). Dose-Response 2014, 12, 288–341. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Finkel, T. Mitohermesis. Cell Metab. 2014, 19, 757–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irrcher, I.; Adhihetty, P.J.; Joseph, A.M.; Ljubicic, V.; Hood, D.A. Regulation of Mitochondrial Biogenesis in Muscle by Endurance Exercise. Sport. Med. 2003, 33, 783–793. [Google Scholar] [CrossRef]
- Chodari, L.; Aytemir, M.D.; Vahedi, P.; Alipour, M.; Vahed, S.Z.; Khatibi, S.M.H.; Ahmadian, E.; Ardalan, M.; Eftekhari, A. Targeting Mitochondrial Biogenesis with Polyphenol Compounds. Oxid. Med. Cell. Longev. 2021, 2021, 4946711. [Google Scholar] [CrossRef]
- Liang, H.; Ward, W.F. PGC-1α: A Key Regulator of Energy Metabolism. Am. J. Physiol.-Adv. Physiol. Educ. 2006, 30, 145–151. [Google Scholar] [CrossRef]
- Lin, J.; Wu, H.; Tarr, P.T.; Zhang, C.Y.; Wu, Z.; Boss, O.; Michael, L.F.; Puigserver, P.; Isotani, E.; Olson, E.N.; et al. Transcriptional Co-Activator PGC-1α Drives the Formation of Slow-Twitch Muscle Fibres. Nature 2002, 418, 797–801. [Google Scholar] [CrossRef]
- Hawley, J.A.; Hargreaves, M.; Joyner, M.J.; Zierath, J.R. Integrative Biology of Exercise. Cell 2014, 159, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groennebaek, T.; Vissing, K. Impact of Resistance Training on Skeletal Muscle Mitochondrial Biogenesis, Content, and Function. Front. Physiol. 2017, 8, 713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacques, M.; Hiam, D.; Craig, J.; Barrès, R.; Eynon, N.; Voisin, S. Epigenetic Changes in Healthy Human Skeletal Muscle Following Exercise– a Systematic Review. Epigenetics 2019, 14, 633–648. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, S.A.; Ambrosini, S.; Lüscher, T.; Paneni, F.; Costantino, S. Epigenetic Control of Mitochondrial Function in the Vasculature. Front. Cardiovasc. Med. 2020, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Hunter, D.J.; James, L.; Hussey, B.; Wadley, A.J.; Lindley, M.R.; Mastana, S.S. Impact of Aerobic Exercise and Fatty Acid Supplementation on Global and Gene-Specific DNA Methylation. Epigenetics 2019, 14, 294–309. [Google Scholar] [CrossRef] [Green Version]
- Silver, J.; Wadley, G.; Lamon, S. Mitochondrial Regulation in Skeletal Muscle: A Role for Non-Coding RNAs? Exp. Physiol. 2018, 103, 1132–1144. [Google Scholar] [CrossRef] [Green Version]
- Kirby, T.J.; McCarthy, J.J. MicroRNAs in Skeletal Muscle Biology and Exercise Adaptation. Free Radic. Biol. Med. 2013, 64, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Chen, X.; Chen, D.; Yu, B.; Li, M.; He, J.; Huang, Z. Regulation of Skeletal Myogenesis by MicroRNAs. J. Cell. Physiol. 2020, 235, 87–104. [Google Scholar] [CrossRef]
- Jia, H.; Zhao, Y.; Li, T.; Zhang, Y.; Zhu, D. MiR-30e Is Negatively Regulated by Myostatin in Skeletal Muscle and Is Functionally Related to Fiber-Type Composition. Acta Biochim. Biophys. Sin. 2017, 49, 392–399. [Google Scholar] [CrossRef] [Green Version]
- Murlasits, Z.; Kneffel, Z.; Thalib, L. The Physiological Effects of Concurrent Strength and Endurance Training Sequence: A Systematic Review and Meta-Analysis. J. Sports Sci. 2018, 36, 1212–1219. [Google Scholar] [CrossRef]
- Cadore, E.L.; Izquierdo, M. New Strategies for the Concurrent Strength-, Power-, and Endurance-Training Prescription in Elderly Individuals. J. Am. Med. Dir. Assoc. 2013, 14, 623–624. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, J.; Millet, G.P.; Place, N.; Kayser, B.; Zanou, N. The Muscle-Brain Axis and Neurodegenerative Diseases: The Key Role of Mitochondria in Exercise-Induced Neuroprotection. Int. J. Mol. Sci. 2021, 22, 6479. [Google Scholar] [CrossRef] [PubMed]
- Pons, V.; Riera, J.; Capó, X.; Martorell, M.; Sureda, A.; Tur, J.A.; Drobnic, F.; Pons, A. Calorie Restriction Regime Enhances Physical Performance of Trained Athletes. J. Int. Soc. Sports Nutr. 2018, 15, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duivenvoorde, L.P.M.; van Schothorst, E.M.; Bunschoten, A.; Keijer, J. Dietary Restriction of Mice on a High-Fat Diet Induces Substrate Efficiency and Improves Metabolic Health. J. Mol. Endocrinol. 2011, 47, 81–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, J.; Ojcius, D.M.; Ko, Y.F.; Ke, P.Y.; Wu, C.Y.; Peng, H.H.; Young, J.D. Hormetic Effects of Phytochemicals on Health and Longevity. Trends Endocrinol. Metab. 2019, 30, 335–346. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.M.; Cordero, M.D.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Afrin, S.; Santos-Buelga, C.; González-Paramás, A.M.; Astolfi, P.; Rubini, C.; et al. Strawberry Consumption Improves Aging-Associated Impairments, Mitochondrial Biogenesis and Functionality through the AMP-Activated Protein Kinase Signaling Cascade. Food Chem. 2017, 234, 464–471. [Google Scholar] [CrossRef]
- Heinonen, S.; Jokinen, R.; Rissanen, A.; Pietiläinen, K.H. White Adipose Tissue Mitochondrial Metabolism in Health and in Obesity. Obes. Rev. 2020, 21, 1–23. [Google Scholar] [CrossRef]
- Sanayei, M.; Hajizadeh-Sharafabad, F.; Amirsasan, R.; Barzegar, A. High Intensity Interval Training with or without Chlorella Vulgaris Supplementation in Obese and Overweight Women: Effects on Mitochondrial Biogenesis, Performance, and Body Composition. Br. J. Nutr. 2021, 1–11. [Google Scholar] [CrossRef]
- Tyrka, A.R.; Parade, S.H.; Price, L.H.; Kao, H.-T.; Porton, B.; Philip, N.S.; Welch, E.S.; Carpenter, L.L. Alterations of Mitochondrial DNA Copy Number and Telomere Length with Early Adversity and Psychopathology. Biol. Psychiatry 2016, 79, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Lilja, S.; Bäck, H.; Stoll, C.; Mayer, A.; Pointner, A.; Hippe, B.; Krammer, U.; Haslberger, A.G. Increased Sirtuin Expression, Senescence Regulating MiRNAs, MtDNA, and Bifidobacteria Correlate with Wellbeing and Skin Appearance after Sirtuin-Activating Drink. Bioact. Compd. Health Dis. 2021, 4, 45–62. [Google Scholar] [CrossRef]
- Barrès, R.; Yan, J.; Egan, B.; Treebak, J.T.; Rasmussen, M.; Fritz, T.; Caidahl, K.; Krook, A.; O’Gorman, D.J.; Zierath, J.R. Acute Exercise Remodels Promoter Methylation in Human Skeletal Muscle. Cell Metab. 2012, 15, 405–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voisin, S.; Eynon, N.; Yan, X.; Bishop, D.J. Exercise Training and DNA Methylation in Humans. Acta Physiol. 2015, 213, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Radom-Aizik, S.; Zaldivar, F.; Oliver, S.; Galassetti, P.; Cooper, D.M. Evidence for MicroRNA Involvement in Exercise-Associated Neutrophil Gene Expression Changes. J. Appl. Physiol. 2010, 109, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, J.; Koulmann, N.; Banzet, S. Circulating MyomiRs: A New Class of Biomarkers to Monitor Skeletal Muscle in Physiology and Medicine. J. Cachexia. Sarcopenia Muscle 2018, 9, 20–27. [Google Scholar] [CrossRef]
- Scott, W.; Stevens, J.; Binder-Macleod, S.A. Human Skeletal Muscle Fiber Type Classifications. Phys. Ther. 2001, 81, 1810–1816. [Google Scholar] [CrossRef]
- Xie, L.L.; Shi, F.; Tan, Z.; Li, Y.; Bode, A.M.; Cao, Y. Mitochondrial Network Structure Homeostasis and Cell Death. Cancer Sci. 2018, 109, 3686–3694. [Google Scholar] [CrossRef]
- Youle, R.J.; Van Der Bliek, A.M. Mitochondrial Fission, Fusion and Stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef] [Green Version]
- Bishop, D.J.; Botella, J.; Genders, A.J.; Lee, M.J.C.; Saner, N.J.; Kuang, J.; Yan, X.; Granata, C. High-Intensity Exercise and Mitochondrial Biogenesis: Current Controversies and Future Research Directions. Physiology 2019, 34, 56–70. [Google Scholar] [CrossRef]
- Liu, C.; Lin, J.D. PGC-1 Coactivators in the Control of Energy Metabolism. Acta Biochim. Biophys. Sin. 2011, 43, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Kitada, M.; Ogura, Y.; Monno, I.; Koya, D. The Impact of Dietary Protein Intake on Longevity and Metabolic Health. EBioMedicine 2019, 43, 632–640. [Google Scholar] [CrossRef] [Green Version]
- Micha, R.; Michas, G.; Mozaffarian, D. Unprocessed Red and Processed Meats and Risk of Coronary Artery Disease and Type 2 Diabetes-An Updated Review of the Evidence. Curr. Atheroscler. Rep. 2012, 14, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellavia, A.; Stilling, F.; Wolk, A. High Red Meat Intake and All-Cause Cardiovascular and Cancer Mortality: Is the Risk Modified by Fruit and Vegetable Intake? Am. J. Clin. Nutr. 2016, 104, 1137–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valerio, A.; D’Antona, G.; Nisoli, E. Branched-Chain Amino Acids, Mitochondrial Biogenesis, and Healthspan: An Evolutionary Perspective. Aging 2011, 3, 464–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolk, A. Potential Health Hazards of Eating Red Meat. J. Intern. Med. 2017, 281, 106–122. [Google Scholar] [CrossRef]
- Hirabara, S.M.; Curi, R.; Maechler, P. Saturated Fatty Acid-Induced Insulin Resistance Is Associated with Mitochondrial Dysfunction in Skeletal Muscle Cells. J. Cell. Physiol. 2010, 222, 187–194. [Google Scholar] [CrossRef]
- Niedzwiecki, A.; Roomi, M.W.; Kalinovsky, T.; Rath, M. Anticancer Efficacy of Polyphenols and Their Combinations. Nutrients 2016, 8, 552. [Google Scholar] [CrossRef] [Green Version]
- Mahn, A.; Reyes, A. An Overview of Health-Promoting Compounds of Broccoli (Brassica Oleracea Var. Italica) and the Effect of Processing. Food Sci. Technol. Int. 2012, 18, 503–514. [Google Scholar] [CrossRef]
- D’Angelo, S. Polyphenols: Potential Beneficial Effects of These Phytochemicals in Athletes. Curr. Sports Med. Rep. 2020, 19, 260–265. [Google Scholar] [CrossRef]
- Eluamai, A.; Brooks, K. Effect of Aerobic Exercise on Mitochondrial DNA and Aging. J. Exerc. Sci. Fit. 2013, 11, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Islam, H.; Hood, D.A.; Gurd, B.J. Looking beyond PGC-1α: Emerging Regulators of Exercise-Induced Skeletal Muscle Mitochondrial Biogenesis and Their Activation by Dietary Compounds. Appl. Physiol. Nutr. Metab. 2020, 45, 11–23. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Monday Training A | Tuesday Endurance | Wednesday Training B | Thursday Break | Friday Training A | Saturday Endurance | Sunday Break |
|---|---|---|---|---|---|---|
| Warm up 5–10 min. stepper | 30 to at least 60 min. in the last weeks of intervention | Warm up 5–10 min. stepper | - | Warm up 5–10 min. stepper | 30 to at least 60 min. in the last weeks of intervention | - |
| Working sets Squats: 3 × 8–12 rep. Gluteus 3 × 8–12 rep. Chest press: 3 × 8–12 rep. Dumbbell row: 3 × 8–12 rep. Plank 3 × 8–12 rep. | Working sets Leg press: 3 × 8–12 rep. Lunges: 3 × 8–12 rep. Push-ups: 3 × 8–12 rep. Inverted row: 3 × 8–12 rep. Abdominal crunches 3 × 8–12 rep. | Working sets Squats: 3 × 8–12 rep. Gluteus 3 × 8–12 rep. Chest press: 3 × 8–12 rep. Dumbbell row: 3 × 8–12 rep. Plank 3 × 8–12 rep. | ||||
| Cool down 10 min stretching | Cool down 10 min stretching | Cool down 10 min stretching |
| Assay ID | Primer | Sequence | No. of CpG Sites |
|---|---|---|---|
| PGC1A | fw: | 5′-TAT AGT TAT TTT GTT ATG AAA TAG GGA GTT TTG -3′ | 1 |
| rev: | 5′- biotin-CCA ATC ACA TAA CAA AAC TAT TAA AAA ATA A -3′ | ||
| seq: | 5′-GGA TTT TGG TTA TTA TAT GGT TAG G -3′ | ||
| Sequence to analyze: | GTT TYG TTT AGA GTT TG |
| Total (n = 36) | Female (n = 21) | Male (n = 15) | |
|---|---|---|---|
| Age ± SD [years] | 31.86 ± 8.02 | 30.14 ± 8.51 | 34.27 ± 6.82 |
| BMI ± SD [T0, kg/m2] | 24.17 ± 2.95 | 23.78 ± 3.18 | 24.71 ± 2.61 |
| BMI ± SD [T1, kg/m2] | 23.88 ± 2.71 | 23.51 ± 2.95 | 24.40 ± 2.33 |
| BMI classes | |||
| Normal weight 18.5–24.9 kg/m2 | 58.3% | 61.9% | 53.3% |
| Overweight, 25–29.9 kg/m2 | 38.9% | 33.3% | 46.7% |
| Obesity grad I, 30–34.9 kg/m2 | 2.8% | 4.8% | - |
| Basal metabolic rate ± SD [T0, kcal] | 1481.11 ± 178.41 | 1355.71 ± 73.46 | 1656.67 ± 123.56 |
| Basal metabolic rate ± SD [T1, kcal] | 1495.81 ± 175.74 | 1369.05 ± 61.72 | 1673.27 ± 118.03 |
| LBM ± SD [T0, kg] | 51.93 ± 9.74 | 45.02 ± 4.96 | 61.60 ± 5.47 |
| LBM ± SD [T1, kg] | 52.44 ± 9.87 | 45.47 ± 4.72 | 62.20 ± 6.07 |
| BFM ± SD [T0, kg] | 20.06 ± 5.67 | 21.21 ± 6.08 | 18.44 ± 4.77 |
| BFM ± SD [T1, kg] | 18.53 ± 5.62 | 19.99 ± 5.84 | 16.47 ± 4.75 |
| BFP ± SD [T0, %] | 27.75 ± 6.21 | 31.24 ± 4.88 | 22.86 ± 4.31 |
| BFP ± SD [T1, %] | 26.04 ± 6.77 | 29.76 ± 5.27 | 20.83 ± 5.02 |
| Intake frequencies | |||
| Red/processed meat | |||
| Rarely or never | 44.4% | 57.2% | 26.6% |
| Once/week | 25.0% | 23.8% | 26.6% |
| 2–3×/week | 22.2% | 9.5% | 40.0% |
| >4×/week or daily | 8.4% | 9.5% | 6.8% |
| Cruciferous vegetables | |||
| Rarely or never | 58.3% | 57.1% | 60.0% |
| ≥once/week | 41.7% | 42.9% | 40.0% |
| Marker | Total (n = 36) 24–48 h Post-Exercise | Reduced Group (n = 27) 24–32 h Post-Exercise | ||
|---|---|---|---|---|
| Fold Change ± SD | p Value | Fold Change ± SD | p Value | |
| PGC-1α Methylation | 1.49 ± 1.42 | 0.826 | 1.88 ± 1.45 | 0.024 * |
| mtDNA | 1.09 ± 0.37 | 0.426 | 1.10 ± 0.39 | 0.583 |
| miR-23a-3p | 0.92 ± 0.35 | 0.023 * | 0.91 ± 0.36 | 0.028 * |
| miR-30e-3p | 0.94 ± 0.34 | 0.047 * | 0.94 ± 0.37 | 0.088 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krammer, U.D.B.; Sommer, A.; Tschida, S.; Mayer, A.; Lilja, S.V.; Switzeny, O.J.; Hippe, B.; Rust, P.; Haslberger, A.G. PGC-1α Methylation, miR-23a, and miR-30e Expression as Biomarkers for Exercise- and Diet-Induced Mitochondrial Biogenesis in Capillary Blood from Healthy Individuals: A Single-Arm Intervention. Sports 2022, 10, 73. https://doi.org/10.3390/sports10050073
Krammer UDB, Sommer A, Tschida S, Mayer A, Lilja SV, Switzeny OJ, Hippe B, Rust P, Haslberger AG. PGC-1α Methylation, miR-23a, and miR-30e Expression as Biomarkers for Exercise- and Diet-Induced Mitochondrial Biogenesis in Capillary Blood from Healthy Individuals: A Single-Arm Intervention. Sports. 2022; 10(5):73. https://doi.org/10.3390/sports10050073
Chicago/Turabian StyleKrammer, Ulrike D. B., Alexandra Sommer, Sylvia Tschida, Anna Mayer, Stephanie V. Lilja, Olivier J. Switzeny, Berit Hippe, Petra Rust, and Alexander G. Haslberger. 2022. "PGC-1α Methylation, miR-23a, and miR-30e Expression as Biomarkers for Exercise- and Diet-Induced Mitochondrial Biogenesis in Capillary Blood from Healthy Individuals: A Single-Arm Intervention" Sports 10, no. 5: 73. https://doi.org/10.3390/sports10050073
APA StyleKrammer, U. D. B., Sommer, A., Tschida, S., Mayer, A., Lilja, S. V., Switzeny, O. J., Hippe, B., Rust, P., & Haslberger, A. G. (2022). PGC-1α Methylation, miR-23a, and miR-30e Expression as Biomarkers for Exercise- and Diet-Induced Mitochondrial Biogenesis in Capillary Blood from Healthy Individuals: A Single-Arm Intervention. Sports, 10(5), 73. https://doi.org/10.3390/sports10050073