1. Introduction
Epidemics caused by viral infections, such as the ongoing COVID-19 pandemic and previous outbreaks of the severe acute respiratory syndrome (SARS) in 2003, H1N1 influenza in 2009, and Middle East respiratory syndrome (MERS) in 2014, have highlighted the significance of airborne transmission of contagious diseases through respiratory droplets and aerosols. This mode of transmission is particularly important for indoor settings with limited ventilation in which respiratory aerosol concentrations can build up [
1,
2]. Exposure to biological aerosols may result in detrimental health outcomes, such as respiratory, neurological, and cardiovascular ailments [
3]. According to the World Health Organization (WHO), COVID-19 had resulted in approximately
million fatalities and around 761 million recorded cases of infection as of March 2023 [
4]. To understand the impact of the disease and infection caused by inhaled pathogens, it is important to understand the emission source, the survival rate of the pathogen in the environment, the mode of transmission, as well as the dose, duration, and route of exposure. While many studies have investigated the transmission risk from respiratory virus emissions [
5], the transmission risk from direct resuspension of infectious viruses that have previously settled onto surfaces is unknown.
Once emitted into the indoor air, respiratory aerosols undergo evaporation, reducing their size, and are removed from the air over minutes to hours via ventilation, filtration, and deposition onto surfaces [
5]. Human activities such as walking, sitting on furniture, and cleaning can resuspend settled particles back into the air [
6]. Many studies have been conducted to characterize particle resuspension from human activity [
7]. Summarizing the resuspension emission rates reported by previous studies, Ferro (2022) [
7] showed that emission rates vary over several orders of magnitude, even for the same size particles. For example, the reported resuspension emission rates for 1
m particles range from
to
for full-scale studies.
Resuspension is a major source of airborne bacteria and fungi in classrooms and other indoor microenvironments [
8,
9]. Similar to other indoor aerosol sources, the concentrations are the highest closest to the source. Khare and Marr (2015) [
10] simulated the resuspension of influenza virus in dust via human walking and estimated that the concentration of the resuspended virus would be higher closer to the floor than at breathing height. Hyytiainen et al. (2018) [
11] reported that bacteria and fungal levels were 8 to 21 fold higher in the breathing zone of a crawling infant robot compared to those measured in the breathing zone of an adult. Numerous other investigations on the resuspension of bioaerosols, primarily bacteria and fungi, have been conducted within the last two decades [
9,
12,
13,
14,
15,
16,
17,
18]. However, the resuspension of viruses or particles containing viruses via human activity has not been studied experimentally.
Particle size is a primary factor for determining particle resuspension from human activity [
6]. Within the particle size range most relevant to human health (particle diameter, or Dp, 10
m and smaller), larger particles resuspend more easily from surfaces than smaller particles. This is because of their smaller ratio of the adhesive force to the detachment force when an external detachment force, such as the squeezed flow beneath a footstep, is applied [
6,
19]. Therefore, due to the relatively small size of viruses (Dp 20–200 nm), we expect the resuspension fraction (fraction of particles on the surface that are resuspended) for viruses to be less than that for larger bioaerosols, such as bacteria and fungi. However, the viral particles are associated with larger carrier particles due to the initial emission of liquid respiratory aerosol as well as the interaction of the liquid aerosol with other existing indoor particles. Therefore, the transport, deposition, and resuspension dynamics of viruses are governed by the properties of the carrier particles. The size distribution and composition of respiratory viral aerosol emissions have been well characterized [
5,
20,
21,
22,
23], but characterization of the carrier particles for resuspended viruses has not been reported in the literature.
The survival rate of different viruses in aerosols and on different surfaces is also an important factor for assessing the importance of virus resuspension, with inactivation dependent on temperature, relative humidity (RH), and ultraviolet radiation [
24,
25]. Yang and Marr (2011) [
26] developed a material balance model with inactivation as an additional loss term to predict infectious influenza resuspension due to human walking. Using a mechanistic biochemical model of virus inactivation kinetics, Morris et al. (2021) [
24] demonstrated that viral inactivation rates for SARS-CoV-2, influenza, and other enveloped viruses are dependent on both temperature and RH, with shorter survival times for enveloped viruses at higher temperatures and RH above a threshold relative humidity. Van Doremalen et al. (2020) [
27] studied the stability of SARS-CoV-1 and SARS-CoV-2 in the laboratory and concluded that these viruses can remain viable and infectious for hours in aerosols and up to days on surfaces. Although determining the infectivity of viruses in aerosols in the field is challenging due to the sensitivity of the viruses to the collection and analysis methods, several studies have found infectious SARS-CoV-2 in aerosols in environments with infected patients [
28,
29,
30,
31].
There is additional evidence that viral resuspension could be an important exposure pathway. A study by Asadi et al. (2020) [
32] found that influenza-spiked dust on the fur of guinea pigs was resuspended by the movement of the guinea pigs and the resuspended virus was transmitted to guinea pigs in adjacent cages. Furthermore, Liu et al. (2020) [
33] found relatively high virus concentrations in hospital staff changing rooms and bathrooms compared with patient rooms, suggesting that some resuspension due to human activity occurred. Thus, further investigation of viral resuspension is important to quantify this exposure pathway for humans.
The present study addresses the need for experimental investigation into virus resuspension. In this study, we nebulized a buffer solution containing live influenza A virus into a full-scale laboratory. After allowing the particles to settle onto indoor surfaces, one person entered the room to resuspend the particles. We measured the airborne concentrations of viruses during the initial emission period and the resuspension activity periods to quantify and determine the relative importance of the resuspension source as compared with direct emission.
2. Materials and Methods
We conducted this study during July–August 2021 in a biosafety level 2 (BSL-2) laboratory at Trudeau Institute, Saranac Lake, NY. The study protocol was approved for BSL-2, which was required due to the use of a live infectious virus (Approval number B-384-21, Trudeau Institute). The experimental setup consisted of nebulizing liquid phosphate bovine serum (PBS) particles containing live virions of influenza A virus x31 H3N2 strain into a
laboratory room. After the nebulized virus-laden particles were allowed to settle for
±
h, an investigator entered, carefully conducted sampling activities for approximately 10 min to collect the pre-resuspension samples and initiate the resuspension samples, and then walked continuously in the room for 20 min to resuspend the particles. We collected the viral particles in impingers and on deposition plates at multiple locations inside the room (
Figure 1 and
Figure 2) during and for approximately
after the virus emission period as well as during and for approximately
after the resuspension activity and compared the results. The experiments were conducted first with clean surfaces such that the nebulized viral aerosol would deposit directly onto the surfaces (clean surface condition). The experiments were also conducted after the emission and deposition of inorganic test dust such that some of the nebulized aerosols would deposit onto the settled dust before the resuspension activity (dusty surface condition). Prior to the clean surface condition and dusty surface condition experiments, an initial experiment was performed that included only the nebulized aerosol emission and no resuspension activity (no resuspension condition).
2.1. Room Preparation and Decontamination
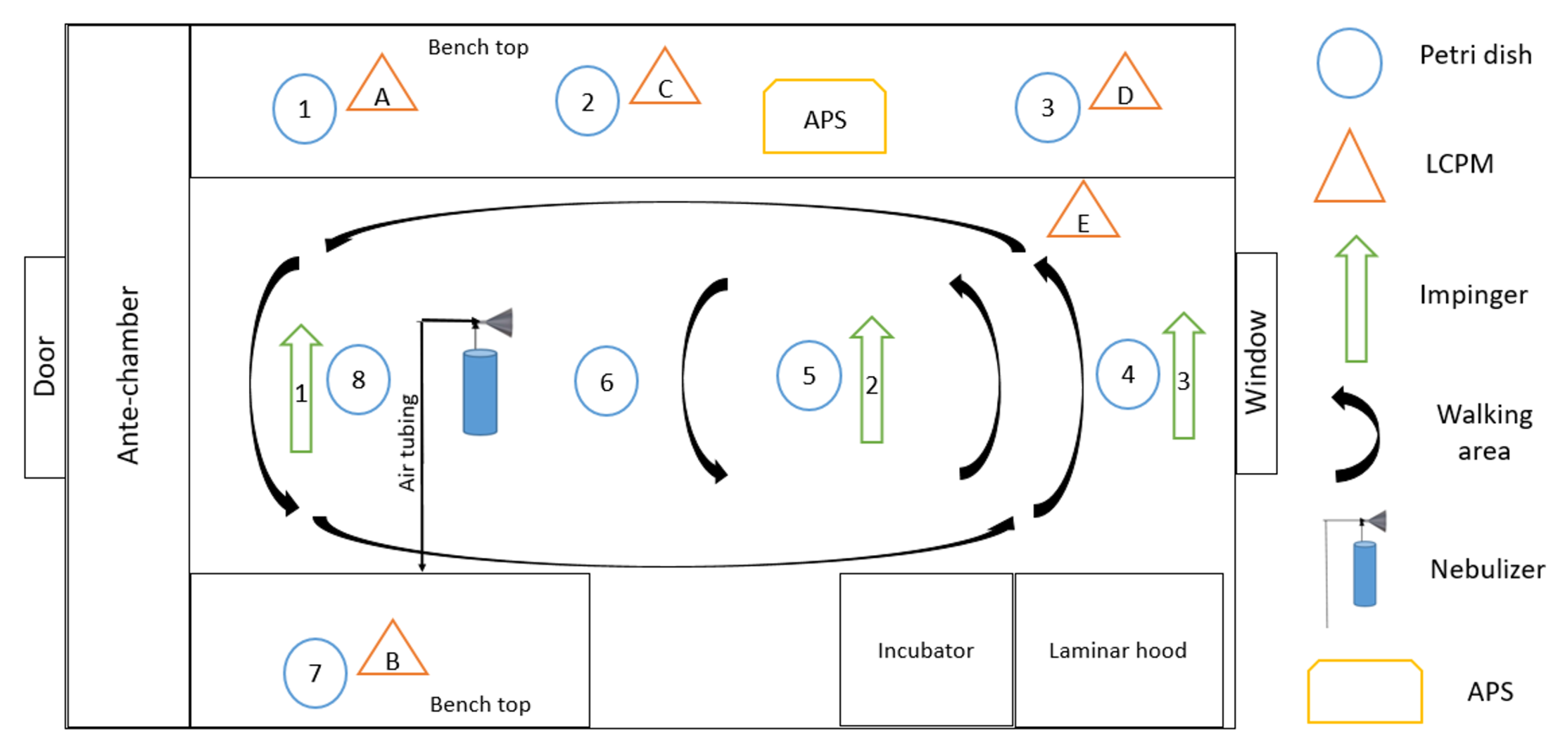
Figure 2 provides a schematic of the
(L) ×
(W) ×
(H) room with vinyl tile flooring used for the study. In accordance with the biosafety protocol, the room ventilation system was turned off and covered with plastic sheeting. Before starting the experiments, we removed all portable laboratory equipment and covered the walls, cabinets lining the wall, and non-portable laboratory equipment with plastic sheeting to avoid viral contamination of the non-target surfaces. We expect there were additional particle losses to the plastic sheeting compared with a more conductive material such as aluminum foil [
34]. The particle deposition rate, which accounts for the deposition on all surfaces in the room, was estimated from the particle concentration time series and incorporated in the resuspension modeling, as described below in
Section 2.10.
Using additional plastic sheeting, we created an ante-chamber inside the laboratory by the door to the hallway. The ante-chamber was used as a doffing area for personal protective equipment (PPE) worn inside the experimental area and as a staging area for the air sampling. Accounting for the benches, cabinets, large equipment, and volume separated by sheeting, the mixing volume of the room was estimated to be . Before the experiments, all the surfaces in the room were wiped down using a mop (floor) and sponge (benchtop) with lab-prepared ethanol solution. Following the experiments, all the surfaces inside the room were thoroughly cleaned with disinfectant (Quatricide PV-15, Pharmacal, Waterbury, CT, USA).
2.2. Virus Generation and Sampling
Liquid aerosol containing influenza A x31 H3N2 strain was generated at a height of
using a
stainless steel nebulizer. We placed 7 mL of virus stock in the nebulizer, which had a concentration of
×
egg infectious dose
for a total of 1 ×
. Approximately 3 mL of the stock solution was nebulized, resulting in a maximum nebulized amount of
×
. Pressurized air at
was provided to the nebulizer by a compressed air cylinder stored in the adjacent room and connected to the nebulizer via tubing. The nebulizer was run for 30 min, resulting in an emission rate of
×
viral RNA copies
. To deposit a sufficient number of viruses on the flooring such that virus resuspension from human walking could be quantified, the emission rate is approximately 100 times higher than estimates for respiratory viral emissions for a single, infected person. As a comparison, Stadnytskyi et al. (2020) [
35] estimated the viral emission rate for a person speaking who is infected with SARS-CoV-2 to be
copies
based on respiratory aerosol emissions and viral content in the sputum; Ma et al. (2021) [
36] estimated SARS-CoV-2 viral emission to be 2 ×
–4 ×
copies
from 52 exhaled breath condensate samples collected from 49 patients; and Wang et al. (2020) [
37] estimated the SARS-CoV-2 emissions from a single cough to be
copies.
The particle volume (and mass) size distribution measured in the laboratory during the virus emission period ranged from
to 5
m, a mean particle volume diameter of 2
m, and a mean particle number diameter of
m. The influenza A virus, contained within the nebulized PBS, is 80–120
m [
38].
Supplementary Information Figure S1A,B provides detailed information regarding the particle size distributions of the nebulized aerosol without dust. Additionally,
Supplementary Information Figure S2B,E presents the particle size distributions for the nebulized aerosol and dust.
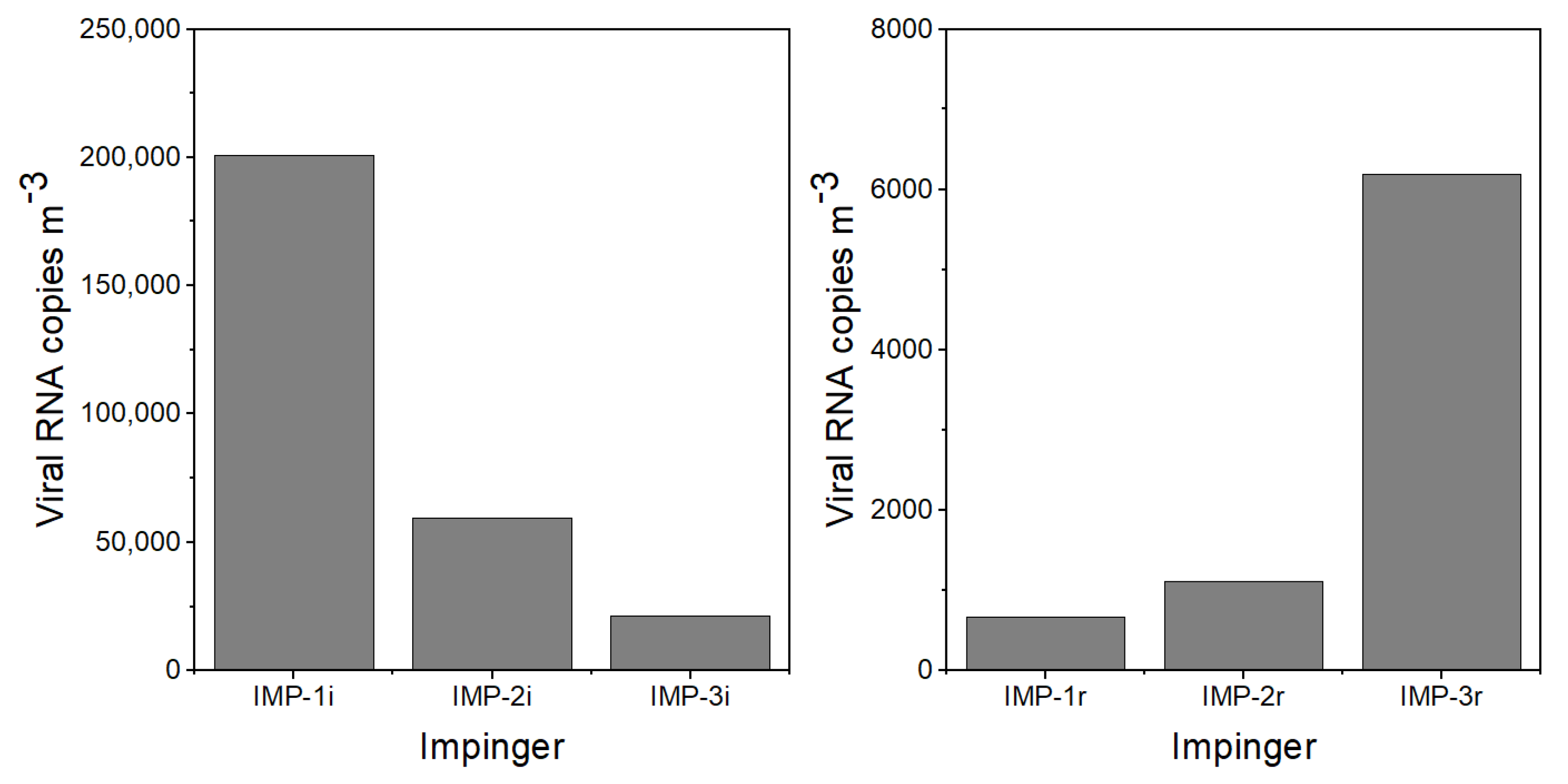
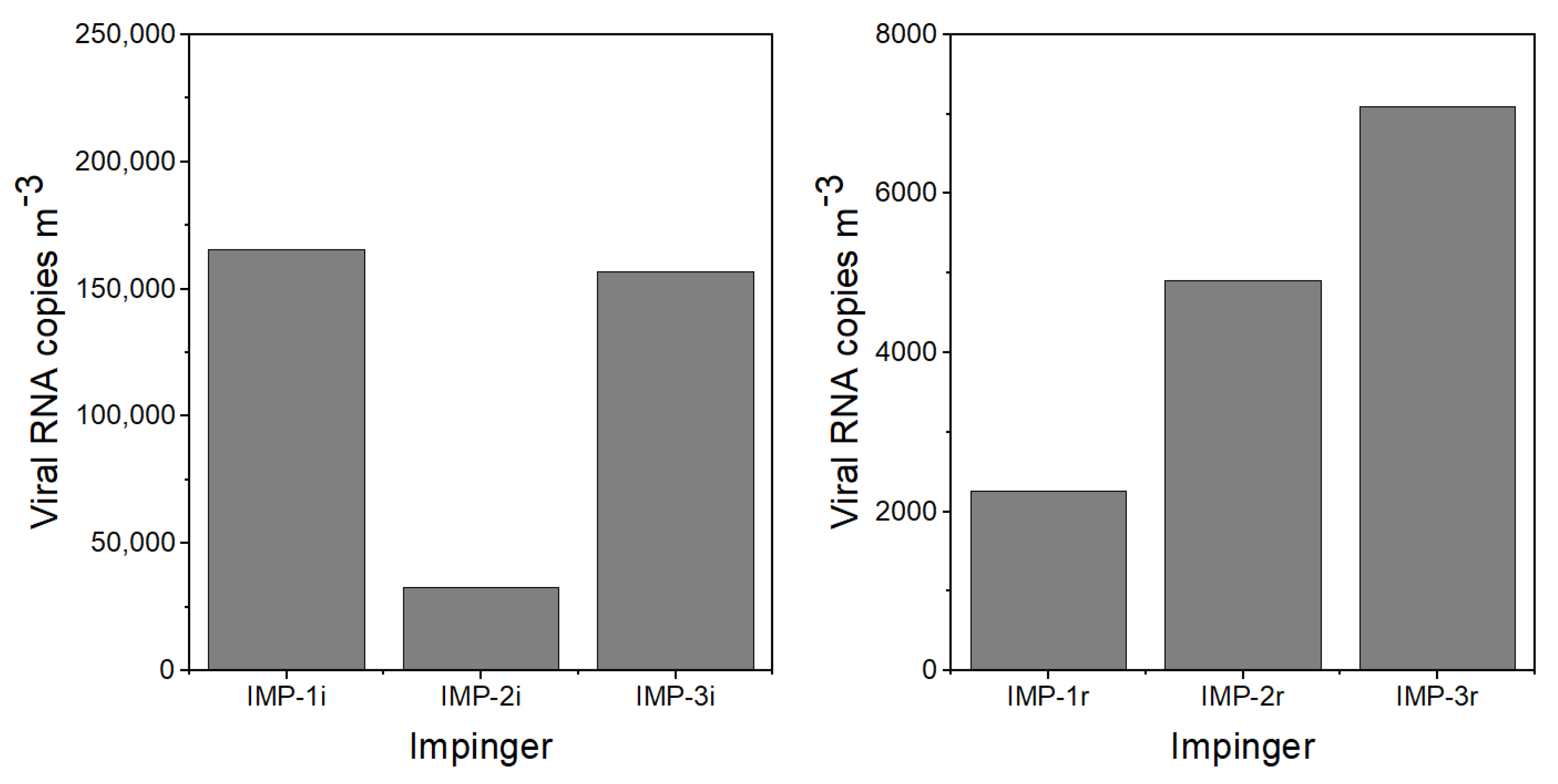
The viral aerosol was collected by 3 Glass Midget Impingers (SKC Inc., Eighty Four, PA, USA) containing 7 mL PBS. The impingers were placed along the center aisle of the room
behind the nebulizer and
and
in front of the nebulizer, all at a height of
. The impinger pumps were calibrated to the target flow rate of 3 L m
prior to each experiment with a DryCal DC-Lite primary flow meter (BIOS International Corporation, NJ, USA), and the pump flow rates were checked at the beginning and end of each experiment. The impinger pumps for the initial virus emission samples were turned on prior to the nebulizer and turned off at the beginning of the resuspension activity. The impinger pumps for the resuspension activity samples were turned on at the beginning of the resuspension activity and turned off after approximately
h. The individual pump flow rates and sampling times are provided in
Supplementary Information Tables S1–S3.
Deposition samples were collected with
diameter Petri dishes (Falcon standard tissue culture dishes) containing
of PBS placed in 8 different locations throughout the room (
Figure 2). The viral RNA copies per mL were multiplied by
and divided by the area of the Petri dish to determine the viral loading (viral RNA copies
).
2.3. Test Dust Generation
Most indoor surfaces have a layer of dust on them. Based on particle adhesion theory and results from previous experimental studies, resuspension can be enhanced for multilayer deposits compared with monolayer deposits [
39,
40]. Thus, for the current study, we simulated a dusty surface condition and compared the results with those for the clean surface condition. A solid aerosol generator model 410 by TOPAS GmbH was used to introduce a total of 6 g of ISO 12103-1 A1 ultrafine test dust into the room at a feed rate of 2.5 and pressure level set at 137.9 kPa for 40 min. Assuming approximately 55% of the emitted dust deposited on the indoor surfaces versus removal via air exchange, and 80% of the deposited dust settled on horizontal surfaces, the estimated dust loading was 0.2
. This is within the range of typical dust loading values reported in the literature for hard flooring in real environments of 0.1–1
[
39,
41,
42].
The dust has a volume particle size range of
to 10
m in diameter, with a mean volume (and mass) particle diameter of 5
m and a mean particle number diameter of
m. Following the emission, we allowed the generated dust to mix in the room air and settle for
before generating the virus (as described in the previous section). During the 1.5 h decay period, most of the test dust particles had settled onto surfaces or were removed via the air exchange. As measured by a TSI (Shoreline, MN, USA) Model 3321 aerodynamic particle sizer (APS), the mass concentration of
had decreased by 97%,
had decreased by 88%, and
had decreased by 86% when the virus was emitted. The particle size distribution plot for the generated dust aerosol is provided in the
Supplementary Information Figure S2A,D.
2.4. Aerosol Monitoring
An array of aerosol monitors was placed in the room, including the TSI APS and five PurpleAir (Draper, UT, USA) Model PA-II low-cost particle monitors (LCPMs). The APS uses time-of-flight technology to count and size particles aerodynamically from 0.5–20
m as well as a light scattering to estimate the particle concentration from 0.37–20
m. The APS sample time was set to 1 min. Given the experimental conditions, the liquid particles would be expected to reach equilibrium by the time they are measured by the APS [
20]. The PurpleAir LCPMs use optical sensors to estimate
,
, and
with a sampling time of approximately 1 min. The values included in our analyses are for
. The APS and LCPMs were turned on two hours before the release of aerosol into the room and turned off the morning after the experiment.
The APS and LCPMs were used to determine the aerosol concentration throughout the room during the different phases of the experiment. A collocation experiment was conducted with the APS and the LCPMs placed together to compare and adjust the calibration of the LCPMs so that they could be directly compared during the experiments to assess the mixing conditions of the room (see
Supplementary Information Figures S3 and S4).
2.5. Relative Humidity, Temperature, and Carbon Dioxide Measurement
Relative humidity and temperature were measured using Onset (Bourne, MA, USA) HOBO Model X temperature/RH data loggers. During the experiments, the temperature (30 ± C) and RH ( ± %) remained relatively constant. The laboratory was on the north side of the building and there was no direct sunlight coming into the room. Carbon dioxide (CO2) was measured in two locations in the room using Telaire (Amphenol Advanced Sensors, St. Marys, PA, USA) Model T7000 Series handheld indoor air quality (IAQ) monitors. CO2 was used to determine the air change rate in the room. We emitted CO2 into the room by opening a container of dry ice for 5–10 min to increase the CO2 levels. We turned on a fan during the emission period to improve mixing conditions in the room. After the emission, we let the concentration decay for 1 h. We performed this experiment 3 times on three different days. We estimated the air change rate in the room to be ± h−1 using the slope of the natural log of the CO2 concentration decay. The air change rate was also determined during the resuspension experiments using the decay of the CO2 increase from the human occupant respiratory emissions.
2.6. Experimental Protocol for Initial Virus Emission and Resuspension Activity
The experimental protocol for the experiments was as follows: (1) Two hours before the virus emission, the APS and LCPMs were turned on to record the background particle concentration and size distribution. (2) A few minutes before the nebulizer was turned on to initiate the virus emission, impingers IMP-1i, IMP-2i, and IMP-3i were turned on and Petri dishes P-1i–P-8i were uncovered. (3) The nebulizer containing virus-laden PBS was turned on for 30 min. (4) Following the emission, the room was left empty and quiescent to allow the particles to settle. (5) After 5 h, an investigator entered the room wearing PPE kit, including an N95 mask and Tyvek coveralls with a hood and booties, to collect the initial emission impingers and Petri dish samples, start impingers IMP-1r, IMP-2r, and IMP-3r, and uncover Petri dishes P-1r–P-8r. This activity took approximately 10 min. (6) The investigator performed a walking activity across the room for 20 min to resuspend the particles from the floor. (7) The investigator left the room and the room was left untouched overnight. (8) Impingers IMP-1r, IMP-2r, and IMP-3r were turned off after approximately h and remained undisturbed for approximately 10 h. (9) Resuspension activity samples were collected and transferred the following morning to perform the viral RNA extraction and PCR analysis.
For the dusty surface condition experiment, the test dust was released prior to the emission of the nebulized viral aerosol. For the no resuspension condition experiment, steps 5 through 9 were omitted.
2.7. PCR Analysis
After collection, the impinger and Petri dish samples were immediately transferred and stored at
C. The viral RNA was extracted from sample material and collected in an elution buffer using QIAamp viral RNA mini kits (QIAGEN, Germantown, MD, USA) prior to RT-PCR. PCR amplification of the Influenza A polymerase (PA) gene was performed using the SuperScript™ III Platinum™ One-Step qRT-PCR kit (Invitrogen, Waltham, MA, USA). The primers used were as shown: 5′ -/56-FAM/CCA AGT CAT/ ZEN/ GAA GGA GAG GGA ATA CCG CT/3 IAB kFQ/-3′ (probe), 5′ -CGG TCC AAA TTC CTG CTG AT-3′ (forward), 5′ -CAT TGG GTT CCT TCC ATC CA-3′ (reverse). A known concentration of PA-containing plasmid was used to generate a standard curve in all reactions [
43]. The prepared reactions were run with a standard cycling program of
C for
min,
C for 2 s, followed by 44 cycles of
C for 15 s, and
C for 30 s.
The number of viral RNA copies
was calculated using Equation (1):
where the viral RNA copies
was determined from 1 mL of the impinger sample collected during the experiment, Ct is the cycle threshold value that was estimated using RT-PCR, and the slope and intercept were from the RNA copies dilution standard curve with dilutions (
to
) prepared from the
viral stock (the assay was capable of detecting as low as 10 viral RNA copies
).
The number of viral RNA copies
of air was calculated using Equation (2):
where viral RNA copies
is calculated using Equation (1),
is the total volume in the impinger (mL) at the end of the sampling period, Q is the flow rate of the impinger pump (L m
), and t is the sampling time (min).
The number of viral RNA copies
for the deposition samples was calculated using Equation (3):
where viral RNA copies
is calculated using Equation (1),
is the total volume in the Petri dish (mL), and
is the surface area of the Petri dish (
).
2.8. Viability Analysis
We attempted to determine the viability of the collected samples using a Madin–Darby canine kidney assay (MDCK). MDCK cells were seeded in complete minimal essential media (MEM, Corning, Glendale, AZ) supplemented with fetal bovine serum (FBS) in a 96-well plate at a density of cells per well. After 24 h, complete MEM was removed and replaced with Trypsin-Zero-Serum Refeed Medium (mg trypsin/EDTA in Zero-Serum Refeed Medium, Diagnostic Hybrids, Athens, OH, USA). The virus was diluted in a separate plate from to as a control and added to the MDCK monolayer while samples obtained from the experiments were added neat and diluted 2-fold. The plate was spun to allow the virus to adhere and the media was replaced with fresh Trypsin-Zero-Serum Refeed Medium, and the plate was incubated at C for 24 h. After incubation, the cells were fixed in acetone and stained for viral replication using a biotinylated mouse anti-nucleoprotein antibody (Millipore Sigma, Burlington, MA) followed by streptavidin-phycoerythrin (BD Biosciences, San Jose, CA, USA). We visualized the viral foci using an Immunospot Plate Reader (CTL, Shaker Heights, OH, USA) at 580 nm and/or 625 nm. During one of our reduced-scale pilot studies, we were able to detect viable viruses for a nebulization experiment conducted in a small (<1 m3) chamber and relatively short time period (≈1 h). However, we did not achieve positive results for the room-scale experiments. This may be due to the degradation of x31 virus over the longer period of time as well as from the nebulizer emission and sampling protocols.
2.9. Modeling the Contribution of Initial Emission to Resuspension Activity Measurements
A limitation of these findings is a result of the sampling time associated with the approved BSL-2 protocol. Because investigators could not be in the facility after working hours, the resuspension activity needed to be initiated before the particle concentration returned completely to the background concentration following the initial nebulized virus emission. Therefore, some of the viruses that were collected during the resuspension activity sampling period may have been viruses remaining in the air from the initial emission or may have come from surfaces other than the flooring. However, we expect that this contribution was minimal. The investigator conducted the resuspension activity wearing clean PPE, limiting the contribution of resuspended viral particles from the clothing. Furthermore, the activity was focused on the flooring, not on the other surfaces. A previous study conducted by Bhangar et al. (2016) [
44], found that the flooring was responsible for ≈
of the resuspension emissions during a chamber study that included human activity.
To estimate the relative contribution of the initial virus emission and the resuspension activity to the measured concentration, we applied a one-compartment material balance model for indoor particle dynamics [
45] to the APS
particle concentration time series. In addition to the assumptions required for applying the material balance model, we assumed that the ratio of viral concentration to the airborne particle concentration remained constant throughout the decay of the initial emission source. This ratio was then applied to the modeled concentration decay of the initial emission source after the resuspension activity had begun (
Supplementary Figures S5 and S6). The approach for estimating the contribution of a previous particle source to the increase in concentration from a current particle source using a concentration time series is described in detail in Ferro et al. (2004) [
46].
2.10. Resuspension Modeling
A basic resuspension model was applied to estimate the expected resuspension of the settled viral particles due to human walking, as shown in Equation (4) and described in Tian et al. (2014) [
40].
where
is the resuspension emission rate for particle size
j (RNA copies
),
is the stepping rate (steps
),
is the contact area of the shoe with the flooring (
),
is the floor loading for particle size
j (RNA copies
), and
is the resuspension fraction per footstep for particle size
j. The resuspension emission rate is then applied to a material balance model to estimate the concentration of resuspended particles in the air during the resuspension activity sampling period, as shown in Equation (5).
where
is the viral concentration for particle size j at time t (RNA copies
),
is the steady-state concentration for particle size j (copies
),
is the initial concentration for particle size j (RNA copies
),
a is the air change rate (
), and
is the deposition rate for particle size j (
).
is calculated as per Equation (6).
where
is calculated as per Equation (4) and
V is the mixing volume of the room (
).
The inputs to the model are provided in
Supplementary Information Table S4 and the results from this analysis are provided in
Supplementary Information Table S5. For the present study, the stepping rate and contact area of the shoe were determined for the investigator performing the resuspension activity and the floor loading was estimated from the deposition samples. We selected parameter values based on estimates from the scientific literature and did not conduct further parameter fitting. Because resuspension is strongly dependent on particle size and the viral surface loading estimates are not size-resolved, we used the size distribution for the initial nebulized virus emission (
Figures S1 and S2) to divide the measured viral surface loading results into different size bins (nominal particle diameters of
,
,
, and
m). We selected resuspension fraction values toward the center of the range for non-biological particles, as summarized by Ferro (2022) [
7]. These were
,
,
, and
for nominal particle diameters of
,
,
, and
m, respectively. The viral resuspension was estimated for the four size bins and then totaled.
4. Discussion
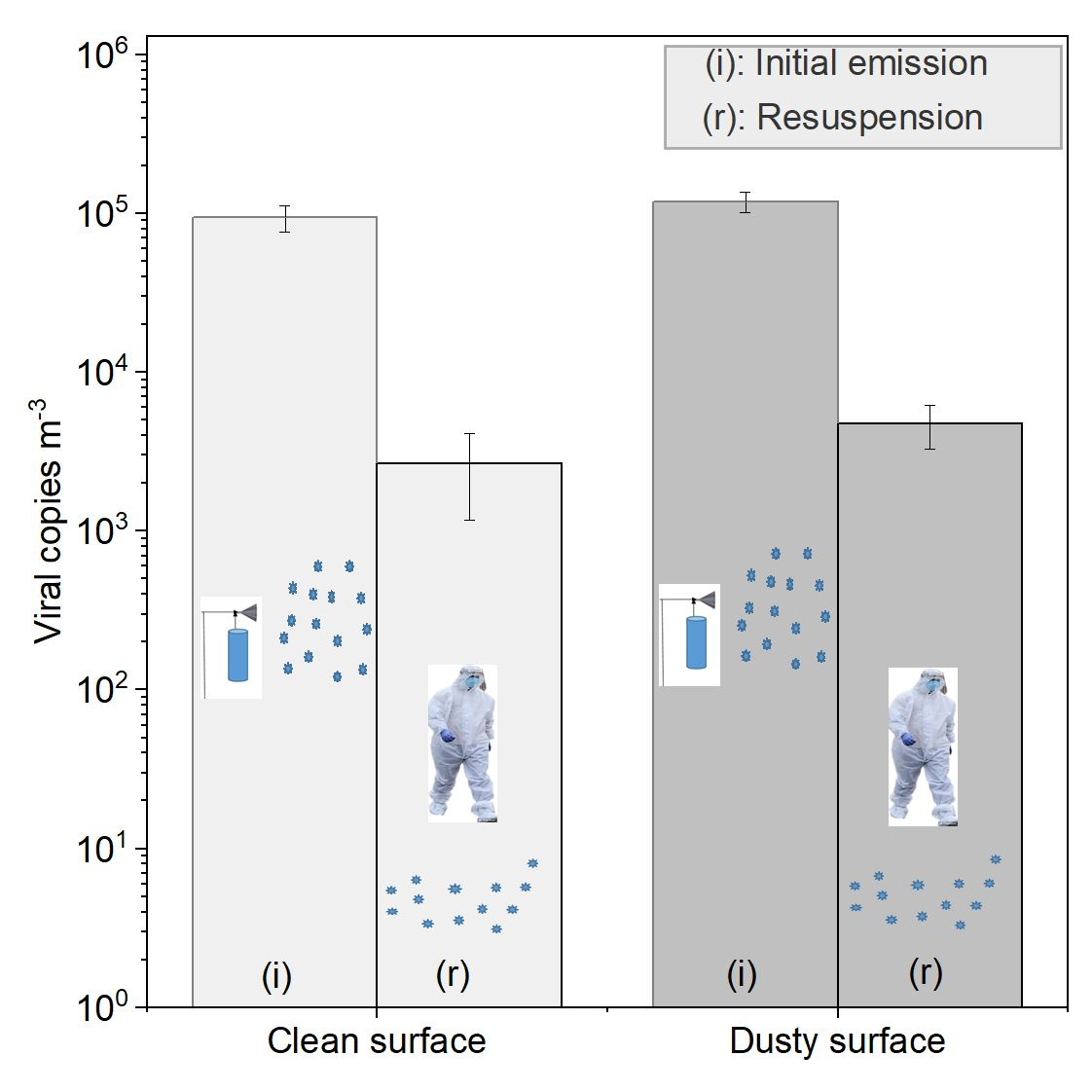
The findings from this study support the hypothesis that the influenza virus can be resuspended via human activity. For our experiments, the concentration of the resuspended, airborne virus was approximately two orders of magnitude lower than the concentration during the initial emission of the virus into the room. Different scenarios, such as changes in viral loading, air change rate, volume of the room, type and length of the resuspension activity, shoe type, surface types, temperature and RH, etc., would adjust the ratio of resuspension versus initial emission source. In the present study, the air change rate was lower than the particle deposition rate. Therefore, more than half of the nebulized viral particle mass settled on room surfaces and was available for resuspension. In real environments, with more complex human emissions, activities, and cleaning patterns, the viral loading on surfaces would not be as directly linked to the initial viral emissions and would be more difficult to predict.
Previous research has investigated the influence of environmental and other parameters on resuspension phenomena (Qian et al. (2014)) [
6]. For most studies, higher RH levels have led to reduced resuspension rates. However, the relationship between RH and resuspension is complex due to the competing effects of capillary forces and electrostatic forces. While our study did not report infectivity, RH, temperature, lighting, and surface type would also impact the infectivity of the virus [
5].
Existing resuspension models can be applied to determine the resuspension rate of settled particles due to human walking. However, the parameter values for the models vary widely [
6,
7] and parameter values specifically for viruses have not been determined [
48]. Our model using parameter values from the literature resulted in lower resuspension rates than what we observed. The resulting resuspension emission factor was similar to that reported in a previous study for 1–2
m [
47] non-biological particles.
In this study, we did not determine the particle size range associated with the resuspensed virus. We expect that the resuspended virus is associated with larger particles than the virus (≈0.1
m) due to the size distribution of the nebulized aerosol (
to 5
m) and generated test dust (
to 10
m). Based on theory, the larger particles would be easier to resuspend than smaller particles for the particle size range relevant to this study due to their higher ratio of the applied external detachment force to the adhesive force. Furthermore, mineral particles, such as those comprising the test dust, would be easier to resuspend than the liquid aerosol particles generated by the nebulizer due to the higher surface roughness, smaller contact area with the surface, and lack of a meniscus between the particle and the surface [
19]. Accordingly, the study results indicate increased viral resuspension for the dusty surface condition compared to the clean surface condition, although these results should be considered preliminary.
Currently, there is a large scientific knowledge gap concerning the resuspension of viral particles as well as non-biological particles smaller than
m. Most previous experimental resuspension studies have focused on particles larger than
m in diameter due to the aerosol instrumentation typically applied in these studies, which use either optical or time-of-flight measurement principles [
6,
40,
46,
49]. Only one study to date has experimentally determined resuspension rates for particles smaller than
m [
50]. It is unknown which size fraction the resuspended viral particles are most associated with given that the virus particles are contained within liquid respiratory aerosols and the deposited particles may land on existing surface dust. Viral resuspension studies that include size-resolved collection are recommended to address this knowledge gap.
Unfortunately, we were not able to determine the viability of the virus for this study. Various factors influence the recovery of viable viruses, including collection methods, flow rates, sampling times, and sampling media [
51,
52]. At the time we designed the study, the viral resuspension rates were unknown, but expected to be low. Therefore, to ensure that we had sufficient viral load to quantify the resuspended virus using qPCR analysis, we ran the impinger samplers for the full 5 h decay period. With the long sampling time and relatively high flow rate, none of the collected virus was viable. However, given the resuspension estimates obtained from this study and improved methods for collecting and measuring viral infectivity [
53,
54], one could reduce the viral sampling time to a fraction of the decay period and obtain estimates for the viable resuspended viruses.
Previous work has shown that viruses can survive in aerosols for as long as three hours. A study conducted by Bean et al. (1982) [
55] reported that influenza A and B viruses survived longer on hard surfaces such as stainless steel and plastic (24–48 h) compared to cloth, paper, and tissues (<8–12 h), whereas influenza A virus was detected on hands from contact with stainless steel for 24 h and tissue for up to 15 min. A laboratory study conducted with SARS-CoV-2 found that the virus could stay viable in aerosols for as long as 3 h, on plastic and stainless steel surfaces for up to 3 days, on copper surfaces for up to 4 h, and on cardboard surfaces for up to a day [
27]. Similarly, Fears et al. (2020) [
56] presented a study that highlighted that laboratory conditions had enabled SARS-CoV-2 to remain viable and infectious within aerosols for up to 16 h. Riddle et al. (2020) [
57] found viable SARS-CoV-2 virus 28 days after inoculation on glass, polymer, stainless steel, vinyl, and paper (all non-porous surfaces stored in the dark at 20 °C). These results suggest that the SARS-CoV-2 virus could be stable enough for resuspension to be a viable exposure pathway.
During the resuspension activity, some of the resuspended viruses may have originated from clothing and surfaces other than the flooring. However, we anticipate that the contribution of these sources was minimal. The investigator wore clean Tyvek coveralls and booties to minimize contributions from clothing, and the human activity (walking) specifically targeted the flooring. In a previous chamber study with human participants, Bhangar et al. (2016) [
44] reported that flooring accounted for two-thirds or more of the resuspended bioaerosol emissions.
Resuspension of viral particles from clothing could be a more important exposure pathway than resuspension from flooring during human activity due to a higher intake fraction (fraction of emitted particles that are inhaled). There is evidence from a study in a hospital environment that resuspension of SARS-CoV-2 from indoor surfaces and clothing may be an important exposure pathway due to the relatively high virus concentrations found in the hospital changing rooms and bathrooms [
33]. Ren et al. (2022) [
58] demonstrated that clothing effectively transports and resuspends bioaerosols. In an experimental study using fluorescent particles as tracers, they found almost
of the fluorescent particles deposited on clothing resuspended into the room. Licina et al. (2017) [
59] found that the emission rate is lower but the intake fraction is substantially higher for particles resuspended from seated movements than from walking indoors. This makes sense given the closer proximity of the breathing zone to the clothing. Furthermore, viruses on the clothing may be fresher and more viable than those on indoor surfaces, although this has not been studied. The issue of viral resuspension from both indoor surfaces and clothing requires further study.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








