
Babesia microti Immunoreactive Rhoptry-Associated Protein-1 Paralogs Are Ancestral Members of the Piroplasmid-Confined RAP-1 Family

 ,
, 
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
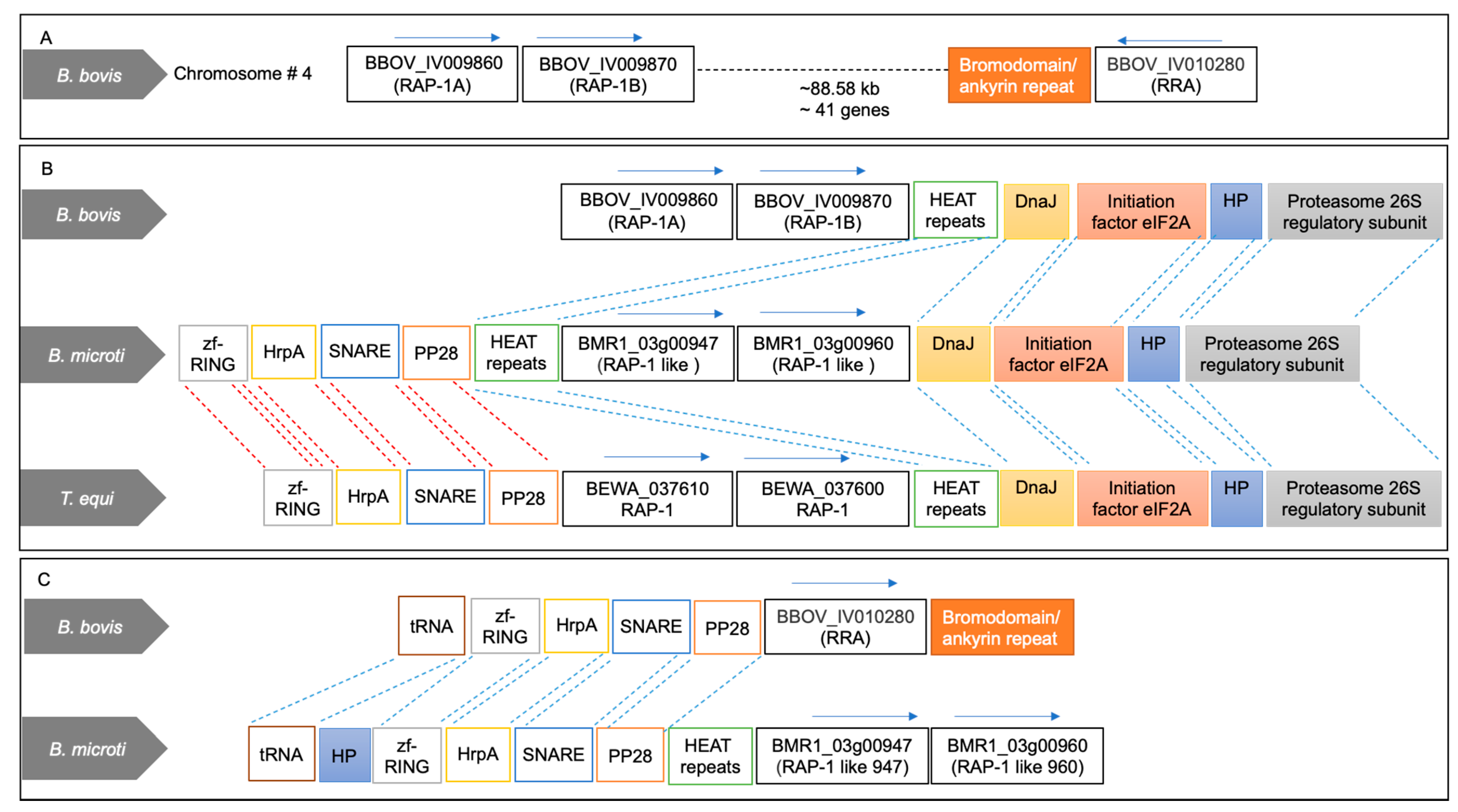
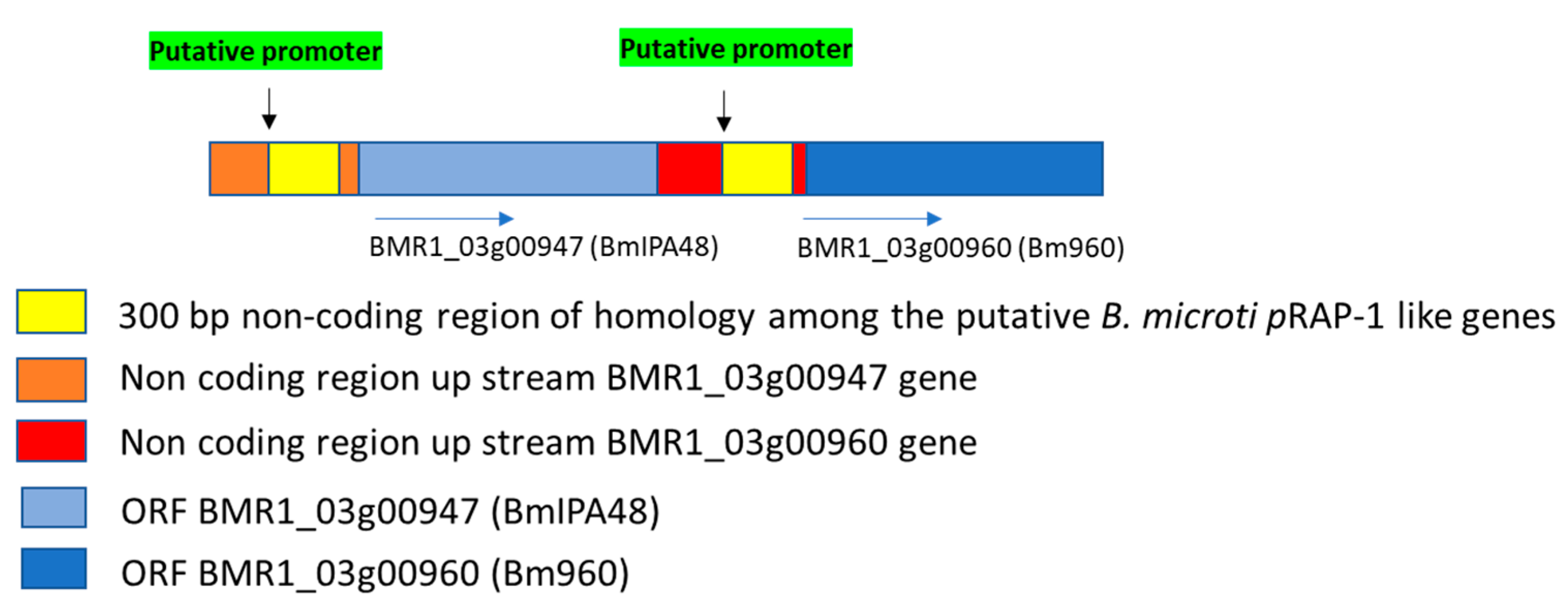
2.1. Two Tandemly Arranged RAP-1 Syntenic Genes of B. microti Encode Proteins Containing Non-Canonical Piroplasmid RAP-1 Cys-Rich Domains
2.2. Significance of Synteny Relationships among rap-1 and rra Genes of Babesia and Theileria
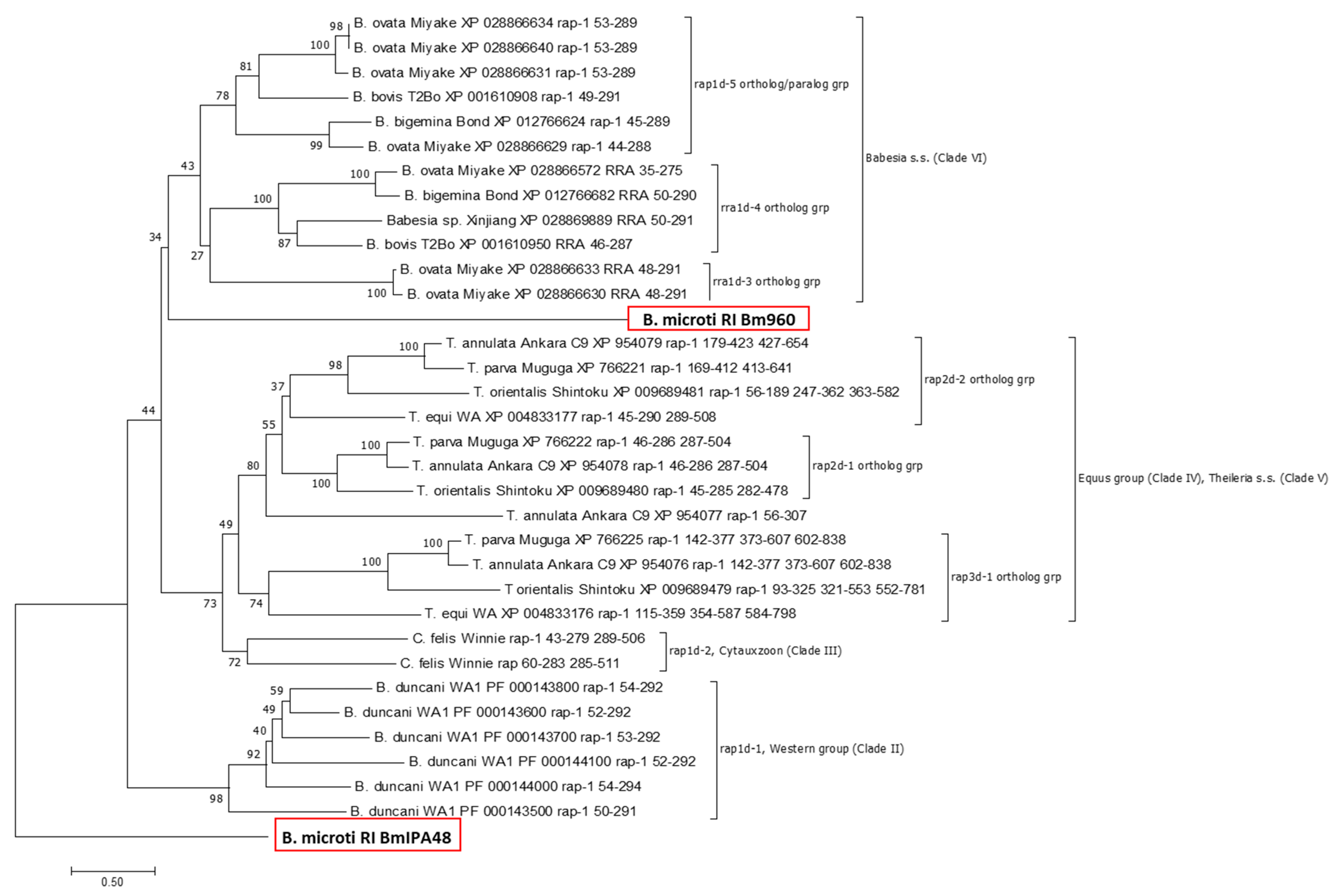
2.3. Phylogeny of Piroplasmid RAP-1 Proteins Recapitulates Piroplasmid Phylogeny
2.4. BmIPA48 and Bm960 Are Immunogenic during Infection in Humans
3. Discussion
4. Materials and Methods
4.1. Expression of Recombinant B. microti pRAP-1 like Proteins
4.2. Human Serum Samples
4.3. ELISA Procedure
4.4. Immunoblot Analysis
4.5. Bioinformatic Analysis
4.6. Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef]
- Jalovecka, M.; Hajdusek, O.; Sojka, D.; Kopacek, P.; Malandrin, L. The Complexity of Piroplasms Life Cycles. Front. Cell. Infect. Microbiol. 2018, 8, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalovecka, M.; Sojka, D.; Ascencio, M.; Schnittger, L. Babesia Life Cycle-When Phylogeny Meets Biology. Trends Parasitol. 2019, 35, 356–368. [Google Scholar] [CrossRef]
- Yang, Y.; Christie, J.; Köster, L.; Du, A.; Yao, C. Emerging Human Babesiosis with “Ground Zero” in North America. Microorganisms 2021, 9, 440. [Google Scholar] [CrossRef]
- Karshima, S.N.; Karshima, M.N.; Ahmed, M.I. Animal reservoirs of zoonotic Babesia species: A global systematic review and meta-analysis of their prevalence, distribution and species diversity. Vet. Parasitol. 2021, 298, 109539. [Google Scholar] [CrossRef] [PubMed]
- Lobo, C.A.; Singh, M.; Rodriguez, M. Human babesiosis: Recent advances and future challenges. Curr. Opin. Hematol. 2020, 27, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.T.M.; Tran, M.-H. Transfusion transmitted babesiosis: A systematic review of reported cases. Transfus. Apher. Sci. 2020, 59, 102843. [Google Scholar] [CrossRef] [PubMed]
- Alkishe, A.; Raghavan, R.; Peterson, A. Likely geographic distributional shifts among medically important tick species and tick-associated diseases under climate change in North America: A review. Insects 2021, 12, 225. [Google Scholar] [CrossRef]
- Tufts, D.M.; A Diuk-Wasser, M. Vertical Transmission: A vector-independent transmission pathway of Babesia microti in the natural reservoir host. Peromyscus. Leucopus. J. Infect. Dis. 2020, 223, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- White, D.J.; Talarico, J.; Chang, H.-G.; Birkhead, G.S.; Heimberger, T.; Morse, D.L. Human babesiosis in New York State. Arch. Intern. Med. 1998, 158, 2149–2154. [Google Scholar] [CrossRef]
- Gubbels, M.-J.; Duraisingh, M.T. Evolution of apicomplexan secretory organelles. Int. J. Parasitol. 2012, 42, 1071–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert-Paganin, J.; Xu, X.-P.; Swift, M.F.; Auguin, D.; Robblee, J.P.; Lu, H.; Fagnant, P.M.; Krementsova, E.B.; Trybus, K.M.; Houdusse, A.; et al. The actomyosin interface contains an evolutionary conserved core and an ancillary interface involved in specificity. Nat. Commun. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Ben Chaabene, R.; Lentini, G.; Soldati-Favre, D. Biogenesis and discharge of the rhoptries: Key organelles for entry and hijack of host cells by the Apicomplexa. Mol. Microbiol. 2020, 115, 453–465. [Google Scholar] [CrossRef]
- Thekkiniath, J.; Kilian, N.; Lawres, L.; A Gewirtz, M.; Graham, M.M.; Liu, X.; Ledizet, M.; Ben Mamoun, C. Evidence for vesicle-mediated antigen export by the human pathogen. Babesia Microti. Life Sci. Alliance. 2019, 2, e201900382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, C.E.; Palmer, G.H.; Hotzel, I.; McElwain, T.F. Structure, sequence, and transcriptional analysis of the Babesia bovis rap-1 multigene locus. Mol. Biochem. Parasitol. 1998, 93, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Kappmeyer, L.; Perryman, L.E.; Hines, S.A.; Baszler, T.V.; Katz, J.B.; Hennager, S.G.; Knowles, D.P. Detection of equine antibodies to Babesia caballi by recombinant B. caballi rhoptry-associated protein 1 in a competitive-inhibition enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1999, 37, 2285–2290. [Google Scholar] [CrossRef] [Green Version]
- Suarez, C.E.; Palmer, G.H.; Florin-Christensen, M.; Hines, S.A.; Hötzel, I.; McElwain, T.F. Organization, transcription, and expression of rhoptry associated protein genes in the Babesia bigemina rap-1 locus. Mol. Biochem. Parasitol. 2003, 127, 101–112. [Google Scholar] [CrossRef]
- Zhou, J.; Jia, H.; Nishikawa, Y.; Fujisaki, K.; Xuan, X. Babesia gibsoni rhoptry-associated protein 1 and its potential use as a diagnostic antigen. Vet. Parasitol. 2007, 145, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Terkawi, M.A.; Amornthep, A.; Ooka, H.; Aboge, G.; Jia, H.; Goo, Y.-K.; Nelson, B.; Yamagishi, J.; Nishikawa, Y.; Igarashi, I.; et al. Molecular characterizations of three distinct Babesia gibsoni rhoptry-associated protein-1s (RAP-1s). Parasitology 2009, 136, 1147–1160. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; Alhassan, A.; Ord, R.L.; Cursino-Santos, J.R.; Singh, M.; Gray, J.; Lobo, C.A. Identification and characterization of the RouenBd1987 Babesia divergens rhopty-associated protein 1. PLoS ONE 2014, 9, e107727. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; He, L.; Zhang, W.-J.; Cheng, J.-X.; Hu, J.-F.; Miao, X.-Y.; Huang, Y.; Fan, L.-Z.; Khan, M.K.; Zhou, Y.-Q.; et al. Molecular cloning and characterization of Babesia orientalis rhoptry-associated protein 1. Vet. Parasitol. 2014, 205, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Bonsergent, C.; Guan, G.; Yin, H.; Malandrin, L. Sequence and organization of the rhoptry-associated-protein-1 (rap-1) locus for the sheep hemoprotozoan Babesia sp. BQ1 Lintan (B. motasi phylogenetic group). Vet. Parasitol. 2013, 198, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Marchand, J.; Yang, C.; Bonsergent, C.; Guan, G.; Yin, H.; Malandrin, L. Rhoptry-associated protein (rap-1) genes in the sheep pathogen Babesia sp. Xinjiang: Multiple transcribed copies differing by 3′ end repeated sequences. Vet. Parasitol. 2015, 211, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.K. Cell invasion by Theileria sporozoites. Trends Parasitol. 2003, 19, 2–6. [Google Scholar] [CrossRef]
- Yokoyama, N.; Suthisak, B.; Hirata, H.; Matsuo, T.; Inoue, N.; Sugimoto, C.; Igarashi, I. Cellular localization of Babesia bovis merozoite rhoptry-associated protein 1 and its erythrocyte-binding activity. Infect. Immun. 2002, 70, 5822–5826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, C.E.; Laughery, J.M.; Bastos, R.G.; Johnson, W.C.; Norimine, J.; Asenzo, G.; Brown, W.C.; Florin-Christensen, M.; Goff, W.L. A novel neutralization sensitive and subdominant RAP-1-related antigen (RRA) is expressed by Babesia bovis merozoites. Parasitology 2011, 138, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Moreno, R.; Pöltl-Frank, F.; Stüber, D.; Matile, H.; Mutz, M.; Weiss, N.A.; Pluschke, G. Rhoptry-associated protein 1-binding monoclonal antibody raised against a heterologous peptide sequence inhibits Plasmodium falciparum growth in vitro. Infect. Immun. 2001, 69, 2558–2568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushe, T.C.; Palmer, G.H.; Sotomayor, L.; Figueroa, J.V.; Buening, G.M.; E Perryman, L.; McElwain, T.F. Antibody response to a Babesia bigemina rhoptry-associated protein 1 surface-exposed and neutralization-sensitive epitope in immune cattle. Infect. Immun. 1994, 62, 5698–5701. [Google Scholar] [CrossRef] [Green Version]
- Mosqueda, J.; McElwain, T.F.; Stiller, D.; Palmer, G.H. Babesia bovis Merozoite surface antigen 1 and rhoptry-associated protein 1 are expressed in sporozoites, and specific antibodies inhibit sporozoite attachment to erythrocytes. Infect. Immun. 2002, 70, 1599–1603. [Google Scholar] [CrossRef] [Green Version]
- Norimine, J.; Suarez, C.E.; McElwain, T.F.; Florin-Christensen, M.; Brown, W.C. Immunodominant epitopes in Babesia bovis rhoptry-associated protein 1 that elicit memory CD4(+)-T-lymphocyte responses in B. bovis-immune individuals are located in the amino-terminal domain. Infect. Immun. 2002, 70, 2039–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonchit, S.; Xuan, X.; Yokoyama, N.; Goff, W.L.; Waghela, S.D.; Wagner, G.; Igarashi, I. Improved enzyme-linked immunosorbent assay using c-terminal truncated recombinant antigens of Babesia bovis rhoptry-associated protein-1 for detection of specific antibodies. J. Clin. Microbiol. 2004, 42, 1601–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonchit, S.; Alhassan, A.; Chan, B.; Xuan, X.; Yokoyama, N.; Ooshiro, M.; Goff, W.L.; Waghela, S.D.; Wagner, G.; Igarashi, I. Expression of C-terminal truncated and full-length Babesia bigemina rhoptry-associated protein 1 and their potential use in enzyme-linked immunosorbent assay. Vet. Parasitol. 2006, 137, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Suarez, C.E.; Noh, S. Emerging perspectives in the research of bovine babesiosis and anaplasmosis. Vet. Parasitol. 2011, 180, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Liu, Z.; Yang, J.; Yu, P.; Pan, Y.; Zhai, B.; Luo, J.; Moreau, E.; Guan, G.; Yin, H. Expression analysis and biological characterization of Babesia sp. BQ1 (Lintan) (Babesia motasi-like) rhoptry-associated protein 1 and its potential use in serodiagnosis via ELISA. Parasites Vectors. 2016, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Cornillot, E.; McCracken, C.; Usmani-Brown, S.; Dwivedi, A.; Ifeonu, O.O.; Crabtree, J.; Gotia, H.T.; Virji, A.Z.; Reynes, C.; et al. Genome-wide diversity and gene expression profiling of Babesia microti isolates identify polymorphic genes that mediate host-pathogen interactions. Sci. Rep. 2016, 6, 35284. [Google Scholar] [CrossRef] [PubMed]
- Magni, R.; Luchini, A.; Liotta, L.; Molestina, R.E. Analysis of the Babesia microti proteome in infected red blood cells by a combination of nanotechnology and mass spectrometry. Int. J. Parasitol. 2018, 49, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Magni, R.; Luchini, A.; Liotta, L.; Molestina, R.E. Proteomic analysis reveals pathogen-derived biomarkers of acute babesiosis in erythrocytes, plasma, and urine of infected hamsters. Parasitol. Res. 2020, 119, 2227–2235. [Google Scholar] [CrossRef]
- Moreno-Hagelsieb, G.; Trevino, V.; Perez-Rueda, E.; Smith, T.F.; Collado-Vides, J. Transcription unit conservation in the three domains of life: A perspective from Escherichia coli. Trends Genet. 2001, 17, 175–177. [Google Scholar] [CrossRef]
- Florin-Christensen, M.; Suarez, C.E.; Hines, S.A.; Palmer, G.H.; Brown, W.C.; McElwain, T.F. The Babesia bovis merozoite surface antigen 2 locus contains four tandemly arranged and expressed genes encoding immunologically distinct proteins. Infect. Immun. 2002, 70, 3566–3575. [Google Scholar] [CrossRef] [Green Version]
- Suarez, C.E.; Norimine, J.; Lacy, P.; McElwain, T.F. Characterization and gene expression of Babesia bovis elongation factor-1α. Int. J. Parasitol. 2006, 36, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.G.; Knowles, D.P.; Mazuz, M.L.; Cooke, B.M.; Suarez, C.E. Stable transformation of Babesia bigemina and Babesia bovis using a single transfection plasmid. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Suarez, C.E.; Palmer, G.H.; Jasmer, D.P.; Hines, S.A.; Perryman, L.E.; McElwain, T.F. Characterization of the gene encoding a 60-kilodalton Babesia bovis merozoite protein with conserved and surface exposed epitopes. Mol. Biochem. Parasitol. 1991, 46, 45–52. [Google Scholar] [CrossRef]
- Paing, M.M.; Tolia, N.H. multimeric assembly of host-pathogen adhesion complexes involved in apicomplexan invasion. PLOS Pathog. 2014, 10, e1004120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Bastos, R.G.; Franceschi, V.; Tebaldi, G.; Connelley, T.; Morrison, W.I.; Knowles, D.P.; Donofrio, G.; Fry, L.M. Molecular and antigenic properties of mammalian cell-expressed Theileria parva antigen Tp9. Front. Immunol. 2019, 10, 897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, A.P.; Otto, T.; Darby, A.; Ramaprasad, A.; Xia, D.; Echaide, I.E.; Farber, M.; Gahlot, S.; Gamble, J.; Gupta, D.; et al. The evolutionary dynamics of variant antigen genes in Babesia reveal a history of genomic innovation underlying host-parasite interaction. Nucleic Acids Res. 2014, 42, 7113–7131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Brayton, K.; Lau, A.; Herndon, D.R.; Hannick, L.; Kappmeyer, L.; Berens, S.J.; Bidwell, S.L.; Brown, W.C.; Crabtree, J.; Fadrosh, D.; et al. Genome sequence of Babesia bovis and comparative analysis of apicomplexan hemoprotozoa. PLoS Pathog. 2007, 3, e148. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, J.; Asada, M.; Hakimi, H.; Tanaka, T.Q.; Sugimoto, C.; Kawazu, S.-I. Whole-genome assembly of Babesia ovata and comparative genomics between closely related pathogens. BMC Genom. 2017, 18, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tarigo, J.L.; Scholl, E.H.; Bird, D.; Brown, C.C.; Cohn, L.A.; Dean, G.A.; Levy, M.G.; Doolan, D.L.; Trieu, A.; Nordone, S.K.; et al. A novel candidate vaccine for Cytauxzoonosis inferred from comparative apicomplexan genomics. PLoS ONE 2013, 8, e71233. [Google Scholar] [CrossRef]
- Pain, A.; Renauld, H.; Berriman, M.; Murphy, L.; Yeats, C.A.; Weir, W.; Kerhornou, A.; Aslett, M.; Bishop, R.; Bouchier, C.; et al. Genome of the host-cell transforming parasite Theileria annulata compared with T. parva. Science 2005, 309, 131–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappmeyer, L.S.; Thiagarajan, M.; Herndon, D.R.; Ramsay, J.D.; Caler, E.; Djikeng, A.; Gillespie, J.J.; Lau, A.O.; Roalson, E.H.; Silva, J.C.; et al. Comparative genomic analysis and phylogenetic position of Theileria equi. BMC Genom. 2012, 13, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashida, K.; Hara, Y.; Abe, T.; Yamasaki, C.; Toyoda, A.; Kosuge, T.; Suzuki, Y.; Sato, Y.; Kawashima, S.; Katayama, T.; et al. Comparative genome analysis of three eukaryotic parasites with differing abilities to transform leukocytes reveals key mediators of Theileria-induced leukocyte transformation. mBio 2012, 3, e00204-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, M.J.; Bishop, R.; Shah, T.; de Villiers, E.P.; Carlton, J.M.; Hall, N.; Ren, Q.; Paulsen, I.T.; Pain, A.; Berriman, M.; et al. Genome sequence of Theileria parva, a bovine pathogen that transforms lymphocytes. Science 2005, 309, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T.; Taylor, W.R.; Thornton, J. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (Reference Genome) | Clade (Sensu Schnittger et al. 2012) | rap-1 Domain Architectures | Number of rap-1 Paralogs | Nomenclature (Proposed) |
|---|---|---|---|---|
| B. microti | I (B. microti-group) |  | 2× | rap1d-like |
| B. duncani | II (Western group |  | 6× | rap1d-1 |
| C. felis | III (Cytauxzoon) |  | 2× | rap1d-2 |
| T. equi | IV (Equus group) |  | 1× 1× | rap2d-1 rap3d-1 |
| T. annulata T. parva T. orientalis | V (Theileria s.s.) |    | Ta:1×, Tp:1×,To:0 Ta:2×,Tp:2×,To:2× Ta:1×, Tp:1×,To:2× | rap2d-2 rap2d-1 rap3d-1 |
| B. bovis B. bigemina B. ovata Babesia sp. Xinjiang | VI (Babesia s.s.) |  | 3× 2× 8× 1× | rap1d-3 rap1d-4 rap1d-5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bastos, R.G.; Thekkiniath, J.; Ben Mamoun, C.; Fuller, L.; Molestina, R.E.; Florin-Christensen, M.; Schnittger, L.; Alzan, H.F.; Suarez, C.E. Babesia microti Immunoreactive Rhoptry-Associated Protein-1 Paralogs Are Ancestral Members of the Piroplasmid-Confined RAP-1 Family. Pathogens 2021, 10, 1384. https://doi.org/10.3390/pathogens10111384
Bastos RG, Thekkiniath J, Ben Mamoun C, Fuller L, Molestina RE, Florin-Christensen M, Schnittger L, Alzan HF, Suarez CE. Babesia microti Immunoreactive Rhoptry-Associated Protein-1 Paralogs Are Ancestral Members of the Piroplasmid-Confined RAP-1 Family. Pathogens. 2021; 10(11):1384. https://doi.org/10.3390/pathogens10111384
Chicago/Turabian StyleBastos, Reginaldo G., Jose Thekkiniath, Choukri Ben Mamoun, Lee Fuller, Robert E. Molestina, Monica Florin-Christensen, Leonhard Schnittger, Heba F. Alzan, and Carlos E. Suarez. 2021. "Babesia microti Immunoreactive Rhoptry-Associated Protein-1 Paralogs Are Ancestral Members of the Piroplasmid-Confined RAP-1 Family" Pathogens 10, no. 11: 1384. https://doi.org/10.3390/pathogens10111384
APA StyleBastos, R. G., Thekkiniath, J., Ben Mamoun, C., Fuller, L., Molestina, R. E., Florin-Christensen, M., Schnittger, L., Alzan, H. F., & Suarez, C. E. (2021). Babesia microti Immunoreactive Rhoptry-Associated Protein-1 Paralogs Are Ancestral Members of the Piroplasmid-Confined RAP-1 Family. Pathogens, 10(11), 1384. https://doi.org/10.3390/pathogens10111384