Human Herpesvirus 6A Induces Dendritic Cell Death and HMGB1 Release without Virus Replication

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
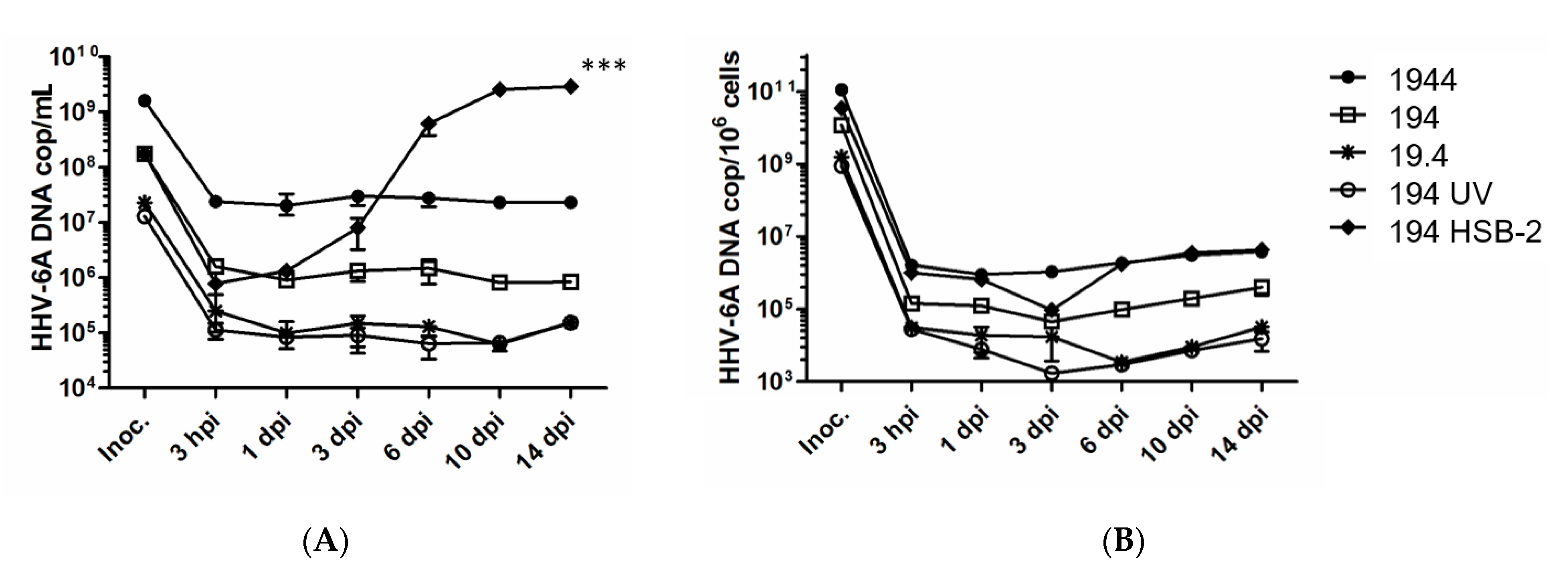
2.1. DC Do Not Support HHV-6A Viral Replication
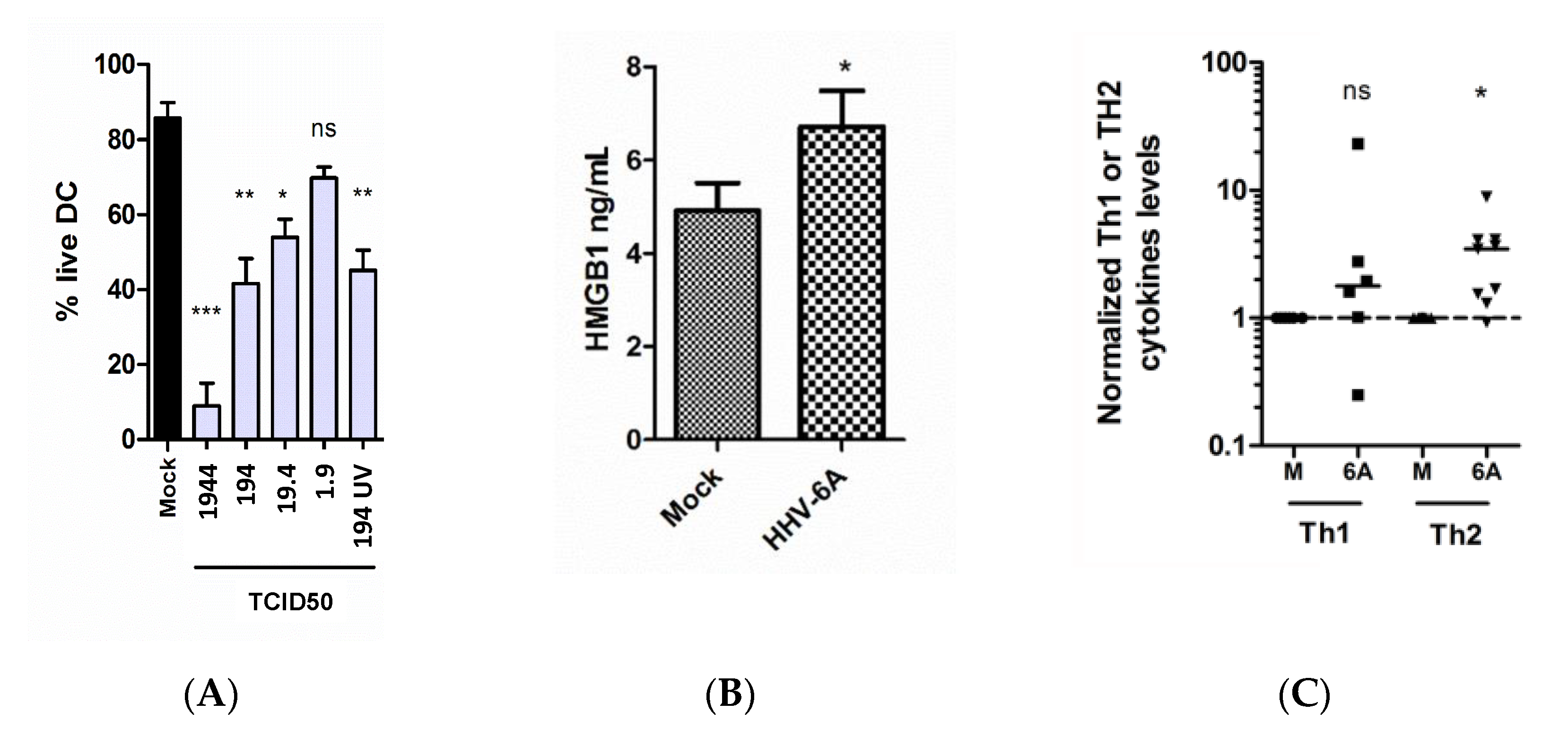
2.2. Exposure of HHV-6A by DC Induces Virus Titer-Dependent Cell Death, HMGB1 Release and Th2 Skewing
3. Discussion
4. Materials and Methods
4.1. Isolation of Peripheral Blood Monocytes and Generation of DC
4.2. HHV-6A Propagation and Inoculation
4.3. Assessment of HHV-6A Replication in DC
4.4. Assessment of HMGB1 Release
4.5. Co-Culture of DC and Allogenic T Cells and Cytokine Measurements
4.6. Statistics
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salahuddin, S.Z.; Ablashi, D.V.; Markham, P.D.; Josephs, S.F.; Sturzenegger, S.; Kaplan, M.; Halligan, G.; Biberfeld, P.; Wong-Staal, F.; Kramarsky, B. Isolation of a new virus, HBLV, in patients with lymphoproliferative disorders. Science 1986, 234, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Okuno, T.; Takahashi, K.; Balachandra, K.; Shiraki, K.; Yamanishi, K.; Takahashi, M.; Baba, K. Seroepidemiology of human herpesvirus 6 infection in normal children and adults. J. Clin. Microbiol. 1989, 27, 651–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerr, D.M.; Meier, A.S.; Selke, S.S.; Frenkel, L.M.; Huang, M.L.; Wald, A.; Rhoads, M.P.; Nguy, L.; Bornemann, R.; Morrow, R.A.; et al. A population-based study of primary human herpesvirus 6 infection. N. Engl. J. Med. 2005, 352, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Ablashi, D.; Agut, H.; Alvarez-Lafuente, R.; Clark, D.A.; Dewhurst, S.; DiLuca, D.; Flamand, L.; Frenkel, N.; Gallo, R.; Gompels, U.A.; et al. Classification of HHV-6A and HHV-6B as distinct viruses. Arch. Virol. 2014, 159, 863–870. [Google Scholar] [CrossRef]
- Yamanishi, K.; Okuno, T.; Shiraki, K.; Takahashi, M.; Kondo, T.; Asano, Y.; Kurata, T. Identification of human herpesvirus-6 as a causal agent for exanthem subitum. Lancet 1988, 1, 1065–1067. [Google Scholar] [CrossRef]
- Soldan, S.S.; Leist, T.P.; Juhng, K.N.; McFarland, H.F.; Jacobson, S. Increased lymphoproliferative response to human herpesvirus type 6A variant in multiple sclerosis patients. Ann. Neurol. 2000, 47, 306–313. [Google Scholar] [CrossRef]
- Akhyani, N.; Berti, R.; Brennan, M.B.; Soldan, S.S.; Eaton, J.M.; McFarland, H.F.; Jacobson, S. Tissue distribution and variant characterization of human herpesvirus (HHV)-6: Increased prevalence of HHV-6A in patients with multiple sclerosis. J. Infect. Dis. 2000, 182, 1321–1325. [Google Scholar] [CrossRef]
- Virtanen, J.O.; Färkkilä, M.; Multanen, J.; Uotila, L.; Jääskeläinen, A.J.; Vaheri, A.; Koskiniemi, M. Evidence for human herpesvirus 6 variant A antibodies in multiple sclerosis: Diagnostic and therapeutic implications. J. Neurovirol. 2007, 13, 347–352. [Google Scholar] [CrossRef]
- Engdahl, E.; Gustafsson, R.; Huang, J.; Biström, M.; Lima Bomfim, I.; Stridh, P.; Khademi, M.; Brenner, N.; Butt, J.; Michel, A.; et al. Increased Serological Response Against Human Herpesvirus 6A Is Associated with Risk for Multiple Sclerosis. Front. Immunol. 2019, 10, 2715. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.H.; Park, J.H.; Jee, M.H.; Keum, S.J.; Cho, M.S.; Yoon, S.K.; Jang, S.K. Hepatitis C virus infection is blocked by HMGB1 released from virus-infected cells. J. Virol. 2011, 85, 9359–9368. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wang, H.; Andersson, U. Targeting Inflammation Driven by HMGB1. Front. Immunol. 2020, 11, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, J.; Loh, Z.; Ullah, M.A.; Lynch, J.P.; Werder, R.B.; Collinson, N.; Zhang, V.; Dondelinger, Y.; Bertrand, M.J.M.; Everard, M.L.; et al. Respiratory Syncytial Virus Infection Promotes Necroptosis and HMGB1 Release by Airway Epithelial Cells. Am. J. Respir. Crit. Care Med. 2020, 201, 1358–1371. [Google Scholar] [CrossRef] [PubMed]
- Borde, C.; Barnay-Verdier, S.; Gaillard, C.; Hocini, H.; Maréchal, V.; Gozlan, J. Stepwise release of biologically active HMGB1 during HSV-2 infection. PLoS ONE 2011, 6, e16145. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, H.; Fujiyama, T.; Kanebayashi, J.; Kito, Y.; Hata, M.; Yagi, H. Skin recruitment of monomyeloid precursors involves human herpesvirus-6 reactivation in drug allergy. Allergy 2013, 68, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Fujino, Y.; Nakajima, M.; Inoue, H.; Kusuhara, T.; Yamada, T. Human herpesvirus 6 encephalitis associated with hypersensitivity syndrome. Ann. Neurol. 2002, 51, 771–774. [Google Scholar] [CrossRef]
- Descamps, V.; Ranger-Rogez, S. DRESS syndrome. Joint Bone Spine 2014, 81, 15–21. [Google Scholar] [CrossRef]
- Santoro, F.; Kennedy, P.E.; Locatelli, G.; Malnati, M.S.; Berger, E.A.; Lusso, P. CD46 is a cellular receptor for human herpesvirus 6. Cell 1999, 99, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, R.K.; Engdahl, E.E.; Hammarfjord, O.; Adikari, S.B.; Lourda, M.; Klingström, J.; Svensson, M.; Fogdell-Hahn, A. Human Herpesvirus 6A Partially Suppresses Functional Properties of DC without Viral Replication. PLoS ONE 2013, 8, e58122. [Google Scholar]
- Gustafsson, R.; Svensson, M.; Fogdell-Hahn, A. Modulatory effects on dendritic cells by human herpesvirus 6. Front. Microbiol. 2015, 6, 388. [Google Scholar] [CrossRef]
- Jaworska, J.; Gravel, A.; Flamand, L. Divergent susceptibilities of human herpesvirus 6 variants to type I interferons. Proc. Natl. Acad. Sci. USA 2010, 107, 8369–8374. [Google Scholar] [CrossRef] [Green Version]
- Hafler, D.A.; Slavik, J.M.; Anderson, D.E.; O’Connor, K.C.; De Jager, P.; Baecher-Allan, C. Multiple sclerosis. Immunol. Rev. 2005, 204, 208–231. [Google Scholar] [CrossRef] [PubMed]
- Beecham, A.H.; Patsopoulos, N.A.; Xifara, D.K.; Davis, M.F.; Kemppinen, A.; Cotsapas, C.; Shah, T.S.; Spencer, C.; Booth, D.; Goris, A.; et al. Analysis of immune-related loci identifies 48 new susceptibility variants for multiple sclerosis. Nat. Genet. 2013, 45, 1353–1360. [Google Scholar] [PubMed]
- Sawcer, S.; Hellenthal, G.; Pirinen, M.; Spencer, C.C.; Patsopoulos, N.A.; Moutsianas, L.; Dilthey, A.; Su, Z.; Freeman, C.; Hunt, S.E.; et al. Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature 2011, 476, 214–219. [Google Scholar] [PubMed]
- Murdaca, G.; Tonacci, A.; Negrini, S.; Greco, M.; Borro, M.; Puppo, F.; Gangemi, S. Emerging role of vitamin D in autoimmune diseases: An update on evidence and therapeutic implications. Autoimmun. Rev. 2019, 18, 102350. [Google Scholar] [CrossRef]
- Hedström, A.K.; Olsson, T.; Alfredsson, L. The Role of Environment and Lifestyle in Determining the Risk of Multiple Sclerosis. Curr. Top. Behav. Neurosci. 2015, 26, 87–104. [Google Scholar] [CrossRef]
- Paudel, Y.N.; Angelopoulou, E.; Bhuvan, K.C.; Piperi, C.; Othman, I. High mobility group box 1 (HMGB1) protein in Multiple Sclerosis (MS): Mechanisms and therapeutic potential. Life Sci. 2019, 238, 116924. [Google Scholar] [CrossRef]
- Andersson, A.; Covacu, R.; Sunnemark, D.; Danilov, A.I.; Dal Bianco, A.; Khademi, M.; Wallström, E.; Lobell, A.; Brundin, L.; Lassmann, H.; et al. Pivotal advance: HMGB1 expression in active lesions of human and experimental multiple sclerosis. J. Leukoc. Biol. 2008, 84, 1248–1255. [Google Scholar] [CrossRef] [Green Version]
- Sternberg, Z.; Sternberg, D.; Chichelli, T.; Drake, A.; Patel, N.; Kolb, C.; Chadha, K.; Yu, J.; Hojnacki, D. High-mobility group box 1 in multiple sclerosis. Immunol. Res. 2016, 64, 385–391. [Google Scholar] [CrossRef]
- Gustafsson, R.K.; Engdahl, E.E.; Fogdell-Hahn, A. Development and validation of a Q-PCR based TCID50 method for human herpesvirus 6. Virol. J. 2012, 9, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitsche, A.; Müller, C.W.; Radonic, A.; Landt, O.; Ellerbrok, H.; Pauli, G.; Siegert, W. Human herpesvirus 6A DNA Is detected frequently in plasma but rarely in peripheral blood leukocytes of patients after bone marrow transplantation. J. Infect. Dis. 2001, 183, 130–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gustafsson, R. Human Herpesvirus 6A Induces Dendritic Cell Death and HMGB1 Release without Virus Replication. Pathogens 2021, 10, 57. https://doi.org/10.3390/pathogens10010057
Gustafsson R. Human Herpesvirus 6A Induces Dendritic Cell Death and HMGB1 Release without Virus Replication. Pathogens. 2021; 10(1):57. https://doi.org/10.3390/pathogens10010057
Chicago/Turabian StyleGustafsson, Rasmus. 2021. "Human Herpesvirus 6A Induces Dendritic Cell Death and HMGB1 Release without Virus Replication" Pathogens 10, no. 1: 57. https://doi.org/10.3390/pathogens10010057
APA StyleGustafsson, R. (2021). Human Herpesvirus 6A Induces Dendritic Cell Death and HMGB1 Release without Virus Replication. Pathogens, 10(1), 57. https://doi.org/10.3390/pathogens10010057