Intermittent Dieting: Theoretical Considerations for the Athlete

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
3. Adaptive Responses to Energy Restriction
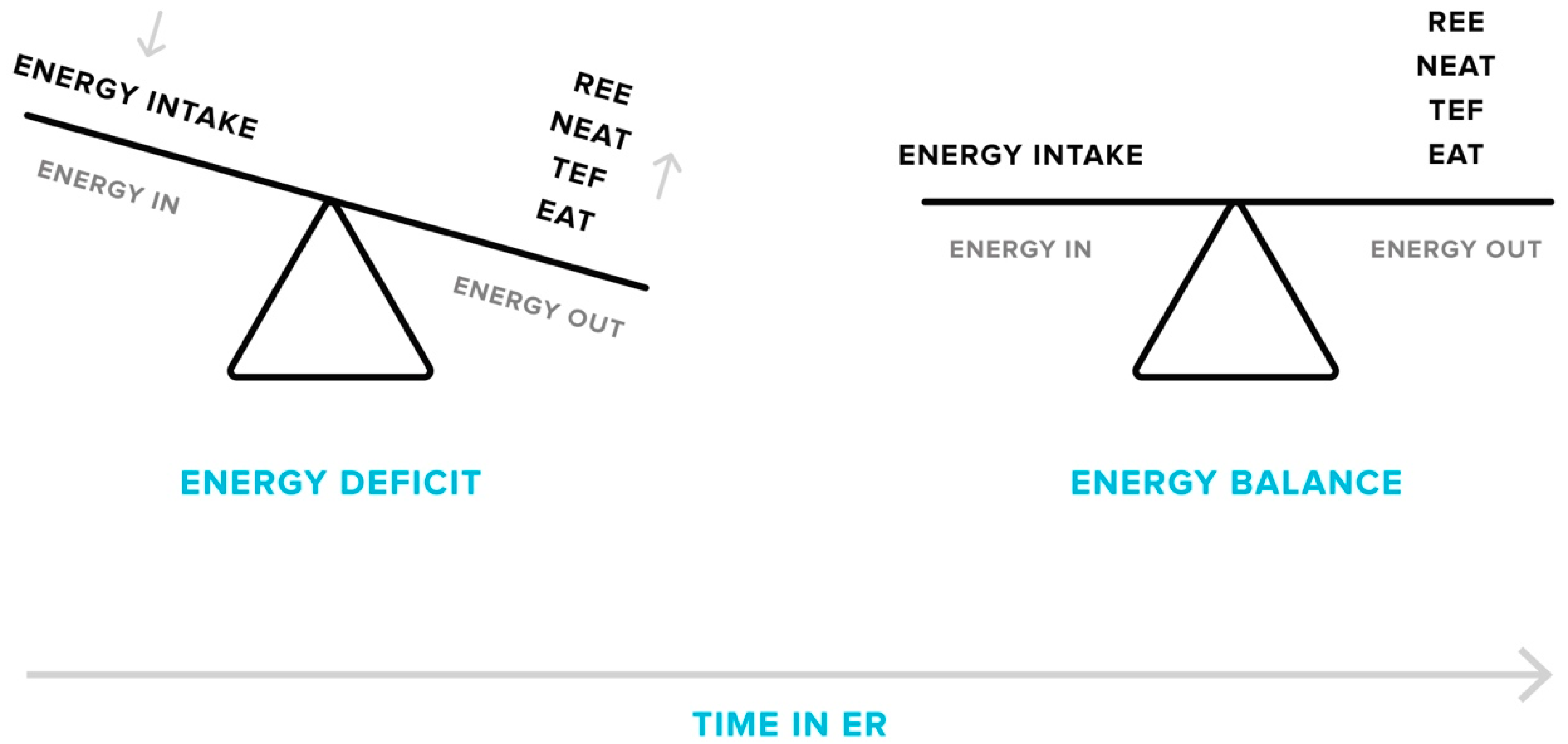
3.1. Energy Expenditure
3.2. Endocrine Responses
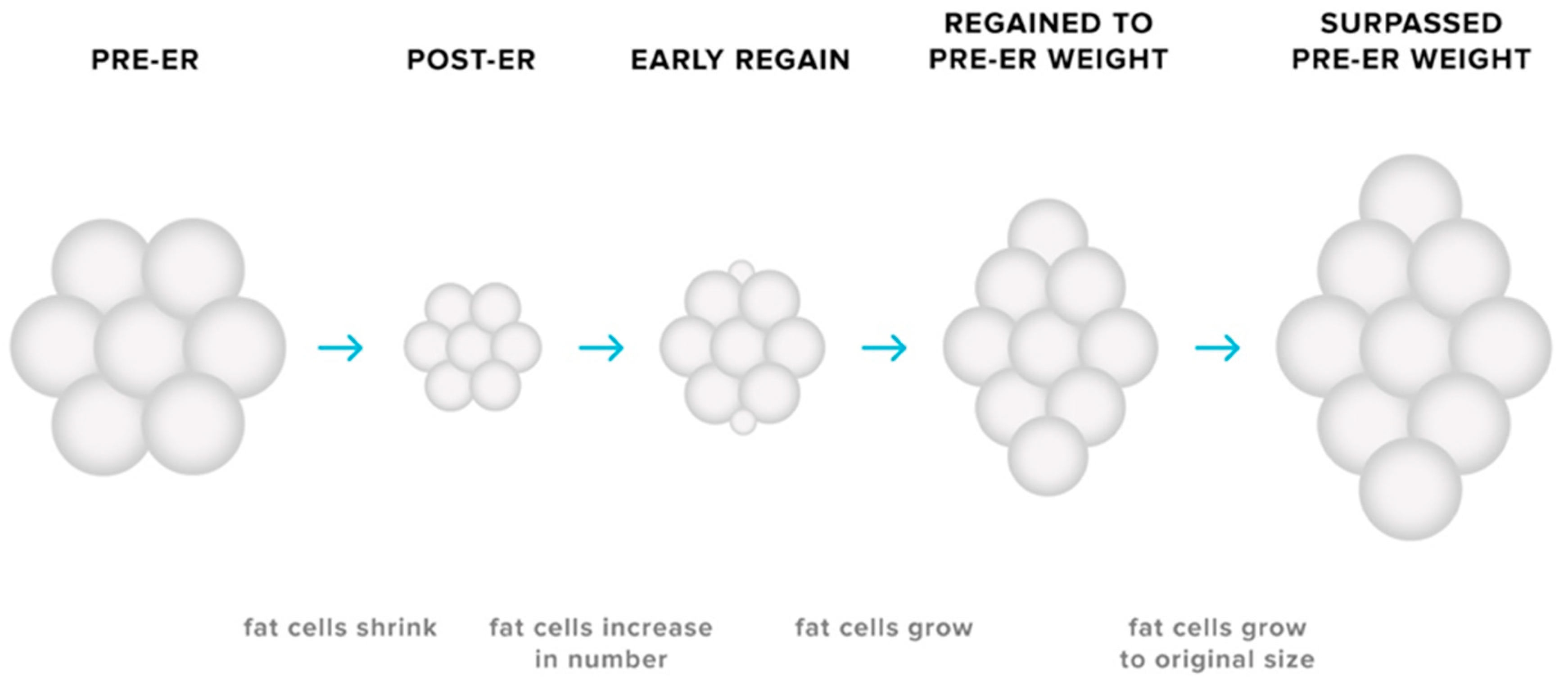
3.3. Adaptations in Adipose Tissues
4. Intermittent Energy Restriction: An Overview of Effectiveness
4.1. Intermittent Fasting
4.2. IER Regimes Involving Longer Periods of ER
4.3. Limitations of the Existing Literature
5. Practical Considerations: Intermittent Energy Restriction for Athletes
5.1. Resistance Exercise
5.2. Avoiding Severe Energy Restriction
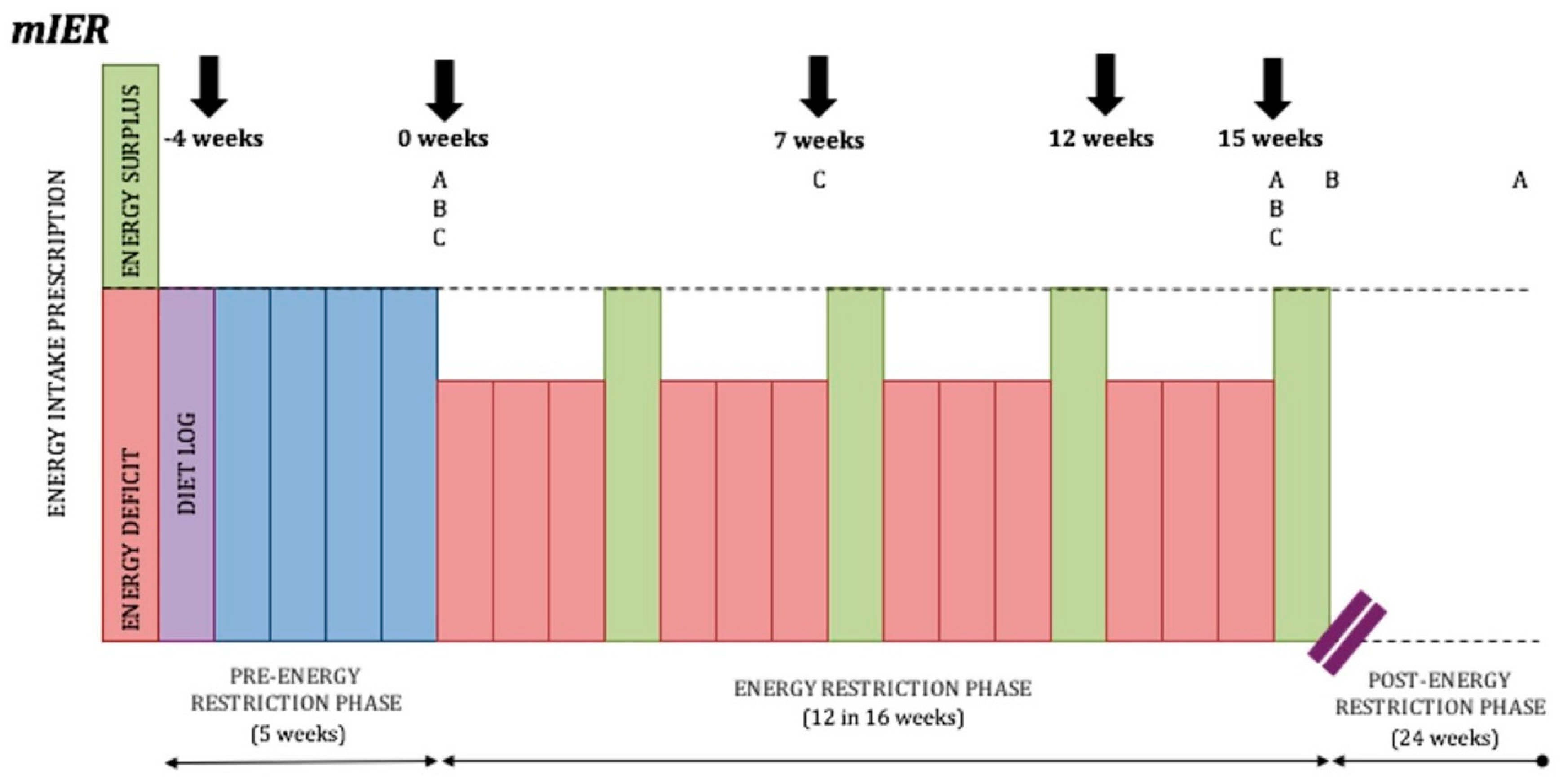
5.3. Duration of Weight Loss and Refeeds
6. Dietary Considerations: Intermittent Energy Restriction for Athletes
6.1. Protein Intake
6.2. Carbohydrate Intake
6.2.1. The Carbohydrate-Insulin Fat Loss Hypothesis
6.2.2. Performance Considerations
6.2.3. Refeeds
6.3. Fat Intake
7. Summary
- Avoid severe IER and/or rapid weight loss. Severe ER may cause greater FFM losses than moderate ER, particularly in lean athletes. Severe ER may also adversely affect health and performance outcomes including reduced muscle strength, glycogen stores, and reflexes and increase the risk of injury due to fatigue and loss of FFM. It would be practicable for an athlete to adopt a moderate level of ER that encourages absolute body weight losses of 0.5–1% per week. Alternatively, an athlete may elect to reduce energy intake by a maximum of 35% relative to energy requirements for weight maintenance.
- Resistance exercise. Athletes implementing IER should be encouraged to partake in regimented resistance exercise as a means to attenuate FFM losses. Greater retention of FFM will likely minimise performance decrement during ER and may lead to greater fat loss efficiency by mitigating compensatory reductions in REE.
- Duration and ratios of ER and refeeds. With the limited human research available, a conservative practical recommendation is to alternate two weeks of moderate ER with two weeks in EB. Currently it is unknown whether this manipulation of energy intake is ideal for maximal fat loss and FFM retention or if additional arrangements of ER and refeeds may be superior.
- Coordinating refeed periods. It may be advantageous to synchronise intervals of EB with outcome-focused or high-volume training periods. This tactic may allow the athlete to perform optimally during training sessions by providing additional nutritional support and negating the adverse consequences of sustained, daily ER.
- High protein intake. High protein intakes may be beneficial to an athlete during IER by reducing FFM losses, providing greater satiety and increasing energy expenditure through the thermic effect of feeding. A daily protein intake range between 2.3 and 3.1 g/kg of lean body mass (which equates to approximately 2.0–2.6 g per kg of absolute body mass for an 80 kg athlete with 15% body fat) is likely an appropriate practical recommendation for athletes undergoing IER with concurrent resistance exercise.
- Emphasizing carbohydrate intake during refeeds. Although yet to be confirmed, it seems wise to place emphasis on increasing intake of carbohydrate during refeed periods opposed to increasing protein or fat. Elevated levels of leptin following carbohydrate feeding may cause stimulatory effects on energy expenditure and suppress appetite, leading to greater fat loss efficiency and easier diet adherence. Greater carbohydrate availability during refeed periods may also result in more pronounced anabolic responses when IER is applied in concert with resistance exercise, potentially reducing FFM losses during ER.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Manore, M.M. Weight Management for Athletes and Active Individuals: A Brief Review. Sports Med. 2015, 45 (Suppl. 1), S83–S92. [Google Scholar] [CrossRef]
- Trexler, E.T.; Smith-Ryan, A.E.; Norton, L.E. Metabolic adaptation to weight loss: Implications for the athlete. J. Int. Soc. Sports Nutr. 2014, 11, 7. [Google Scholar] [CrossRef]
- Franchini, E.; Brito, C.J.; Artioli, G.G. Weight loss in combat sports: Physiological, psychological and performance effects. J. Int. Soc. Sports Nutr. 2012, 9, 52. [Google Scholar] [CrossRef]
- Slater, G.; Rice, A.; Jenkins, D.; Hahn, A. Body mass management of lightweight rowers: Nutritional strategies and performance implications. Br. J. Sports Med. 2014, 48, 1529–1533. [Google Scholar] [CrossRef]
- Pettersson, S.; Pipping Ekstrom, M.; Berg, C.M. The food and weight combat. A problematic fight for the elite combat sports athlete. Appetite 2012, 59, 234–242. [Google Scholar] [CrossRef]
- Wilson, G.; Hawken, M.B.; Poole, I.; Sparks, A.; Bennett, S.; Drust, B.; Morton, J.; Close, G.L. Rapid weight-loss impairs simulated riding performance and strength in jockeys: Implications for making-weight. J. Sports Sci. 2014, 32, 383–391. [Google Scholar] [CrossRef]
- Brito, C.J.; Roas, A.F.C.M.; Brito, I.S.S.; Marins, J.C.B.; Córdova, C.; Franchini, E. Methods of body mass reduction by combat sport athletes. Int. J. Sport Nutr. Exerc. Metab. 2012, 22, 89–97. [Google Scholar] [CrossRef]
- Langan-Evans, C.; Close, G.L.; Morton, J.P. Making weight in combat sports. Strength Cond. J. 2011, 33, 25–39. [Google Scholar] [CrossRef]
- Varady, K.A. Intermittent versus daily calorie restriction: Which diet regimen is more effective for weight loss? Obes. Rev. Off. J. Int. Assoc. Study Obes. 2011, 12, e593–e601. [Google Scholar] [CrossRef]
- Dixon, J.B.; Lambert, E.A.; Grima, M.; Rice, T.; Lambert, G.W.; Straznicky, N.E. Fat-free mass loss generated with weight loss in overweight and obese adults: What may we expect? Diabetes Obes. Metab. 2015, 17, 91–93. [Google Scholar] [CrossRef]
- Kouda, K.; Nakamura, H.; Kohno, H.; Okuda, T.; Higashine, Y.; Hisamori, K.; Ishihara, H.; Tokunaga, R.; Sonoda, Y. Metabolic response to short-term 4-day energy restriction in a controlled study. Environ. Health Prev. Med. 2006, 11, 89–92. [Google Scholar] [CrossRef] [Green Version]
- MacLean, P.S.; Higgins, J.A.; Giles, E.D.; Sherk, V.D.; Jackman, M.R. The role for adipose tissue in weight regain after weight loss. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2015, 16 (Suppl. 1), 45–54. [Google Scholar] [CrossRef] [Green Version]
- Maestu, J.; Eliakim, A.; Jurimae, J.; Valter, I.; Jurimae, T. Anabolic and catabolic hormones and energy balance of the male bodybuilders during the preparation for the competition. J. Strength Cond. Res. 2010, 24, 1074–1081. [Google Scholar] [CrossRef]
- Maestu, J.; Jurimae, J.; Valter, I.; Jurimae, T. Increases in ghrelin and decreases in leptin without altering adiponectin during extreme weight loss in male competitive bodybuilders. Metab. Clin. Exp. 2008, 57, 221–225. [Google Scholar] [CrossRef]
- Sloth, B.; Due, A.; Larsen, T.M.; Holst, J.J.; Heding, A.; Astrup, A. The effect of a high-MUFA, low-glycaemic index diet and a low-fat diet on appetite and glucose metabolism during a 6-month weight maintenance period. Br. J. Nutr. 2009, 101, 1846–1858. [Google Scholar] [CrossRef]
- Donnelly, J.E.; Blair, S.N.; Jakicic, J.M.; Manore, M.M.; Rankin, J.W.; Smith, B.K. American College of Sports Medicine Position Stand. Appropriate physical activity intervention strategies for weight loss and prevention of weight regain for adults. Med. Sci. Sports Exerc. 2009, 41, 459–471. [Google Scholar] [CrossRef]
- Mountjoy, M.; Junge, A.; Benjamen, S.; Boyd, K.; Diop, M.; Gerrard, D.; van den Hoogenband, C.R.; Marks, S.; Martinez-Ruiz, E.; Miller, J.; et al. Competing with injuries: Injuries prior to and during the 15th FINA World Championships 2013 (aquatics). Br. J. Sports Med. 2015, 49, 37–43. [Google Scholar] [CrossRef]
- Mountjoy, M.; Sundgot-Borgen, J.; Burke, L.; Carter, S.; Constantini, N.; Lebrun, C.; Meyer, N.; Sherman, R.; Steffen, K.; Budgett, R.; et al. The IOC consensus statement: Beyond the Female Athlete Triad--Relative Energy Deficiency in Sport (RED-S). Br. J. Sports Med. 2014, 48, 491–497. [Google Scholar] [CrossRef]
- Sundgot-Borgen, J.; Garthe, I. Elite athletes in aesthetic and Olympic weight-class sports and the challenge of body weight and body compositions. J. Sports Sci. 2011, 29 (Suppl. 1), S101–S114. [Google Scholar] [CrossRef]
- Pettersson, S.; Ekström, M.P.; Berg, C.M. Practices of weight regulation among elite athletes in combat sports: A matter of mental advantage? J. Athl. Train. 2013, 48, 99–108. [Google Scholar] [CrossRef]
- Aragon, A.A.; Schoenfeld, B.J.; Wildman, R.; Kleiner, S.; VanDusseldorp, T.; Taylor, L.; Earnest, C.P.; Arciero, P.J.; Wilborn, C.; Kalman, D.S.; et al. International society of sports nutrition position stand: Diets and body composition. J. Int. Soc. Sports Nutr. 2017, 14, 16. [Google Scholar] [CrossRef]
- Byrne, N.M.; Sainsbury, A.; King, N.A.; Hills, A.P.; Wood, R.E. Intermittent energy restriction improves weight loss efficiency in obese men: The MATADOR study. Int. J. Obes. 2017. [Google Scholar] [CrossRef]
- Harris, L.; McGarty, A.; Hutchison, L.; Ells, L.; Hankey, C. Short-term intermittent energy restriction interventions for weight management: A systematic review and meta-analysis. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2018, 19, 1–13. [Google Scholar] [CrossRef]
- Harvie, M.; Howell, A. Potential Benefits and Harms of Intermittent Energy Restriction and Intermittent Fasting Amongst Obese, Overweight and Normal Weight Subjects-A Narrative Review of Human and Animal Evidence. Behav. Sci. 2017, 7, 4. [Google Scholar] [CrossRef]
- Trepanowski, J.F.; Kroeger, C.M.; Barnosky, A.; Klempel, M.C.; Bhutani, S.; Hoddy, K.K.; Gabel, K.; Freels, S.; Rigdon, J.; Rood, J.; et al. Effect of Alternate-Day Fasting on Weight Loss, Weight Maintenance, and Cardioprotection Among Metabolically Healthy Obese Adults: A Randomized Clinical Trial. JAMA Intern. Med. 2017, 177, 930–938. [Google Scholar] [CrossRef]
- Trepanowski, J.F.; Kroeger, C.M.; Barnosky, A.; Klempel, M.C.; Bhutani, S.; Hoddy, K.K.; Rood, J.; Ravussin, E.; Varady, K.A. Effects of alternate-day fasting or daily calorie restriction on body composition, fat distribution, and circulating adipokines: Secondary analysis of a randomized controlled trial. Clin. Nutr. 2017, 37, 1871–1878. [Google Scholar] [CrossRef]
- Seimon, R.V.; Shi, Y.C.; Slack, K.; Lee, K.; Fernando, H.A.; Nguyen, A.D.; Zhang, L.; Lin, S.; Enriquez, R.F.; Lau, J.; et al. Intermittent Moderate Energy Restriction Improves Weight Loss Efficiency in Diet-Induced Obese Mice. PLoS ONE 2016, 11, e0145157. [Google Scholar] [CrossRef]
- Wing, R.R.; Jeffery, R.W. Prescribed “breaks” as a means to disrupt weight control efforts. Obes. Res. 2003, 11, 287–291. [Google Scholar] [CrossRef]
- Seimon, R.V.; Roekenes, J.A.; Zibellini, J.; Zhu, B.; Gibson, A.A.; Hills, A.P.; Wood, R.E.; King, N.A.; Byrne, N.M.; Sainsbury, A. Do intermittent diets provide physiological benefits over continuous diets for weight loss? A systematic review of clinical trials. Mol. Cell. Endocrinol. 2015, 418 Pt 2, 153–172. [Google Scholar] [CrossRef] [Green Version]
- Mettler, S.; Mitchell, N.; Tipton, K.D. Increased protein intake reduces lean body mass loss during weight loss in athletes. Med. Sci. Sports Exerc. 2010, 42, 326–337. [Google Scholar] [CrossRef]
- Elia, M.; Stubbs, R.J.; Henry, C.J. Differences in fat, carbohydrate, and protein metabolism between lean and obese subjects undergoing total starvation. Obes. Res. 1999, 7, 597–604. [Google Scholar] [CrossRef]
- Suryanarayana, B.V.; Kent, J.R.; Meister, L.; Parlow, A.F. Pituitary–gonadal axis during prolonged total starvation in obese men. Am. J. Clin. Nutr. 1969, 22, 767–770. [Google Scholar] [CrossRef]
- Heymsfield, S.B.; Gonzalez, M.C.; Shen, W.; Redman, L.; Thomas, D. Weight loss composition is one-fourth fat-free mass: A critical review and critique of this widely cited rule. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2014, 15, 310–321. [Google Scholar] [CrossRef]
- Rosenbaum, M.; Kissileff, H.R.; Mayer, L.E.; Hirsch, J.; Leibel, R.L. Energy intake in weight-reduced humans. Brain Res. 2010, 1350, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, M.; Leibel, R.L. Adaptive thermogenesis in humans. Int. J. Obes. 2010, 34 (Suppl. 1), S47–S55. [Google Scholar] [CrossRef]
- Doucet, E.; Imbeault, P.; St-Pierre, S.; Almeras, N.; Mauriege, P.; Despres, J.P.; Bouchard, C.; Tremblay, A. Greater than predicted decrease in energy expenditure during exercise after body weight loss in obese men. Clin. Sci. 2003, 105, 89–95. [Google Scholar] [CrossRef]
- Doucet, E.; St-Pierre, S.; Almeras, N.; Despres, J.P.; Bouchard, C.; Tremblay, A. Evidence for the existence of adaptive thermogenesis during weight loss. Br. J. Nutr. 2001, 85, 715–723. [Google Scholar] [CrossRef] [Green Version]
- Goele, K.; Bosy-Westphal, A.; Rumcker, B.; Lagerpusch, M.; Muller, M.J. Influence of changes in body composition and adaptive thermogenesis on the difference between measured and predicted weight loss in obese women. Obes. Facts 2009, 2, 105–109. [Google Scholar] [CrossRef]
- Anderson, J.W.; Konz, E.C.; Frederich, R.C.; Wood, C.L. Long-term weight-loss maintenance: A meta-analysis of US studies. Am. J. Clin. Nutr. 2001, 74, 579–584. [Google Scholar] [CrossRef]
- Tobey, J.A. The Biology of Human Starvation. Am. J. Public Health Nations Health 1951, 41, 236–237. [Google Scholar] [CrossRef] [Green Version]
- Weigle, D.S.; Brunzell, J.D. Assessment of energy expenditure in ambulatory reduced-obese subjects by the techniques of weight stabilization and exogenous weight replacement. Int. J. Obes. 1990, 14 (Suppl. 1), 69–77; discussion 77–81. [Google Scholar]
- Weigle, D.S.; Sande, K.J.; Iverius, P.H.; Monsen, E.R.; Brunzell, J.D. Weight loss leads to a marked decrease in nonresting energy expenditure in ambulatory human subjects. Metab. Clin. Exp. 1988, 37, 930–936. [Google Scholar] [CrossRef]
- Weyer, C.; Hanson, K.; Bogardus, C.; Pratley, R.E. Long-term changes in insulin action and insulin secretion associated with gain, loss, regain and maintenance of body weight. Diabetologia 2000, 43, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Weyer, C.; Pratley, R.E.; Salbe, A.D.; Bogardus, C.; Ravussin, E.; Tataranni, P.A. Energy expenditure, fat oxidation, and body weight regulation: A study of metabolic adaptation to long-term weight change. J. Clin. Endocrinol. Metab. 2000, 85, 1087–1094. [Google Scholar] [CrossRef]
- Weyer, C.; Walford, R.L.; Harper, I.T.; Milner, M.; MacCallum, T.; Tataranni, P.A.; Ravussin, E. Energy metabolism after 2 y of energy restriction: The biosphere 2 experiment. Am. J. Clin. Nutr. 2000, 72, 946–953. [Google Scholar] [CrossRef]
- Miles, C.W.; Wong, N.P.; Rumpler, W.V.; Conway, J. Effect of circadian variation in energy expenditure, within-subject variation and weight reduction on thermic effect of food. Eur. J. Clin. Nutr. 1993, 47, 274–284. [Google Scholar]
- Kim, H.J.; Lee, S.; Kim, T.W.; Kim, H.H.; Jeon, T.Y.; Yoon, Y.S.; Oh, S.W.; Kwak, H.; Lee, J.G. Effects of exercise-induced weight loss on acylated and unacylated ghrelin in overweight children. Clin. Endocrinol. 2008, 68, 416–422. [Google Scholar] [CrossRef]
- Silva, J.E. The thermogenic effect of thyroid hormone and its clinical implications. Ann. Intern. Med. 2003, 139, 205–213. [Google Scholar] [CrossRef]
- Sainsbury, A.; Zhang, L. Role of the hypothalamus in the neuroendocrine regulation of body weight and composition during energy deficit. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2012, 13, 234–257. [Google Scholar] [CrossRef]
- Warren, M.P. Endocrine manifestations of eating disorders. J. Clin. Endocrinol. Metab. 2011, 96, 333–343. [Google Scholar] [CrossRef]
- Rossow, L.M.; Fukuda, D.H.; Fahs, C.A.; Loenneke, J.P.; Stout, J.R. Natural bodybuilding competition preparation and recovery: A 12-month case study. Int. J. Sports Physiol. Perform. 2013, 8, 582–592. [Google Scholar] [CrossRef]
- Sainsbury, A.; Zhang, L. Role of the arcuate nucleus of the hypothalamus in regulation of body weight during energy deficit. Mol. Cell. Endocrinol. 2010, 316, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Margetic, S.; Gazzola, C.; Pegg, G.G.; Hill, R.A. Leptin: A review of its peripheral actions and interactions. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 1407–1433. [Google Scholar] [CrossRef]
- Pandit, R.; Beerens, S.; Adan, R.A.H. Role of leptin in energy expenditure: The hypothalamic perspective. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R938–R947. [Google Scholar] [CrossRef]
- Hagmar, M.; Berglund, B.; Brismar, K.; Hirschberg, A.L. Body composition and endocrine profile of male Olympic athletes striving for leanness. Clin. J. Sport Med. 2013, 23, 197–201. [Google Scholar] [CrossRef]
- Muller, M.J.; Enderle, J.; Pourhassan, M.; Braun, W.; Eggeling, B.; Lagerpusch, M.; Gluer, C.C.; Kehayias, J.J.; Kiosz, D.; Bosy-Westphal, A. Metabolic adaptation to caloric restriction and subsequent refeeding: The Minnesota Starvation Experiment revisited. Am. J. Clin. Nutr. 2015, 102, 807–819. [Google Scholar] [CrossRef]
- Koehler, K.; Hoerner, N.R.; Gibbs, J.C.; Zinner, C.; Braun, H.; De Souza, M.J.; Schaenzer, W. Low energy availability in exercising men is associated with reduced leptin and insulin but not with changes in other metabolic hormones. J. Sports Sci. 2016, 34, 1921–1929. [Google Scholar] [CrossRef]
- Rosenbaum, M.; Goldsmith, R.; Bloomfield, D.; Magnano, A.; Weimer, L.; Heymsfield, S.; Gallagher, D.; Mayer, L.; Murphy, E.; Leibel, R.L. Low-dose leptin reverses skeletal muscle, autonomic, and neuroendocrine adaptations to maintenance of reduced weight. J. Clin. Investig. 2005, 115, 3579–3586. [Google Scholar] [CrossRef] [Green Version]
- Ariyasu, H.; Takaya, K.; Tagami, T.; Ogawa, Y.; Hosoda, K.; Akamizu, T.; Suda, M.; Koh, T.; Natsui, K.; Toyooka, S.; et al. Stomach is a major source of circulating ghrelin, and feeding state determines plasma ghrelin-like immunoreactivity levels in humans. J. Clin. Endocrinol. Metab. 2001, 86, 4753–4758. [Google Scholar] [CrossRef]
- Gil-Campos, M.; Aguilera, C.M.; Canete, R.; Gil, A. Ghrelin: A hormone regulating food intake and energy homeostasis. Br. J. Nutr. 2006, 96, 201–226. [Google Scholar] [CrossRef]
- Labayen, I.; Ortega, F.B.; Ruiz, J.R.; Lasa, A.; Simon, E.; Margareto, J. Role of baseline leptin and ghrelin levels on body weight and fat mass changes after an energy-restricted diet intervention in obese women: Effects on energy metabolism. J. Clin. Endocrinol. Metab. 2011, 96, E996–E1000. [Google Scholar] [CrossRef]
- Rooyackers, O.E.; Nair, K.S. Hormonal regulation of human muscle protein metabolism. Annu. Rev. Nutr. 1997, 17, 457–485. [Google Scholar] [CrossRef]
- de Luis, D.A.; Sagrado, M.G.; Conde, R.; Aller, R.; Izaola, O. The effects of two different hypocaloric diets on glucagon-like peptide 1 in obese adults, relation with insulin response after weight loss. J. Diabetes Its Complicat. 2009, 23, 239–243. [Google Scholar] [CrossRef]
- Hietaniemi, M.; Jokela, M.; Rantala, M.; Ukkola, O.; Vuoristo, J.T.; Ilves, M.; Rysa, J.; Kesaniemi, Y. The effect of a short-term hypocaloric diet on liver gene expression and metabolic risk factors in obese women. Nutr. Metab. Cardiovasc. Dis. NMCD 2009, 19, 177–183. [Google Scholar] [CrossRef]
- Keim, N.L.; Stern, J.S.; Havel, P.J. Relation between circulating leptin concentrations and appetite during a prolonged, moderate energy deficit in women. Am. J. Clin. Nutr. 1998, 68, 794–801. [Google Scholar] [CrossRef]
- Lien, L.F.; Haqq, A.M.; Arlotto, M.; Slentz, C.A.; Muehlbauer, M.J.; McMahon, R.L.; Rochon, J.; Gallup, D.; Bain, J.R.; Ilkayeva, O.; et al. The STEDMAN project: Biophysical, biochemical and metabolic effects of a behavioral weight loss intervention during weight loss, maintenance, and regain. OMICS J. Integr. Biol. 2009, 13, 21–35. [Google Scholar] [CrossRef]
- Strohacker, K.; McCaffery, J.M.; MacLean, P.S.; Wing, R.R. Adaptations of leptin, ghrelin or insulin during weight loss as predictors of weight regain: A review of current literature. Int. J. Obes. 2014, 38, 388–396. [Google Scholar] [CrossRef]
- Ivy, J.L. Regulation of muscle glycogen repletion, muscle protein synthesis and repair following exercise. J. Sports Sci. Med. 2004, 3, 131–138. [Google Scholar]
- Friedl, K.E.; Moore, R.J.; Hoyt, R.W.; Marchitelli, L.J.; Martinez-Lopez, L.E.; Askew, E.W. Endocrine markers of semistarvation in healthy lean men in a multistressor environment. J. Appl. Physiol. 2000, 88, 1820–1830. [Google Scholar] [CrossRef]
- De Maddalena, C.; Vodo, S.; Petroni, A.; Aloisi, A.M. Impact of testosterone on body fat composition. J. Cell Physiol. 2012, 227, 3744–3748. [Google Scholar] [CrossRef]
- Mah, P.M.; Wittert, G.A. Obesity and testicular function. Mol. Cell. Endocrinol. 2010, 316, 180–186. [Google Scholar] [CrossRef]
- Smith, W.J.; Underwood, L.E.; Clemmons, D.R. Effects of caloric or protein restriction on insulin-like growth factor-I (IGF-I) and IGF-binding proteins in children and adults. J. Clin. Endocrinol. Metab. 1995, 80, 443–449. [Google Scholar]
- Redman, L.M.; Veldhuis, J.D.; Rood, J.; Smith, S.R.; Williamson, D.; Ravussin, E.; Pennington, C.T. The effect of caloric restriction interventions on growth hormone secretion in nonobese men and women. Aging Cell 2010, 9, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Simmons, P.S.; Miles, J.M.; Gerich, J.E.; Haymond, M.W. Increased proteolysis. An effect of increases in plasma cortisol within the physiologic range. J. Clin. Investig. 1984, 73, 412–420. [Google Scholar] [CrossRef]
- Zakrzewska, K.E.; Cusin, I.; Sainsbury, A.; Rohner-Jeanrenaud, F.; Jeanrenaud, B. Glucocorticoids as counterregulatory hormones of leptin: Toward an understanding of leptin resistance. Diabetes 1997, 46, 717–719. [Google Scholar] [CrossRef]
- Mastorakos, G.; Pavlatou, M. Exercise as a stress model and the interplay between the hypothalamus-pituitary-adrenal and the hypothalamus-pituitary-thyroid axes. Horm. Metab. Res. 2005, 37, 577–584. [Google Scholar] [CrossRef]
- Georgopoulos, N.A.; Rottstein, L.; Tsekouras, A.; Theodoropoulou, A.; Koukkou, E.; Mylonas, P.; Polykarpou, G.; Lampropoulou, E.; Iconomou, G.; Leglise, M.; et al. Abolished circadian rhythm of salivary cortisol in elite artistic gymnasts. Steroids 2011, 76, 353–357. [Google Scholar] [CrossRef]
- Seimon, R.V.; Hostland, N.; Silveira, S.L.; Gibson, A.A.; Sainsbury, A. Effects of energy restriction on activity of the hypothalamo-pituitary-adrenal axis in obese humans and rodents: Implications for diet-induced changes in body composition. Horm. Mol. Biol. Clin. Investig. 2013, 15, 71–80. [Google Scholar] [CrossRef]
- MacLean, P.S.; Higgins, J.A.; Jackman, M.R.; Johnson, G.C.; Fleming-Elder, B.K.; Wyatt, H.R.; Melanson, E.L.; Hill, J.O. Peripheral metabolic responses to prolonged weight reduction that promote rapid, efficient regain in obesity-prone rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1577–R1588. [Google Scholar] [CrossRef]
- MacLean, P.S.; Higgins, J.A.; Wyatt, H.R.; Melanson, E.L.; Johnson, G.C.; Jackman, M.R.; Giles, E.D.; Brown, I.E.; Hill, J.O. Regular exercise attenuates the metabolic drive to regain weight after long-term weight loss. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R793–R802. [Google Scholar] [CrossRef] [Green Version]
- Gurr, M.I.; Jung, R.T.; Robinson, M.P.; James, W.P. Adipose tissue cellularity in man: The relationship between fat cell size and number, the mass and distribution of body fat and the history of weight gain and loss. Int. J. Obes. 1982, 6, 419–436. [Google Scholar]
- Portillo, M.P.; Cantoral, R.; Macarulla, M.T. Effects of dietary fat content on adiposity during energy restriction in genetically obese rats. Reprod. Nutr. Dev. 1999, 39, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Jacobsson, B.; Smith, U. Effect of cell size on lipolysis and antilipolytic action of insulin in human fat cells. J. Lipid Res. 1972, 13, 651–656. [Google Scholar]
- Olefsky, J.M. Effects of fasting on insulin binding, glucose transport, and glucose oxidation in isolated rat adipocytes: Relationships between insulin receptors and insulin action. J. Clin. Investig. 1976, 58, 1450–1460. [Google Scholar] [CrossRef]
- Bjorntorp, P.; Carlgren, G.; Isaksson, B.; Krotkiewski, M.; Larsson, B.; Sjostrom, L. Effect of an energy-reduced dietary regimen in relation to adipose tissue cellularity in obese women. Am. J. Clin. Nutr. 1975, 28, 445–452. [Google Scholar] [CrossRef]
- Lofgren, P.; Hoffstedt, J.; Naslund, E.; Wiren, M.; Arner, P. Prospective and controlled studies of the actions of insulin and catecholamine in fat cells of obese women following weight reduction. Diabetologia 2005, 48, 2334–2342. [Google Scholar] [CrossRef]
- Svensson, P.A.; Gabrielsson, B.G.; Jernas, M.; Gummesson, A.; Sjoholm, K. Regulation of human aldoketoreductase 1C3 (AKR1C3) gene expression in the adipose tissue. Cell. Mol. Biol. Lett. 2008, 13, 599–613. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Wu, Y.; Fried, S.K. Adipose tissue remodeling in pathophysiology of obesity. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 371–376. [Google Scholar] [CrossRef] [Green Version]
- Mariman, E.C. Human biology of weight maintenance after weight loss. J. Nutr. Nutr. 2012, 5, 13–25. [Google Scholar] [CrossRef]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Naslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef]
- Jackman, M.R.; Steig, A.; Higgins, J.A.; Johnson, G.C.; Fleming-Elder, B.K.; Bessesen, D.H.; MacLean, P.S. Weight regain after sustained weight reduction is accompanied by suppressed oxidation of dietary fat and adipocyte hyperplasia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R1117–R1129. [Google Scholar] [CrossRef]
- Barnosky, A.R.; Hoddy, K.K.; Unterman, T.G.; Varady, K.A. Intermittent fasting vs daily calorie restriction for type 2 diabetes prevention: A review of human findings. Transl. Res. 2014, 164, 302–311. [Google Scholar] [CrossRef]
- Moro, T.; Tinsley, G.; Bianco, A.; Marcolin, G.; Pacelli, Q.F.; Battaglia, G.; Palma, A.; Gentil, P.; Neri, M.; Paoli, A. Effects of eight weeks of time-restricted feeding (16/8) on basal metabolism, maximal strength, body composition, inflammation, and cardiovascular risk factors in resistance-trained males. J. Transl. Med. 2016, 14, 290. [Google Scholar] [CrossRef]
- Varady, K.A.; Allister, C.A.; Roohk, D.J.; Hellerstein, M.K. Improvements in body fat distribution and circulating adiponectin by alternate-day fasting versus calorie restriction. J. Nutr. Biochem. 2010, 21, 188–195. [Google Scholar] [CrossRef]
- Varady, K.A.; Bhutani, S.; Church, E.C.; Klempel, M.C. Short-term modified alternate-day fasting: A novel dietary strategy for weight loss and cardioprotection in obese adults. Am. J. Clin. Nutr. 2009, 90, 1138–1143. [Google Scholar] [CrossRef]
- Varady, K.A.; Bhutani, S.; Klempel, M.C.; Kroeger, C.M.; Trepanowski, J.F.; Haus, J.M.; Hoddy, K.K.; Calvo, Y. Alternate day fasting for weight loss in normal weight and overweight subjects: A randomized controlled trial. Nutr. J. 2013, 12, 146. [Google Scholar] [CrossRef]
- Davis, C.S.; Clarke, R.E.; Coulter, S.N.; Rounsefell, K.N.; Walker, R.E.; Rauch, C.E.; Huggins, C.E.; Ryan, L. Intermittent energy restriction and weight loss: A systematic review. Eur. J. Clin. Nutr. 2016, 70, 292–299. [Google Scholar] [CrossRef]
- Davoodi, S.H.; Ajami, M.; Ayatollahi, S.A.; Dowlatshahi, K.; Javedan, G.; Pazoki-Toroudi, H.R. Calorie shifting diet versus calorie restriction diet: A comparative clinical trial study. Int. J. Prev. Med. 2014, 5, 447–456. [Google Scholar]
- Harris, L.; Hamilton, S.; Azevedo, L.B.; Olajide, O.J.; De Brun, C.; Waller, G.; Whittaker, V.J.; Sharp, T.; Lean, M.; Hankey, C.; et al. Intermittent fasting interventions for the treatment of overweight and obesity in adults aged 18 years and over: A systematic review and meta-analysis. JBI Database Syst. Rev. Implement. Rep. 2018, 16, 507–547. [Google Scholar] [CrossRef]
- Campbell, B.I.; Aguilar, D.; Colenso-Semple, L.; Hartke, K.; Gai, C.; Gaviria, D.; Gorman, J.; Rubio, J.; Ibrahim, A.; Barker, B. The effects of intermittent carbohydrate re-feeds vs. continuous dieting on body composition in resistance trained individuals: A flexible dieting study. In Proceedings of the 15th International Society of Sports Nutrition (ISSN) Conference and Expo, Clearwater Beach, FL, USA, 7–9 June 2018. [Google Scholar]
- Campbell, B.I.; Aguilar, D.; Wong, V.; Rogers, G.; Fox, C.; Hosey, T.; Lindsay, S.; Ford, S.; Radford, D. The effects of intermittent carbohydrate re-feeds vs. continuous dieting on resting metabolic rate in resistance trained individuals: A flexible dieting study. In Proceedings of the 15th International Society of Sports Nutrition (ISSN) Conference and Expo, Clearwater Beach, FL, USA, 7–9 June 2018. [Google Scholar]
- Coelho do Vale, R.; Pieters, R.; Zeelenberg, M. The benefits of behaving badly on occasion: Successful regulation by planned hedonic deviations. J. Consum. Psychol. 2016, 26, 17–28. [Google Scholar] [CrossRef]
- Keogh, J.B.; Pedersen, E.; Petersen, K.S.; Clifton, P.M. Effects of intermittent compared to continuous energy restriction on short-term weight loss and long-term weight loss maintenance. Clin. Obes. 2014, 4, 150–156. [Google Scholar] [CrossRef]
- Bosy-Westphal, A.; Schautz, B.; Lagerpusch, M.; Pourhassan, M.; Braun, W.; Goele, K.; Heller, M.; Glüer, C.C.; Müller, M.J. Effect of weight loss and regain on adipose tissue distribution, composition of lean mass and resting energy expenditure in young overweight and obese adults. Int. J. Obes. 2013, 37, 1371. [Google Scholar] [CrossRef]
- Ravussin, E.; Lillioja, S.; Knowler, W.C.; Christin, L.; Freymond, D.; Abbott, W.G.H.; Boyce, V.; Howard, B.V.; Bogardus, C. Reduced Rate of Energy Expenditure as a Risk Factor for Body-Weight Gain. N. Engl. J. Med. 1988, 318, 467–472. [Google Scholar] [CrossRef]
- Peos, J.J.; Helms, E.R.; Fournier, P.A.; Sainsbury, A. Continuous versus intermittent moderate energy restriction for increased fat mass loss and fat free mass retention in adult athletes: Protocol for a randomised controlled trial-the ICECAP trial (Intermittent versus Continuous Energy restriction Compared in an Athlete Population). BMJ Open Sport Exerc. Med. 2018, 4, e000423. [Google Scholar] [PubMed]
- Henry, R.R.; Scheaffer, L.; Olefsky, J.M. Glycemic effects of intensive caloric restriction and isocaloric refeeding in noninsulin-dependent diabetes mellitus. J. Clin. Endocrinol. Metab. 1985, 61, 917–925. [Google Scholar] [CrossRef]
- Wing, R.R.; Blair, E.H.; Bononi, P.; Marcus, M.D.; Watanabe, R.; Bergman, R.N. Caloric restriction per se is a significant factor in improvements in glycemic control and insulin sensitivity during weight loss in obese NIDDM patients. Diabetes Care 1994, 17, 30–36. [Google Scholar] [CrossRef]
- Collet, T.H.; Sonoyama, T.; Henning, E.; Keogh, J.M.; Ingram, B.; Kelway, S.; Guo, L.; Farooqi, I.S. A Metabolomic Signature of Acute Caloric Restriction. J. Clin. Endocrinol. Metab. 2017, 102, 4486–4495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulloo, A.G.; Jacquet, J. Adaptive reduction in basal metabolic rate in response to food deprivation in humans: A role for feedback signals from fat stores. Am. J. Clin. Nutr. 1998, 68, 599–606. [Google Scholar] [CrossRef]
- Kolaczynski, J.W.; Considine, R.V.; Ohannesian, J.; Marco, C.; Opentanova, I.; Nyce, M.R.; Myint, M.; Caro, J.F. Responses of leptin to short-term fasting and refeeding in humans: A link with ketogenesis but not ketones themselves. Diabetes 1996, 45, 1511–1515. [Google Scholar] [CrossRef] [PubMed]
- Schebendach, J.E.; Golden, N.H.; Jacobson, M.S.; Hertz, S.; Shenker, I.R. The metabolic responses to starvation and refeeding in adolescents with anorexia nervosa. Ann. N. Y. Acad. Sci. 1997, 817, 110–119. [Google Scholar] [CrossRef]
- Tsintzas, K.; Jewell, K.; Kamran, M.; Laithwaite, D.; Boonsong, T.; Littlewood, J.; Macdonald, I.; Bennett, A. Differential regulation of metabolic genes in skeletal muscle during starvation and refeeding in humans. J. Physiol. 2006, 575, 291–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinsier, R.L.; Nagy, T.R.; Hunter, G.R.; Darnell, B.E.; Hensrud, D.D.; Weiss, H.L. Do adaptive changes in metabolic rate favor weight regain in weight-reduced individuals? An examination of the set-point theory. Am. J. Clin. Nutr. 2000, 72, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Mathieson, R.A.; Walberg, J.L.; Gwazdauskas, F.C.; Hinkle, D.E.; Gregg, J.M. The effect of varying carbohydrate content of a very-low-caloric diet on resting metabolic rate and thyroid hormones. Metab. Clin. Exp. 1986, 35, 394–398. [Google Scholar] [CrossRef]
- Chin-Chance, C.; Polonsky, K.S.; Schoeller, D.A. Twenty-four-hour leptin levels respond to cumulative short-term energy imbalance and predict subsequent intake. J. Clin. Endocrinol. Metab. 2000, 85, 2685–2691. [Google Scholar] [CrossRef] [PubMed]
- Douyon, L.; Schteingart, D.E. Effect of obesity and starvation on thyroid hormone, growth hormone, and cortisol secretion. Endocrinol. Metab. Clin N. Am. 2002, 31, 173–189. [Google Scholar] [CrossRef]
- Dirlewanger, M.; di Vetta, V.; Guenat, E.; Battilana, P.; Seematter, G.; Schneiter, P.; Jequier, E.; Tappy, L. Effects of short-term carbohydrate or fat overfeeding on energy expenditure and plasma leptin concentrations in healthy female subjects. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 1413–1418. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, L.; Hackett, D.; Gifford, J.; Estermann, F.; O’Connor, H. Do Bodybuilders Use Evidence-Based Nutrition Strategies to Manipulate Physique? Sports 2017, 5, 76. [Google Scholar] [CrossRef]
- Helms, E.R.; Fitschen, P.J.; Aragon, A.A.; Cronin, J.; Schoenfeld, B.J. Recommendations for natural bodybuilding contest preparation: Resistance and cardiovascular training. J. Sports Med. Phys. Fit. 2015, 55, 164–178. [Google Scholar]
- Kistler, B.M.; Fitschen, P.J.; Ranadive, S.M.; Fernhall, B.; Wilund, K.R. Case study: Natural bodybuilding contest preparation. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 694–700. [Google Scholar] [CrossRef]
- Nindl, B.C.; Barnes, B.R.; Alemany, J.A.; Frykman, P.N.; Shippee, R.L.; Friedl, K.E. Physiological consequences of U.S. Army Ranger training. Med. Sci. Sports Exerc. 2007, 39, 1380–1387. [Google Scholar] [CrossRef]
- Koutedakis, Y.; Pacy, P.J.; Quevedo, R.M.; Millward, D.J.; Hesp, R.; Boreham, C.; Sharp, N.C. The effects of two different periods of weight-reduction on selected performance parameters in elite lightweight oarswomen. Int. J. Sports Med. 1994, 15, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.W.; Haub, M.D.; Wolfe, R.R.; Ferrando, A.A.; Sullivan, D.H.; Apolzan, J.W.; Iglay, H.B. Resistance training preserves fat-free mass without impacting changes in protein metabolism after weight loss in older women. Obesity 2009, 17, 1332–1339. [Google Scholar] [CrossRef] [PubMed]
- Verreijen, A.M.; Engberink, M.F.; Memelink, R.G.; van der Plas, S.E.; Visser, M.; Weijs, P.J. Effect of a high protein diet and/or resistance exercise on the preservation of fat free mass during weight loss in overweight and obese older adults: A randomized controlled trial. Nutr. J. 2017, 16. [Google Scholar] [CrossRef]
- Bryner, R.W.; Ullrich, I.H.; Sauers, J.; Donley, D.; Hornsby, G.; Kolar, M.; Yeater, R. Effects of resistance vs. aerobic training combined with an 800 calorie liquid diet on lean body mass and resting metabolic rate. J. Am. Coll. Nutr. 1999, 18, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Garthe, I.; Raastad, T.; Refsnes, P.E.; Koivisto, A.; Sundgot-Borgen, J. Effect of two different weight-loss rates on body composition and strength and power-related performance in elite athletes. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Mero, A.A.; Huovinen, H.; Matintupa, O.; Hulmi, J.J.; Puurtinen, R.; Hohtari, H.; Karila, T.A. Moderate energy restriction with high protein diet results in healthier outcome in women. J. Int. Soc. Sports Nutr. 2010, 7, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval, W.M.; Heyward, V.H.; Lyons, T.M. Comparison of body composition, exercise and nutritional profiles of female and male body builders at competition. J. Sports Med. Phys. Fit. 1989, 29, 63–70. [Google Scholar]
- Withers, R.T.; Noell, C.J.; Whittingham, N.O.; Chatterton, B.E.; Schultz, C.G.; Keeves, J.P. Body composition changes in elite male bodybuilders during preparation for competition. Aust. J. Sci. Med. Sport. 1997, 29, 11–16. [Google Scholar] [CrossRef]
- van der Ploeg, G.E.; Brooks, A.G.; Withers, R.T.; Dollman, J.; Leaney, F.; Chatterton, B.E. Body composition changes in female bodybuilders during preparation for competition. Eur. J. Clin. Nutr. 2001, 55, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Forbes, G.B. Lean body mass-body fat interrelationships in humans. Nutr. Rev. 1987, 45, 225–231. [Google Scholar] [CrossRef]
- Anderson, J. Obesity. Br. Med. J. 1972, 1, 560–563. [Google Scholar] [CrossRef]
- Antoni, R.; Johnston, K.L.; Collins, A.L.; Robertson, M.D. Investigation into the acute effects of total and partial energy restriction on postprandial metabolism among overweight/obese participants. Br. J. Nutr. 2016, 115, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Bailey, B.W.; Jacobsen, D.J.; Donnelly, J.E. Weight loss and maintenance outcomes using moderate and severe caloric restriction in an outpatient setting. Dis. Manag. 2008, 11, 176–180. [Google Scholar] [CrossRef]
- Christensen, P.; Bliddal, H.; Riecke, B.F.; Leeds, A.R.; Astrup, A.; Christensen, R. Comparison of a low-energy diet and a very low-energy diet in sedentary obese individuals: A pragmatic randomized controlled trial. Clin. Obes. 2011, 1, 31–40. [Google Scholar] [CrossRef]
- Helms, E.R.; Aragon, A.A.; Fitschen, P.J. Evidence-based recommendations for natural bodybuilding contest preparation: Nutrition and supplementation. J. Int. Soc. Sports Nutr. 2014, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Gibson, A.A.; Seimon, R.V.; Franklin, J.; Markovic, T.P.; Byrne, N.M.; Manson, E.; Caterson, I.D.; Sainsbury, A. Fast versus slow weight loss: Development process and rationale behind the dietary interventions for the TEMPO Diet Trial. Obes. Sci. Pract. 2016, 2, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Byrne, N.M.; Weinsier, R.L.; Hunter, G.R.; Desmond, R.; Patterson, M.A.; Darnell, B.E.; Zuckerman, P.A. Influence of distribution of lean body mass on resting metabolic rate after weight loss and weight regain: Comparison of responses in white and black women. Am. J. Clin. Nutr. 2003, 77, 1368–1373. [Google Scholar] [CrossRef] [PubMed]
- Byrne, N.M.; Hills, A.P. Biology or Behavior: Which Is the Strongest Contributor to Weight Gain? Curr. Obes. Rep. 2013, 2, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Elliot, D.L.; Goldberg, L.; Kuehl, K.S.; Bennett, W.M. Sustained depression of the resting metabolic rate after massive weight loss. Am. J. Clin. Nutr. 1989, 49, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Hulmi, J.J.; Isola, V.; Suonpaa, M.; Jarvinen, N.J.; Kokkonen, M.; Wennerstrom, A.; Nyman, K.; Perola, M.; Ahtiainen, J.P.; Hakkinen, K. The Effects of Intensive Weight Reduction on Body Composition and Serum Hormones in Female Fitness Competitors. Front. Physiol. 2016, 7, 689. [Google Scholar] [CrossRef] [PubMed]
- Chappell, A.J.; Simper, T.; Barker, M.E. Nutritional strategies of high level natural bodybuilders during competition preparation. J. Int. Soc. Sports Nutr. 2018, 15, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loucks, A.B.; Verdun, M. Slow restoration of LH pulsatility by refeeding in energetically disrupted women. Am. J. Physiol. 1998, 275, R1218–R1226. [Google Scholar] [CrossRef] [PubMed]
- Olson, B.R.; Cartledge, T.; Sebring, N.; Defensor, R.; Nieman, L. Short-term fasting affects luteinizing hormone secretory dynamics but not reproductive function in normal-weight sedentary women. J. Clin. Endocrinol. Metab. 1995, 80, 1187–1193. [Google Scholar] [PubMed]
- Hu, T.; Mills, K.T.; Yao, L.; Demanelis, K.; Eloustaz, M.; Yancy, W.S., Jr.; Kelly, T.N.; He, J.; Bazzano, L.A. Effects of low-carbohydrate diets versus low-fat diets on metabolic risk factors: A meta-analysis of randomized controlled clinical trials. Am. J. Epidemiol. 2012, 176 (Suppl. 7), S44–S54. [Google Scholar] [CrossRef] [PubMed]
- Position of the American Dietetic Association, Dietitians of Canada, and the American College of Sports Medicine: Nutrition and athletic performance. J. Am. Diet. Assoc. 2000, 100, 1543–1556. [CrossRef]
- Jequier, E. Leptin signaling, adiposity, and energy balance. Ann. N. Y. Acad. Sci. 2002, 967, 379–388. [Google Scholar] [CrossRef]
- Jequier, E. Pathways to obesity. Int. J. Obes. Relat. Metab. Disord. 2002, 26 (Suppl. 2), S12–S17. [Google Scholar] [CrossRef]
- Paddon-Jones, D.; Westman, E.; Mattes, R.D.; Wolfe, R.R.; Astrup, A.; Westerterp-Plantenga, M. Protein, weight management, and satiety. Am. J. Clin. Nutr. 2008, 87, 1558s–1561s. [Google Scholar] [CrossRef]
- Pasiakos, S.M.; Cao, J.J.; Margolis, L.M.; Sauter, E.R.; Whigham, L.D.; McClung, J.P.; Rood, J.C.; Carbone, J.W.; Combs, G.F., Jr.; Young, A.J. Effects of high-protein diets on fat-free mass and muscle protein synthesis following weight loss: A randomized controlled trial. FASEB J. 2013, 27, 3837–3847. [Google Scholar] [CrossRef]
- Weigle, D.S.; Breen, P.A.; Matthys, C.C.; Callahan, H.S.; Meeuws, K.E.; Burden, V.R.; Purnell, J.Q. A high-protein diet induces sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations. Am. J. Clin. Nutr. 2005, 82, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.M. Dietary protein for athletes: From requirements to metabolic advantage. Appl. Physiol. Nutr. Metab. 2006, 31, 647–654. [Google Scholar] [CrossRef]
- Phillips, S.M.; Van Loon, L.J. Dietary protein for athletes: From requirements to optimum adaptation. J. Sports Sci. 2011, 29 (Suppl. S1), S29–S38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemon, P.W. Beyond the zone: Protein needs of active individuals. J. Am. Coll. Nutr. 2000, 19, 513s–521s. [Google Scholar] [CrossRef]
- Tipton, K.D.; Wolfe, R.R. Protein and amino acids for athletes. J. Sports Sci. 2004, 22, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, D.S. Lowering the Bar on the Low-Fat Diet. JAMA 2016, 316, 2087–2088. [Google Scholar] [CrossRef] [PubMed]
- Boaz, M.; Raz, O. Low Fat vs. Low Carbohydrate Diet Strategies for Weight Reduction: A Meta-Analysis. J. Obes. Weight Loss Ther. 2015, 5. [Google Scholar] [CrossRef]
- Hall, K.D. A review of the carbohydrate-insulin model of obesity. Eur. J. Clin. Nutr. 2017, 71, 323–326. [Google Scholar] [CrossRef]
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. Position of the Academy of Nutrition and Dietetics, Dietitians of Canada, and the American College of Sports Medicine: Nutrition and Athletic Performance. J. Acad. Nutr. Diet. 2016, 116, 501–528. [Google Scholar] [CrossRef]
- Cole, M.; Coleman, D.; Hopker, J.; Wiles, J. Improved gross efficiency during long duration submaximal cycling following a short-term high carbohydrate diet. Int. J. Sports Med. 2014, 35, 265–269. [Google Scholar] [CrossRef]
- Spriet, L.L. New insights into the interaction of carbohydrate and fat metabolism during exercise. Sports Med. 2014, 44 (Suppl. 1), S87–S96. [Google Scholar] [CrossRef]
- Leveritt, M.; Abernethy, P.J.; Barry, B.K.; Logan, P.A. Concurrent strength and endurance training. A review. Sports Med. 1999, 28, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Ross, M.L.; Garvican-Lewis, L.A.; Welvaert, M.; Heikura, I.A.; Forbes, S.G.; Mirtschin, J.G.; Cato, L.E.; Strobel, N.; Sharma, A.P.; et al. Low carbohydrate, high fat diet impairs exercise economy and negates the performance benefit from intensified training in elite race walkers. J. Physiol. 2017, 595, 2785–2807. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Hawley, J.A.; Wong, S.H.; Jeukendrup, A.E. Carbohydrates for training and competition. J. Sports Sci. 2011, 29 (Suppl. 1), S17–S27. [Google Scholar] [CrossRef] [Green Version]
- Haff, G.G.; Lehmkuhl, M.J.; McCoy, L.B.; Stone, M.H. Carbohydrate supplementation and resistance training. J. Strength Cond. Res. 2003, 17, 187–196. [Google Scholar] [PubMed]
- Escobar, K.A.; Morales, J.; Vandusseldorp, T.A. The Effect of a Moderately Low and High Carbohydrate Intake on Crossfit Performance. Int. J. Exerc. Sci. 2016, 9, 460–470. [Google Scholar]
- Jenkins, A.B.; Markovic, T.P.; Fleury, A.; Campbell, L.V. Carbohydrate intake and short-term regulation of leptin in humans. Diabetologia 1997, 40, 348–351. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Rhodes, C.J.; Lawrence, J.C., Jr. Activation of mammalian target of rapamycin (mTOR) by insulin is associated with stimulation of 4EBP1 binding to dimeric mTOR complex 1. J. Biol. Chem. 2006, 281, 24293–24303. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.P.; Tarpenning, K.M.; Marino, F.E. Independent and combined effects of liquid carbohydrate/essential amino acid ingestion on hormonal and muscular adaptations following resistance training in untrained men. Eur. J. Appl. Physiol. 2006, 97, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.E.; Goodpaster, B.; Wing, R.R.; Simoneau, J.A. Skeletal muscle fatty acid metabolism in association with insulin resistance, obesity, and weight loss. Am. J. Physiol. 1999, 277, E1130–E1141. [Google Scholar] [CrossRef]
- Kelley, D.E.; Goodpaster, B.H.; Storlien, L. Muscle triglyceride and insulin resistance. Annu. Rev. Nutr. 2002, 22, 325–346. [Google Scholar] [CrossRef]
- Astrup, A.; Buemann, B.; Christensen, N.J.; Madsen, J. 24-hour energy expenditure and sympathetic activity in postobese women consuming a high-carbohydrate diet. Am. J. Physiol. 1992, 262, E282–E288. [Google Scholar] [CrossRef] [PubMed]
- Ballor, D.L.; Harvey-Berino, J.R.; Ades, P.A.; Cryan, J.; Calles-Escandon, J. Decrease in fat oxidation following a meal in weight-reduced individuals: A possible mechanism for weight recidivism. Metab. Clin. Exp. 1996, 45, 174–178. [Google Scholar] [CrossRef]
- Bessesen, D.H.; Rupp, C.L.; Eckel, R.H. Dietary fat is shunted away from oxidation, toward storage in obese Zucker rats. Obes. Res. 1995, 3, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Steig, A.J.; Jackman, M.R.; Giles, E.D.; Higgins, J.A.; Johnson, G.C.; Mahan, C.; Melanson, E.L.; Wyatt, H.R.; Eckel, R.H.; Hill, J.O.; et al. Exercise reduces appetite and traffics excess nutrients away from energetically efficient pathways of lipid deposition during the early stages of weight regain. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R656–R667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schutz, Y. Dietary fat, lipogenesis and energy balance. Physiol. Behav. 2004, 83, 557–564. [Google Scholar] [CrossRef]
- Dorgan, J.F.; Judd, J.T.; Longcope, C.; Brown, C.; Schatzkin, A.; Clevidence, B.A.; Campbell, W.S.; Nair, P.P.; Franz, C.; Kahle, L.; et al. Effects of dietary fat and fiber on plasma and urine androgens and estrogens in men: A controlled feeding study. Am. J. Clin. Nutr. 1996, 64, 850–855. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M. Re-Examining High-Fat Diets for Sports Performance: Did We Call the ‘Nail in the Coffin’ Too Soon? Sports Med. 2015, 45 (Suppl. 1), S33–S49. [Google Scholar] [CrossRef]
- Walberg, J.L.; Leidy, M.K.; Sturgill, D.J.; Hinkle, D.E.; Ritchey, S.J.; Sebolt, D.R. Macronutrient content of a hypoenergy diet affects nitrogen retention and muscle function in weight lifters. Int. J. Sports Med. 1988, 9, 261–266. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peos, J.J.; Norton, L.E.; Helms, E.R.; Galpin, A.J.; Fournier, P. Intermittent Dieting: Theoretical Considerations for the Athlete. Sports 2019, 7, 22. https://doi.org/10.3390/sports7010022
Peos JJ, Norton LE, Helms ER, Galpin AJ, Fournier P. Intermittent Dieting: Theoretical Considerations for the Athlete. Sports. 2019; 7(1):22. https://doi.org/10.3390/sports7010022
Chicago/Turabian StylePeos, Jackson James, Layne Eiseman Norton, Eric Russell Helms, Andrew Jacob Galpin, and Paul Fournier. 2019. "Intermittent Dieting: Theoretical Considerations for the Athlete" Sports 7, no. 1: 22. https://doi.org/10.3390/sports7010022
APA StylePeos, J. J., Norton, L. E., Helms, E. R., Galpin, A. J., & Fournier, P. (2019). Intermittent Dieting: Theoretical Considerations for the Athlete. Sports, 7(1), 22. https://doi.org/10.3390/sports7010022