
A Novel Approach to the Determination of Time- and Fatigue-Dependent Efficiency during Maximal Cycling Sprints

Abstract
:1. Introduction
2. Methods
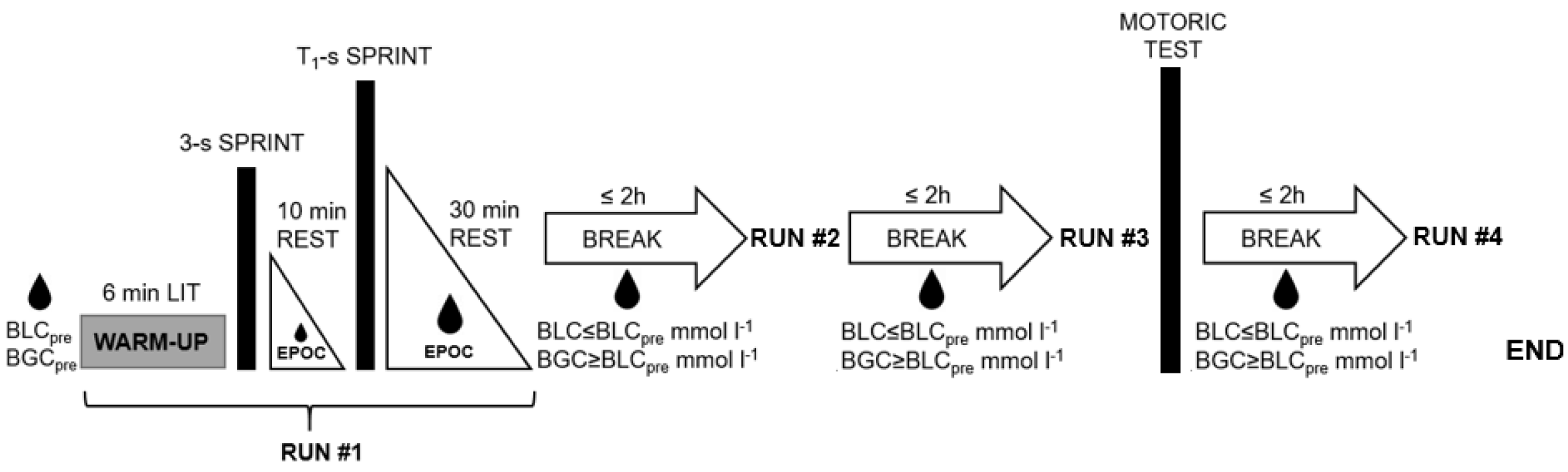
2.1. The Exercise Protocol
2.2. Data Processing
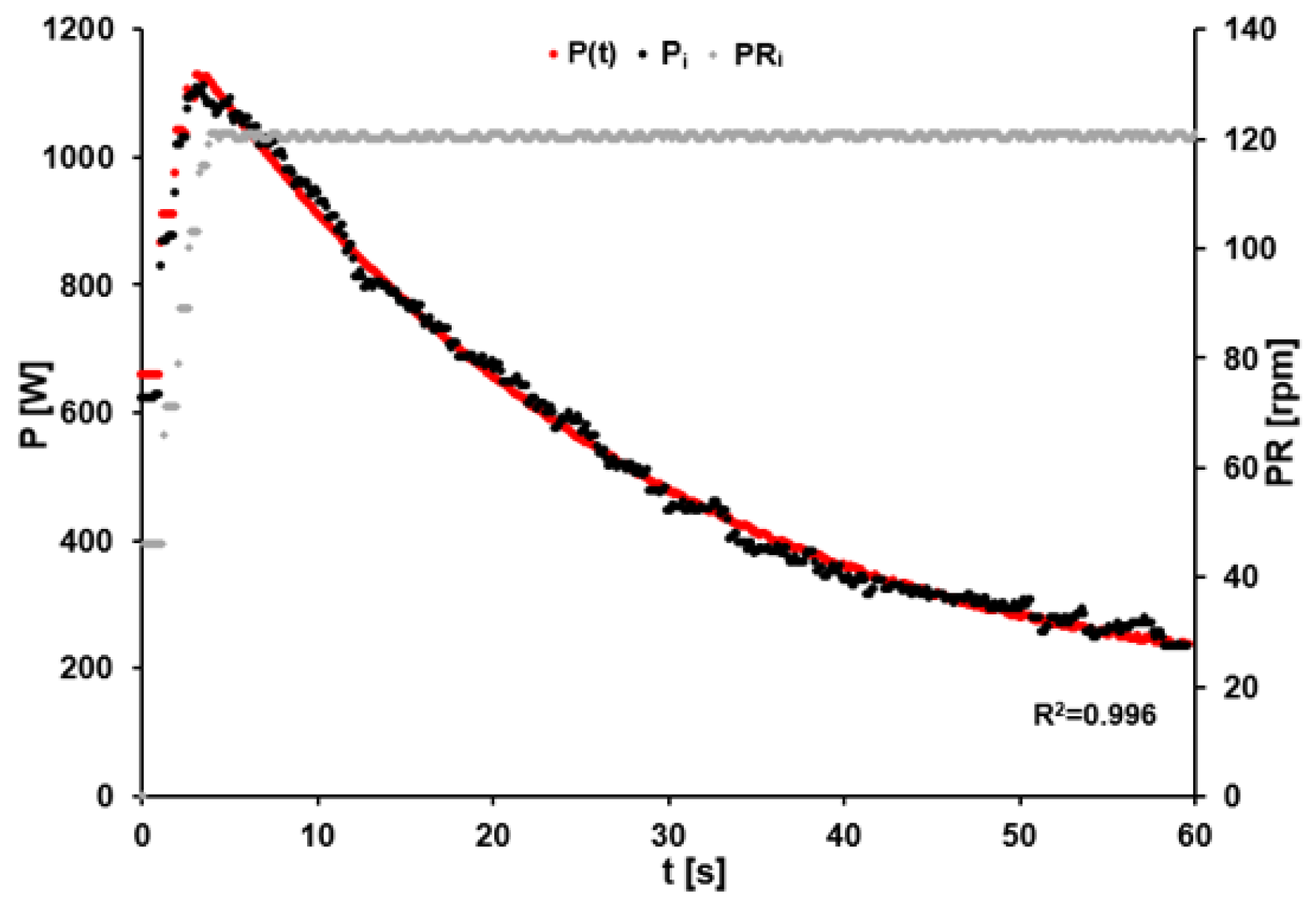
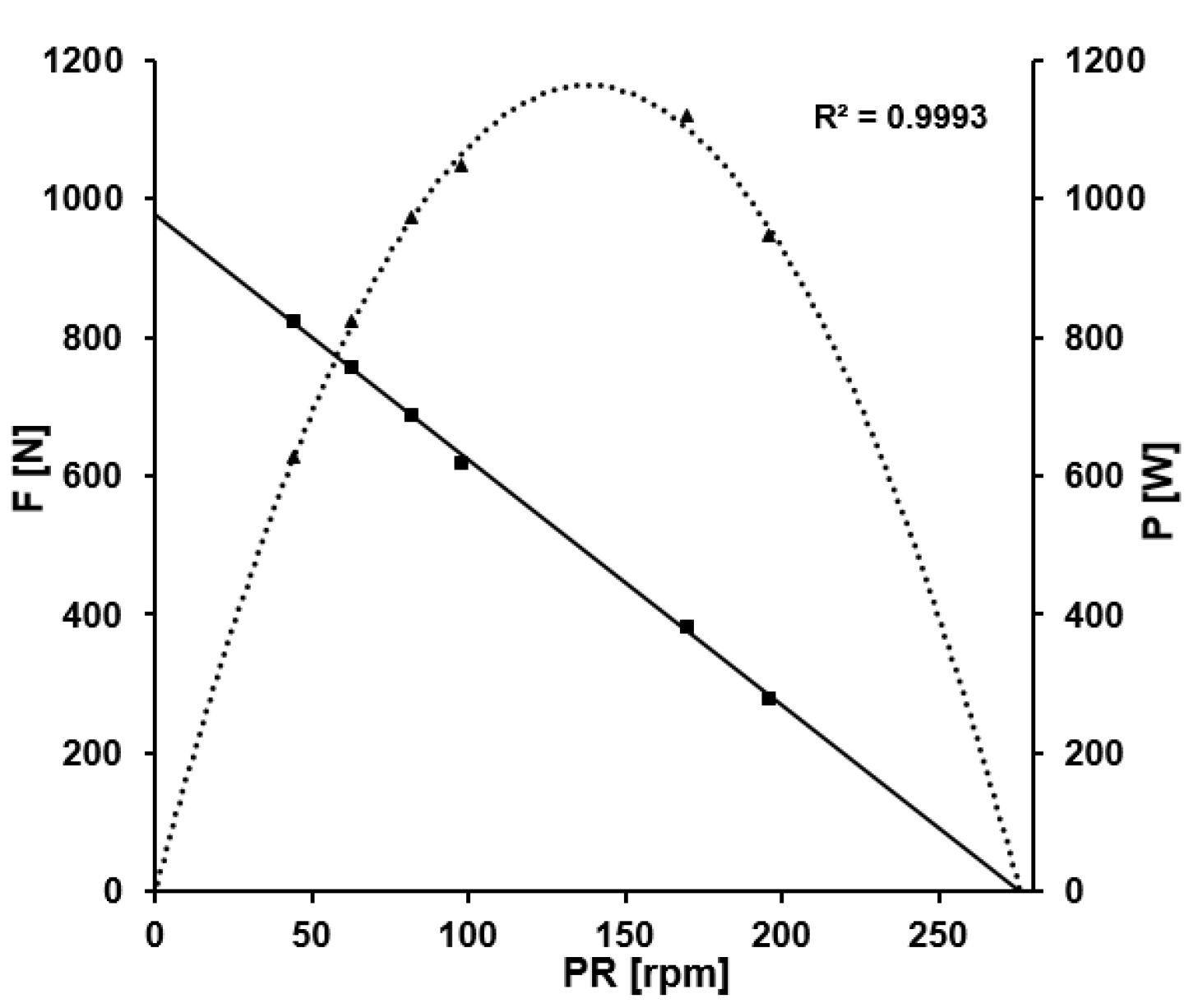
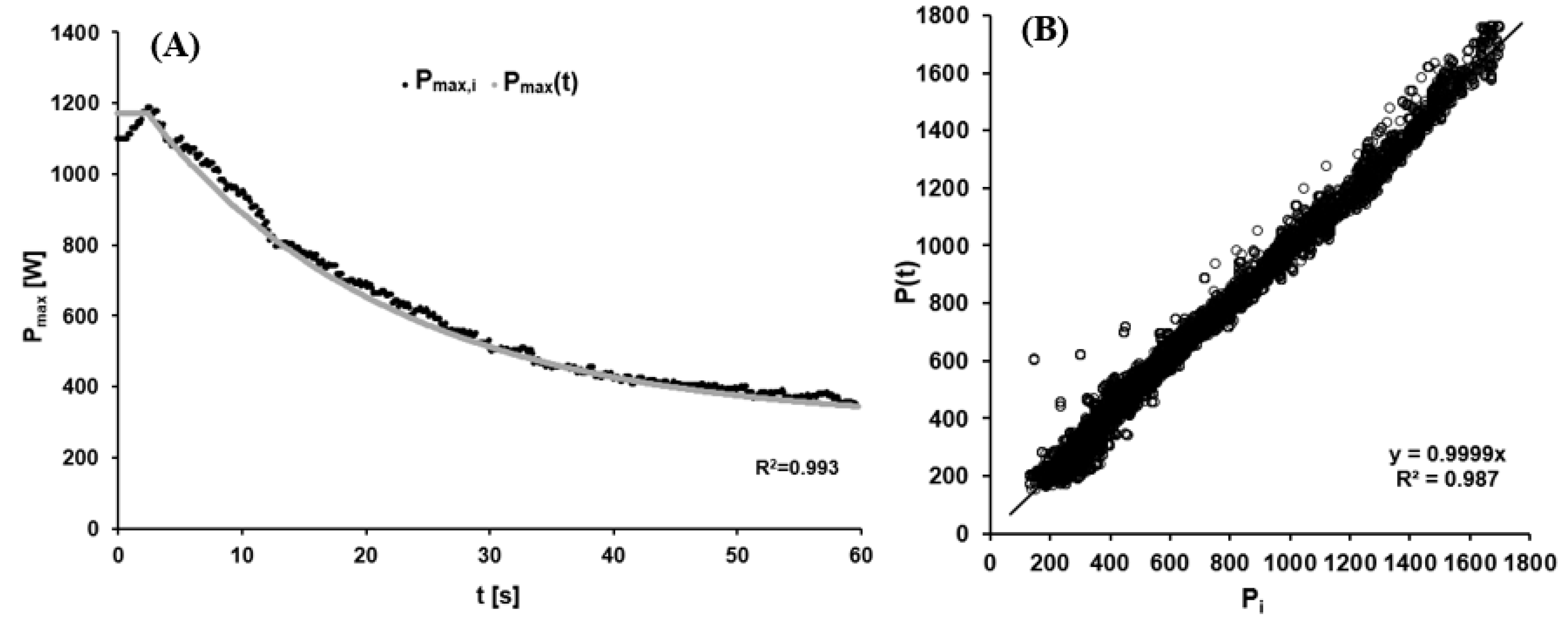
2.3. External Power
2.4. Metabolic Power
2.5. Statistical Analyses
3. Results
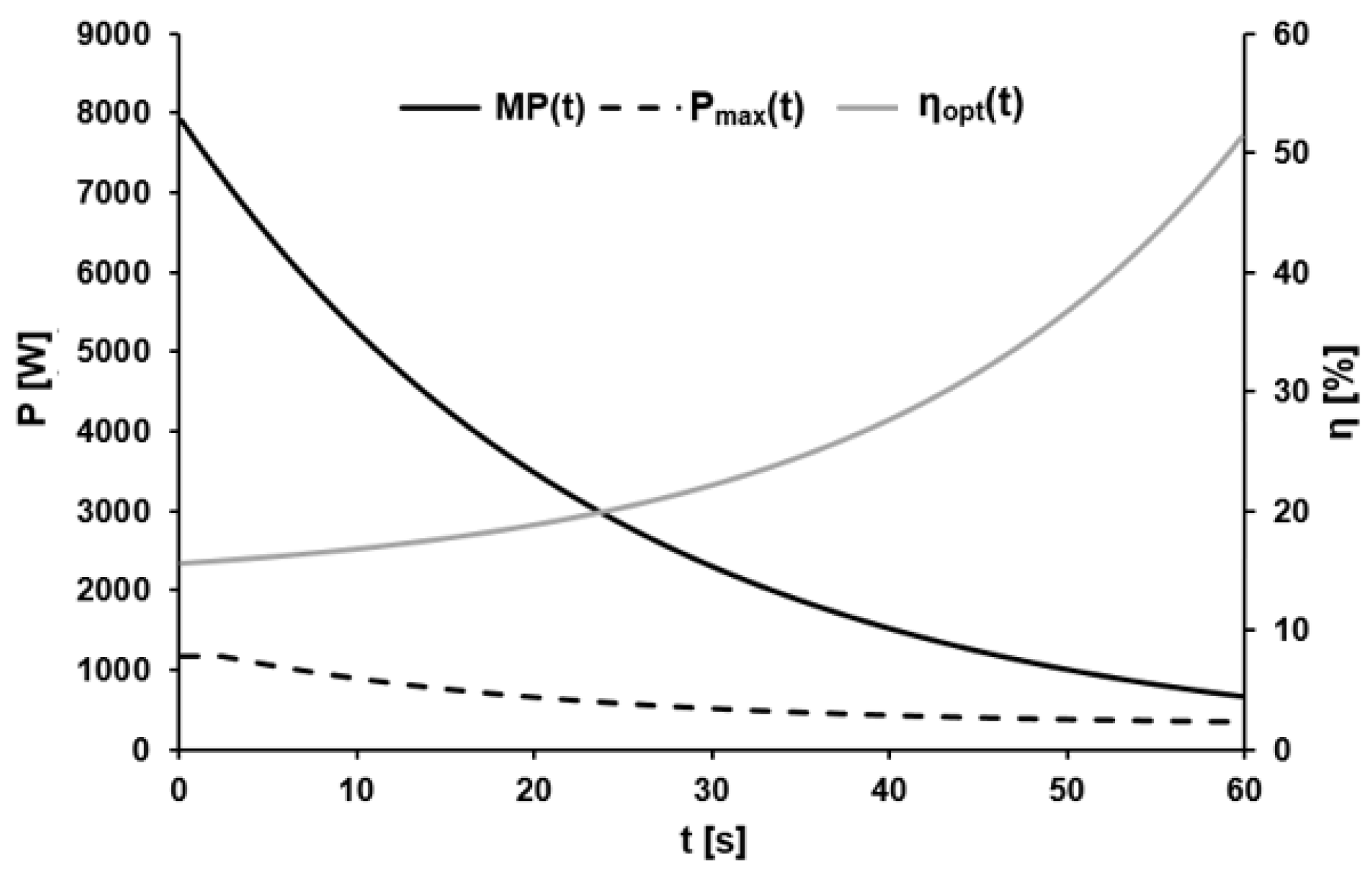
3.1. External Power
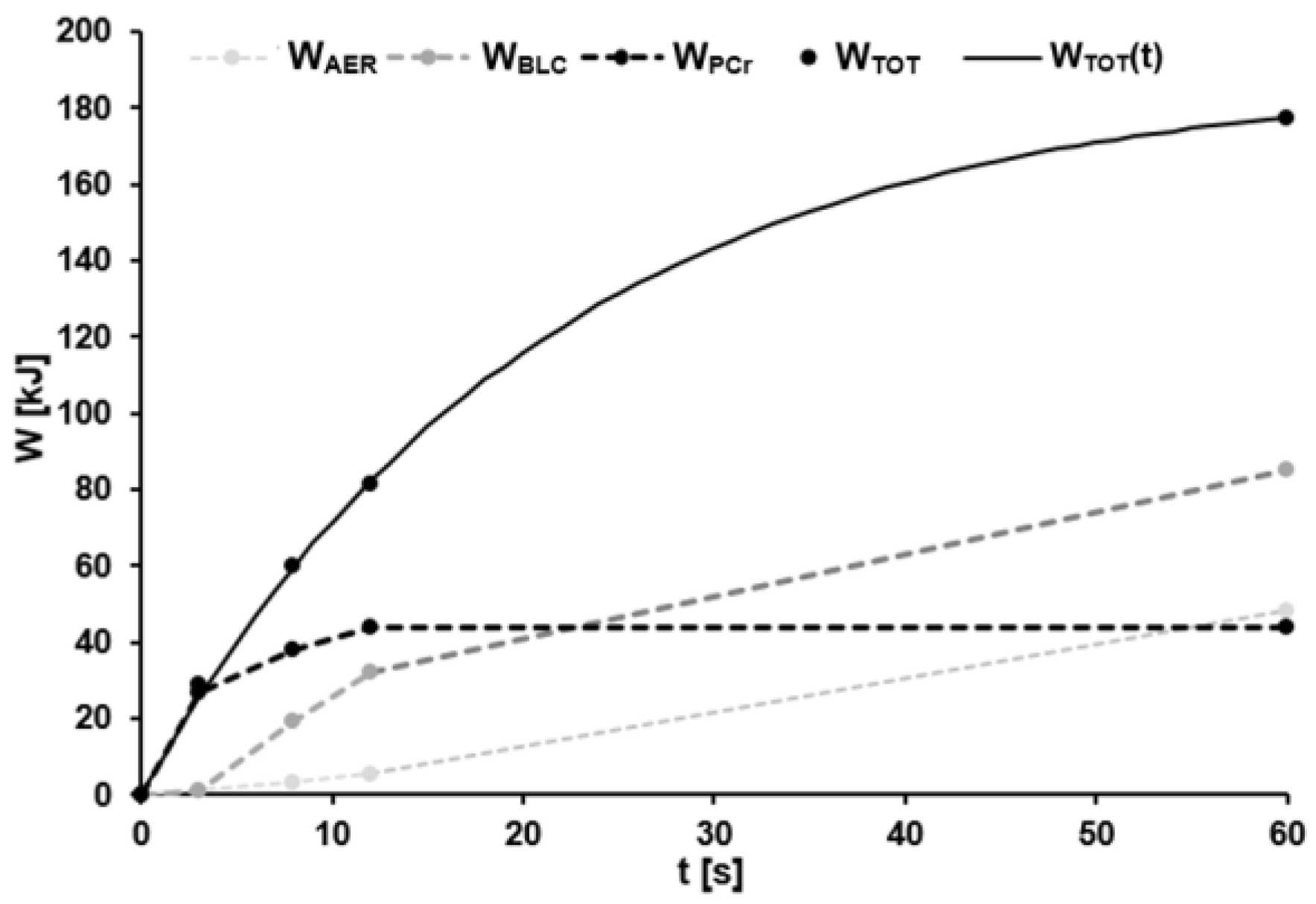
3.2. Metabolic Power
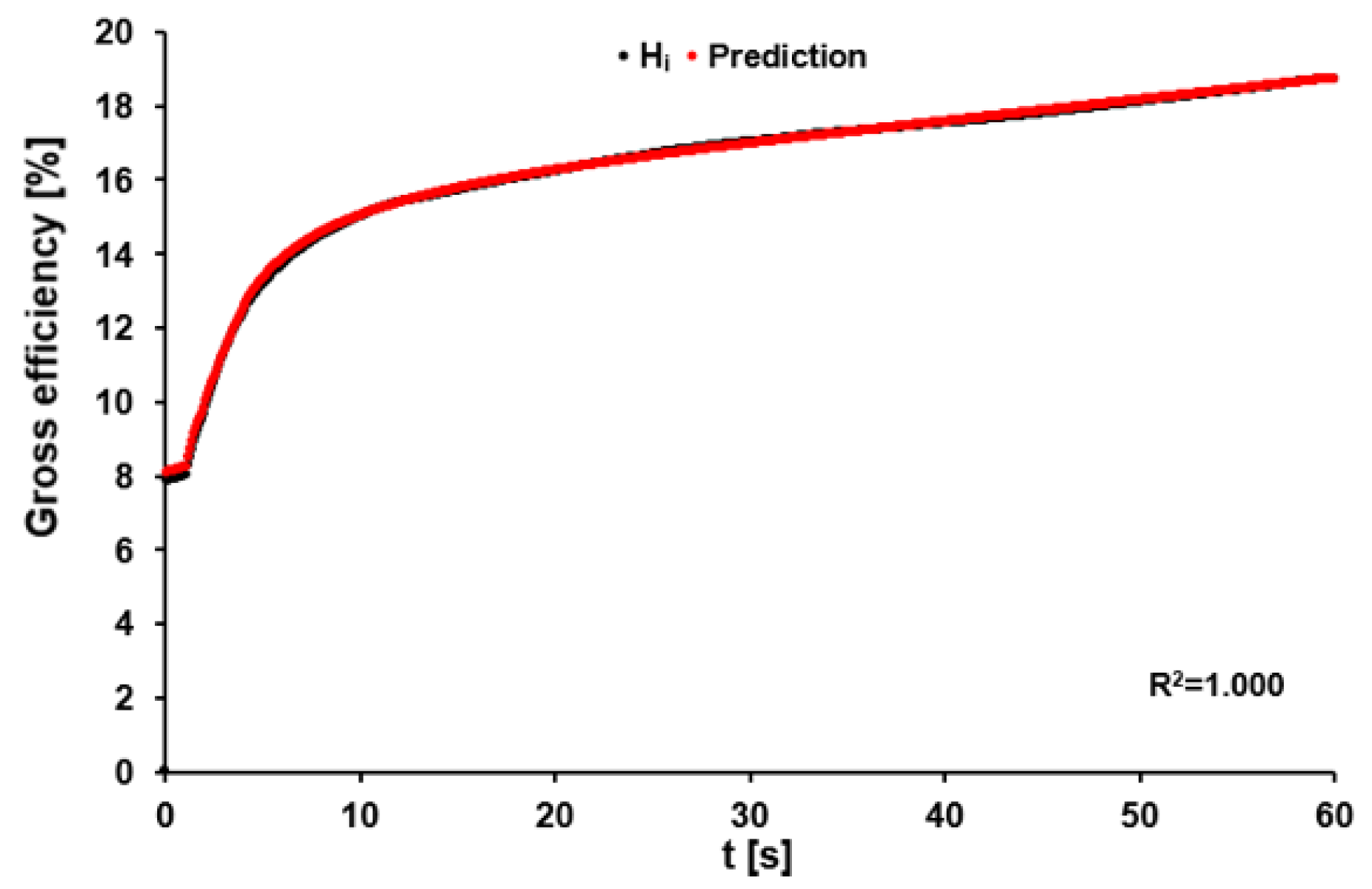
3.3. Optimal Efficiency and Recalculation of Gross Efficiency
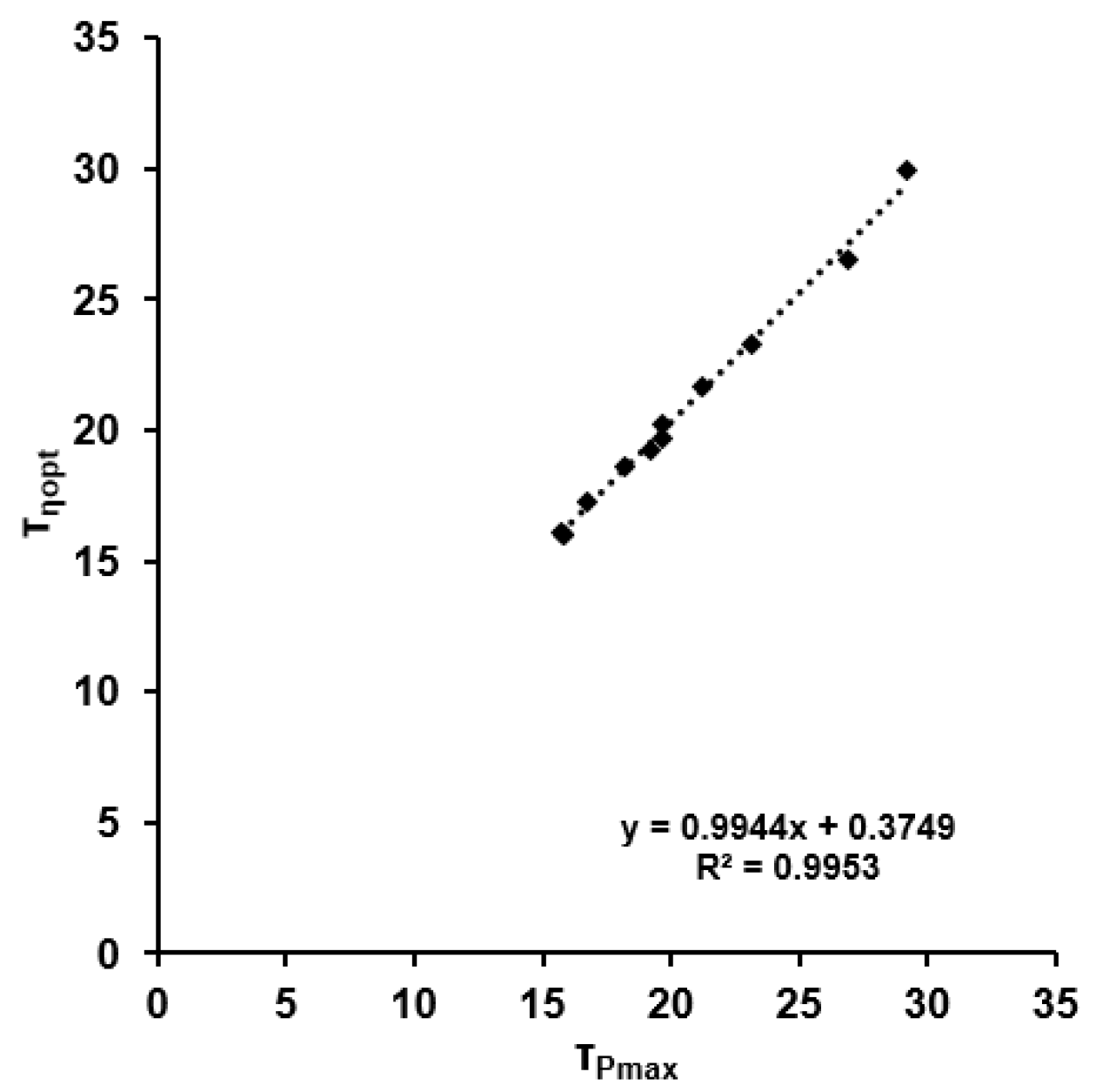
3.4. Correlation Analyses
4. Discussion
4.1. Metabolic Power
4.2. External Power
4.3. Optimal Efficiency
5. Practical Applications
6. Limitations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| a | Slope of the fatigue-free F/v profile |
| A | Amplitude of the extra-vascular lactate concentration |
| AFC | Amplitude of oxygen consumption during the fast component of EPOC |
| AMP | Amplitude of the metabolic power |
| AP | Amplitude of the maximal dynamic power output |
| ASC | Amplitude of oxygen consumption during the slow component of EPOC |
| Aw | Amplitude of the metabolic work |
| b, bi | the intercept of the F/v profile with the y-axis for data point i |
| BGC | Blood glucose concentration |
| BLC | Blood lactate concentration |
| Maximal blood lactate concentration | |
| BLC(t) | Post-exercise blood lactate concentration |
| BLC(0) | Pre-exercise blood lactate concentration |
| BM | Body mass |
| CE | Caloric equivalent |
| CP | The limiting value of maximal dynamic power output |
| EPOC | Excess Post-Exercise Oxygen Consumption |
| F | Mean pedal force |
| F/v | Force/velocity |
| F(v) | Mean pedal force as a function of pedaling rate |
| F(PRi) | Mean pedal force in connection with cadence i |
| H | The amplitude of optimal efficiency |
| Hi | Gross efficiency associated with data point i |
| H0 | The baseline amplitude of optimal efficiency |
| The ith segment of the total time subdivided into n segments | |
| k1 | Rate constants for the appearance of BLC |
| k2 | Rate constants for the disappearance of BLC |
| LAO2E | –lactate equivalent |
| MHC | Myosin heavy chain |
| MP | Metabolic power |
| P | Output of external/mechanical propulsive power |
| P/v | Power/velocity |
| P(v) | Mechanical power output as a function of pedalling rate |
| PCr | Phosphocreatine |
| Pmax, Pmax,i | Maximal power output derived from P/v profile, maximal power output for data point i |
| Pmax(t) | Maximal mechanical power output as a function of time, i.e., the dynamic maximal power output |
| PR, PRi | Pedaling rate, cadence associated with data point i |
| PRmax | Maximal pedaling rate, maximal cadence |
| RER | Respiratory exchange rate |
| rpm | Revolutions of the crank per minute, cadence |
| T | Exercise duration |
| TD | Time delay |
| v | Velocity of the crank or pedal |
| Oxygen uptake | |
| VO2 | Oxygen consumption during exerise |
| Baseline of oxygen uptake at rest | |
| Excess Post-Exercise Oxygen Consumption | |
| Oxygen consumption required to replenish PCr post-exercise; oxygen consumption during the fast component of EPOC | |
| VO2SC | Oxygen consumption during the slow component of EPOC |
| WAER | Aerobic energy supply |
| WBLC | Lactic energy supply |
| WP, WP,i | External work, external work for data point i |
| WPCr | Phosphagenic energy supply, alactic energy supply |
| WTOT, WTOT,i | Total energy supply, total energy expended during pedal stroke i |
| ΔBLC | Blood lactate accumulation |
| η | Efficiency |
| ηopt,i | Optimal efficiency for data point i |
| ηopt | Optimal efficiency |
| τFC | Time constant of the fast component of EPOC(t) |
| τSC | Time constant of the slow component of EPOC(t) |
| τP | Time constant of maximal external power output |
| τMP | Time constant of metabolic power |
| τW | Time constant of metabolic work |
| τη | Time constant of optimal efficiency |
| In | |
| For all |
References
- Martin, J.C.; Wagner, B.M.; Coyle, E.F. Inertial-load method determines maximal cycling power in a single exercise bout. Med. Sci. Sports Exerc. 1997, 29, 1505–1512. [Google Scholar] [CrossRef]
- Dorel, S.; Hautier, C.A.; Rambaud, O.; Rouffet, D.; Van Praagh, E.; Lacour, J.-R.; Bourdin, M. Torque and Power-Velocity Relationships in Cycling: Relevance to Track Sprint Performance in World-Class Cyclists. Int. J. Sports Med. 2005, 26, 739–746. [Google Scholar] [CrossRef]
- Gardner, A.S.; Martin, J.C.; Martin, D.T.; Barras, M.; Jenkins, D.G. Maximal torque- and power-pedaling rate relationships for elite sprint cyclists in laboratory and field tests. Eur. J. Appl. Physiol. 2007, 101, 287–292. [Google Scholar] [CrossRef]
- Abbiss, C.R.; Peiffer, J.J.; Laursen, P. Optimal cadance selection during cycling. Int. J. Sports Med. 2009, 10, 1–15. [Google Scholar]
- Mader, A.; Heck, H. Energiestoffwechselregulation, Erweiterung des theoretischen Konzepts und seiner Begründungen. Nachweis der praktischen Nützlichkeit der Simulation des Energiestoffwechsels. BSW 1994, 8, 124–162. [Google Scholar]
- Hansen, E.A.; Sjøgaard, G. Relationship between efficiency and pedal rate in cycling: Significance of internal power and muscle fiber type composition. Scand. J. Med. Sci. Sports 2006, 17, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Böning, D.; Maassen, N.; Steinach, M. The efficiency of muscular exercise. Dtsch Z Sportmed. 2017, 68, 203–214. [Google Scholar] [CrossRef]
- Di Prampero, P.E. Energetics of muscular exercise. Rev. Physiol. Biochem. Pharmacol. 1981, 89, 143–222. [Google Scholar] [CrossRef]
- MacDougall, K.B.; Falconer, T.M.; MacIntosh, B.R. Efficiency of cycling exercise: Quantification, mechanisms, and misunderstandings. Scand. J. Med. Sci. Sports 2022, 32, 951–970. [Google Scholar] [CrossRef]
- Noordhof, D.A.; Mulder, R.C.; Malterer, K.R.; Foster, C.; de Koning, J.J. The Decline in Gross Efficiency in Relation to Cycling Time-Trial Length. Int. J. Sports Physiol. Perform. 2015, 10, 64–70. [Google Scholar] [CrossRef]
- Gastin, P.B. Energy System Interaction and Relative Contribution During Maximal Exercise. Sports Med. 2001, 31, 725–741. [Google Scholar] [CrossRef] [PubMed]
- Dunst, A.K.; Manunzio, C.; Feldmann, A.; Hesse, C. Applications of near-infrared spectroscopy in “anaerobic” diagnostics—SmO2 kinetics reflect PCr dephosphorylation and correlate with maximal lactate accumulation and maximal pedalling rate. Biol. Sport 2023, Accepted. [Google Scholar]
- Sargeant, A.J. Structural and functional determinants of human muscle power. Exp. Physiol. 2007, 92, 323–331. [Google Scholar] [CrossRef] [PubMed]
- He, Z.-H.; Bottinelli, R.; Pellegrino, M.A.; Ferenczi, M.A.; Reggiani, C. ATP Consumption and Efficiency of Human Single Muscle Fibers with Different Myosin Isoform Composition. Biophys. J. 2000, 79, 945–961. [Google Scholar] [CrossRef] [Green Version]
- Kohler, G.; Boutellier, U. The generalized force–velocity relationship explains why the preferred pedaling rate of cyclists exceedsthe most efficient one. Eur. J. Appl. Physiol. 2005, 94, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Coyle, E.F.; Sidossis, L.S.; Horowitz, J.F.; Beltz, J.D. Cycling efficiency is related to the percentage of type I muscle fibers. Med Sci Sports Exerc. 1992, 24, 782–788. [Google Scholar] [CrossRef]
- Moseley, L.; Jeukendrup, A.E. The reliability of cycling efficiency. Med. Sci. Sports Exerc. 2001, 33, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, M.; Bagger, M.; Pedersen, P.K.; Fernström, M.; Sahlin, K. Cycling efficiency in humans is related to low UCP3 content and to type I fibers but not to mitochondrial efficiency. J. Physiol. 2006, 571, 669–681. [Google Scholar] [CrossRef]
- Beneke, R.; Pollmann, C.; Bleif, I.; Leithäuser, R.M.; Hütler, M. How anaerobic is the Wingate Anaerobic Test for humans? Eur. J. Appl. Physiol. 2002, 87, 388–392. [Google Scholar]
- Henneman, E.; Mendell, L.M. Functional organization of motoneurone pool and its inputs. In Handbook of Physiology, Section 1, The Nervous System, Volume II, Motor Control; Brookhart, J.M., Mountcastle, V.B., Brooks, V.B., Geiger, S.R., Eds.; American Physiological Society: Rockville, MD, USA, 1981. [Google Scholar]
- Beltman, J.G.; Haan, H.; Gerrits, H.L.; van Mechelen, W.; Sargeant, A.J.; de Haan, A. Metabolically assessed recruitment of single muscle fibers during 7 voluntary isometric contractions. Med. Sci. Sports Exerc. 2003, 35, S95. [Google Scholar] [CrossRef]
- Dunst, A.K.; Grüneberger, R.; Holmberg, H.-C. Modeling Optimal Cadence as a Function of Time during Maximal Sprint Exercises Can Improve Performance by Elite Track Cyclists. Appl. Sci. 2021, 11, 12105. [Google Scholar] [CrossRef]
- Gaesser, G.A.; Brooks, G.A. Muscular efficiency during steady-rate exercise: Effects of speed and work rate. J. Appl. Physiol. 1975, 38, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y. Mechanical efficiency of fast- and slow-twitch muscle fibers in man during cycling. J. Appl. Physiol. 1979, 47, 263–267. [Google Scholar] [CrossRef]
- Böning, D.; Gönen, Y.; Maassen, N. Relationship Between Work Load, Pedal Frequency, and Physical Fitness. Int. J. Sports Med. 1984, 5, 92–97. [Google Scholar] [CrossRef]
- Sidossis, L.S.; Horowitz, J.F.; Coyle, E.F. Load and Velocity of Contraction Influence Gross and Delta Mechanical Efficiency. Int. J. Sports Med. 1992, 13, 407–411. [Google Scholar] [CrossRef]
- Chavarren, J.; Calbet, J.A.L. Cycling efficiency and pedalling frequency in road cyclists. Eur. J. Appl. Physiol. 1999, 80, 555–563. [Google Scholar] [CrossRef]
- Dunst, A.K.; Hesse, C.; Ueberschär, O. The Concept of Optimal Dynamic Pedalling Rate and Its Application to Power Output and Fatigue in Track Cycling Sprinters—A Case Study. Sports 2023, 11, 19. [Google Scholar] [CrossRef]
- Beneke, R.; Böning, D. The limits of human performance. Essays Biochem. 2008, 44, 11–25. [Google Scholar] [CrossRef]
- Dunst, A.K.; Hesse, C.; Feldmann, A.; Holmberg, H.C. A novel approach to determine alactic timespan in the assessment of the maximal lactate accumulation rate in elite track cyclists. Int. J. Sport. Physiol. Perform. 2023, 1, 1–7. [Google Scholar] [CrossRef]
- Dunst, A.K.; Hesse, C.; Ueberschär, O.; Holmberg, H.-C. Fatigue-Free Force-Velocity and Power-Velocity Profiles for Elite Track Sprint Cyclists: The Influence of Duration, Gear Ratio and Pedalling Rates. Sports 2022, 10, 130. [Google Scholar] [CrossRef]
- McCartney, N.; Heigenhauser, G.J.; Jones, N.L. Power output and fatigue of human muscle in maximal cycling exercise. J. Appl. Physiol. 1983, 55, 218–224. [Google Scholar] [CrossRef]
- Seow, C.Y. Hill’s equation of muscle performance and its hidden insight on molecular mechanisms. J. Gen. Physiol. 2013, 142, 561–573. [Google Scholar] [CrossRef] [Green Version]
- Buttelli, O.; Seck, D.; Vandewalle, H.; Jouanin, J.C.; Monod, H. Effect of fatigue on maximal velocity and maximal torque during short exhausting cycling. Eur. J. Appl. Physiol. Occup. Physiol. 1996, 73, 175–179. [Google Scholar] [CrossRef]
- Knuttgen, H.G. Oxygen dept after submaximal exercise. J. Appl. Physiol. 1970, 29, 651–657. [Google Scholar] [CrossRef]
- Beneke, R.; Beyer, T.; Jachner, C.; Erasmus, J.; Hütler, M. Energetics of karate kumite. Eur. J. Appl. Physiol. 2004, 92, 518–523. [Google Scholar] [CrossRef]
- Mader, A.; Heck, H.; Hollman, W. Leistung und Leistungsbegrenzung des menschlichen Organismus interpretiert am Modell thermodynamischer offener Systeme. Ein Beitrag zur Diskussion biologischer Grenzen im Hochleistungssport. In Sport an der Grenze Menschlicher Leistungsfähigkeit; Mader, A., Heck, H., Hollman, W., Eds.; Springer: Berlin/Heidelberg, Germany, 1981; pp. 69–83. [Google Scholar]
- Beneke, R.; Jumah, M.D.; Leithäuser, R.M. Modelling the lactate response to short-term all out exercise. Dyn Med. 2007, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Ueberschär, O.; Fleckenstein, D.; Warschun, F.; Walter, N.; Wüstenfeld, J.C.; Wolfarth, B.; Hoppe, M.W. Energy Cost of Running Under Hypogravity in Well-Trained Runners and Triathletes: A Biomechanical Perspective. Int. J. Comput. Sci. Sport 2019, 18, 60–80. [Google Scholar] [CrossRef] [Green Version]
- Lovell, D.; Kerr, A.; Wiegand, A.; Solomon, C.; Harvey, L.; McLellan, C. The contribution of energy systems during the upper body Wingate anaerobic test. Appl. Physiol. Nutr. Metab. 2013, 38, 216–219. [Google Scholar] [CrossRef]
- Harvey, L.; Wiegand, A.; Solomon, C.; McLellan, C.; Lovell, D. A comparison of upper and lower body energetics during high-intensity exercise. . J. Sports Med. Phys. Fit. 2015, 55, 708–713. [Google Scholar]
- Leithäuser, R.M.; Böning, D.; Hütler, M.; Beneke, R. Enhancement on Wingate Anaerobic Test Performance With Hyperventilation. Int. J. Sports Physiol. Perform. 2016, 11, 627–634. [Google Scholar] [CrossRef]
- Julio, U.F.; Panissa, V.L.G.; Cury, R.L.; Agostinho, M.F.; Esteves, J.V.D.C.; Franchini, E. Energy System Contributions in Upper and Lower Body Wingate Tests in Highly Trained Athletes. Res. Q. Exerc. Sport 2019, 90, 244–250. [Google Scholar] [CrossRef] [PubMed]
- de Poli, R.A.B.; Miyagi, W.E.; Zagatto, A.M. Anaerobic Capacity is Associated with Metabolic Contribution and Mechanical Output Measured During the Wingate Test. J. Hum. Kinet. 2021, 79, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Ulupınar, S.; Özbay, S. Energy pathway contributions during 60-second upper-body Wingate test in Greco-Roman wrestlers: Intermittent versus single forms. Res. Sports Med. 2021, 30, 244–255. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | 3-s | 8-s | 12-s | 60-s |
|---|---|---|---|---|
| VO2FC [mL] | 1441 ± 349 | 2039 ± 462 | 2256 ± 473 | 2550 ± 572 |
| ΔBLC [mmol L−1] | 0.64 ± 0.24 | 4.58 ± 0.66 | 7.98 ± 0.89 | 17.69 ± 2.11 |
| VO2 [mL] | 75 ± 23 | 226 ± 57 | 386 ± 98 | 3220 ± 536 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dunst, A.K.; Hesse, C.; Ueberschär, O.; Holmberg, H.-C. A Novel Approach to the Determination of Time- and Fatigue-Dependent Efficiency during Maximal Cycling Sprints. Sports 2023, 11, 29. https://doi.org/10.3390/sports11020029
Dunst AK, Hesse C, Ueberschär O, Holmberg H-C. A Novel Approach to the Determination of Time- and Fatigue-Dependent Efficiency during Maximal Cycling Sprints. Sports. 2023; 11(2):29. https://doi.org/10.3390/sports11020029
Chicago/Turabian StyleDunst, Anna Katharina, Clemens Hesse, Olaf Ueberschär, and Hans-Christer Holmberg. 2023. "A Novel Approach to the Determination of Time- and Fatigue-Dependent Efficiency during Maximal Cycling Sprints" Sports 11, no. 2: 29. https://doi.org/10.3390/sports11020029
APA StyleDunst, A. K., Hesse, C., Ueberschär, O., & Holmberg, H. -C. (2023). A Novel Approach to the Determination of Time- and Fatigue-Dependent Efficiency during Maximal Cycling Sprints. Sports, 11(2), 29. https://doi.org/10.3390/sports11020029