
Characterisation of Macrophage Polarisation in Mice Infected with Ninoa Strain of Trypanosoma cruzi



Abstract
:1. Introduction
2. Results
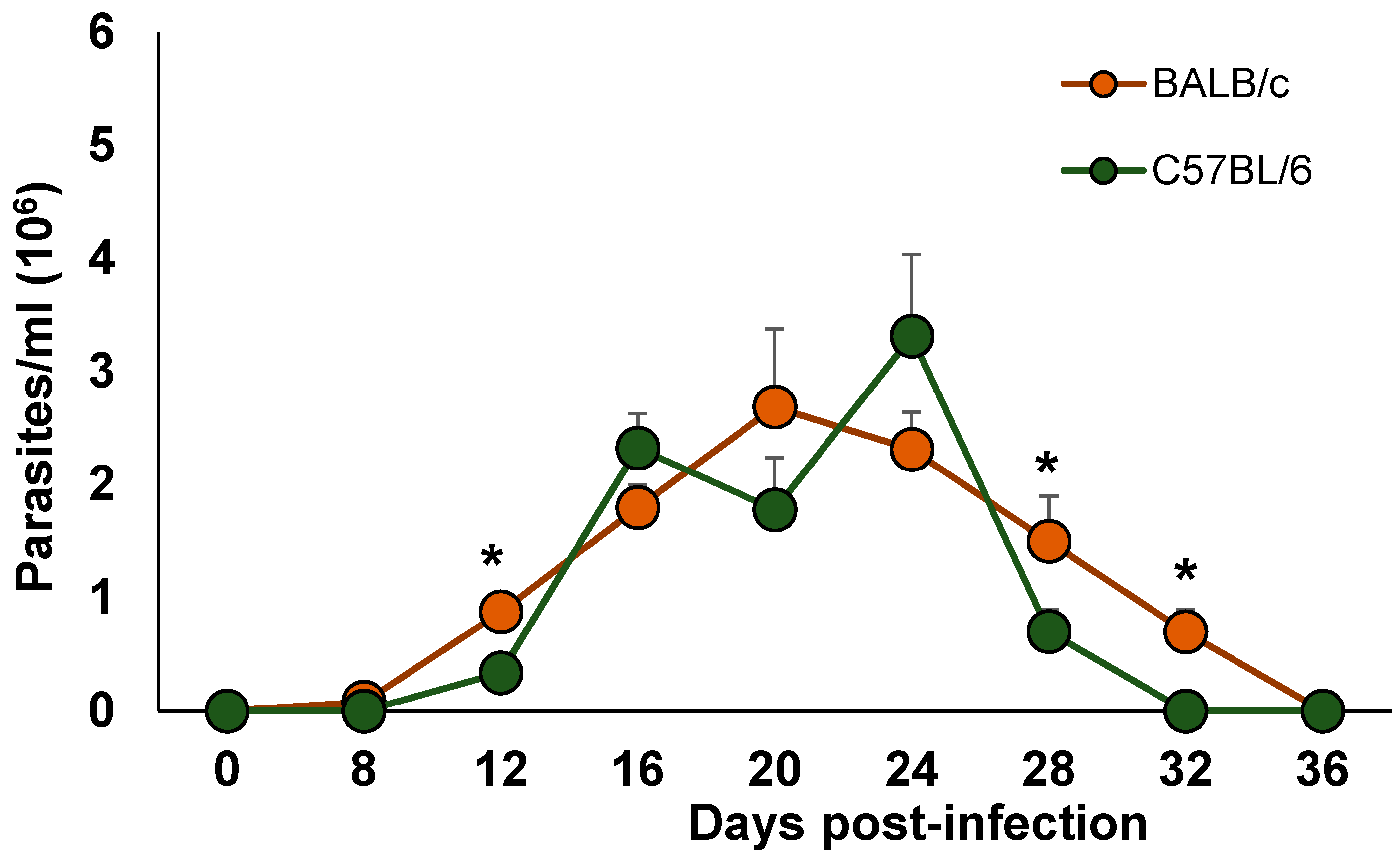
2.1. Parasitemia Burden in Blood of BALB/c and C57BL/6 Mice
2.2. Systemic Changes Due to T. cruzi Infection
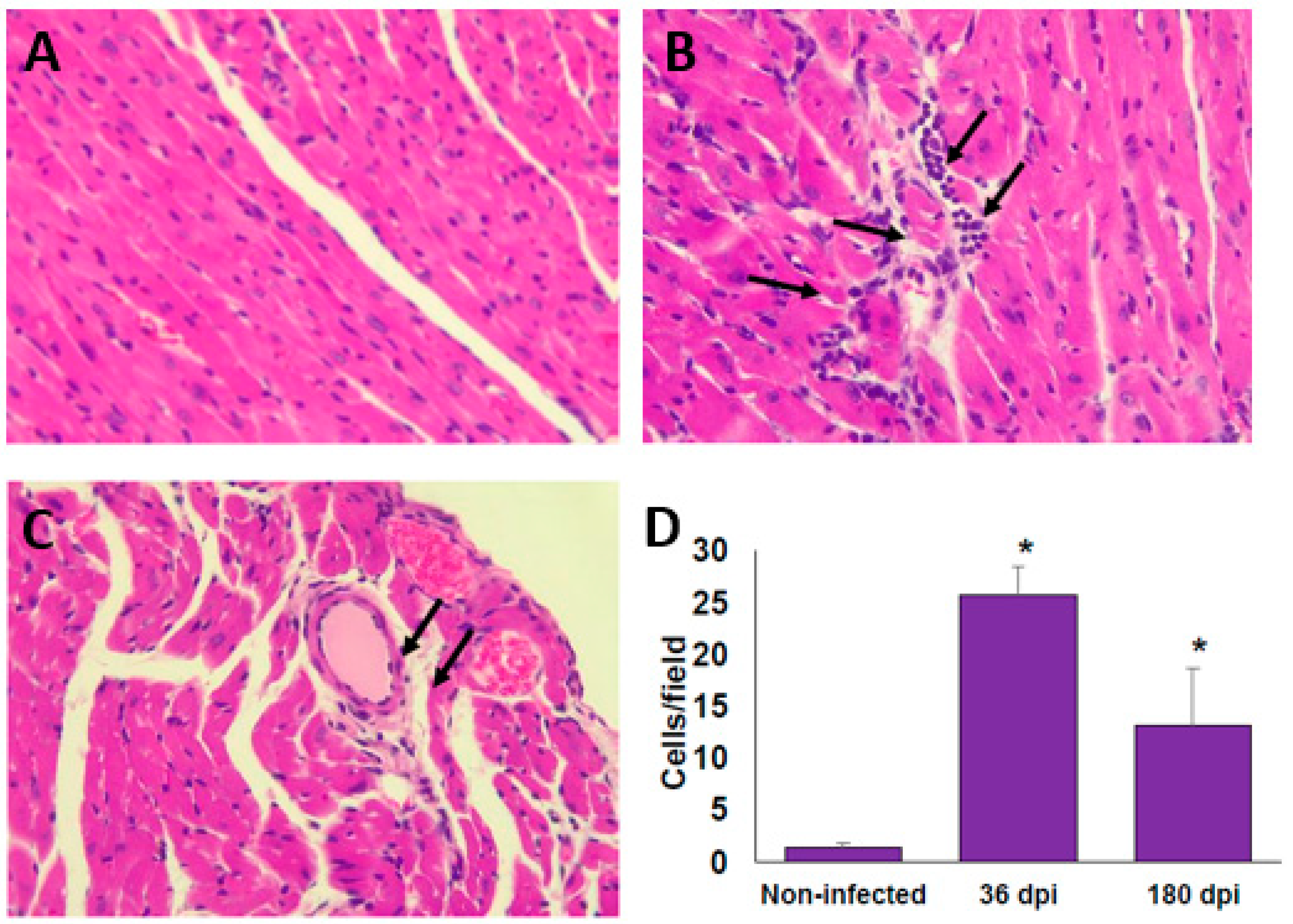
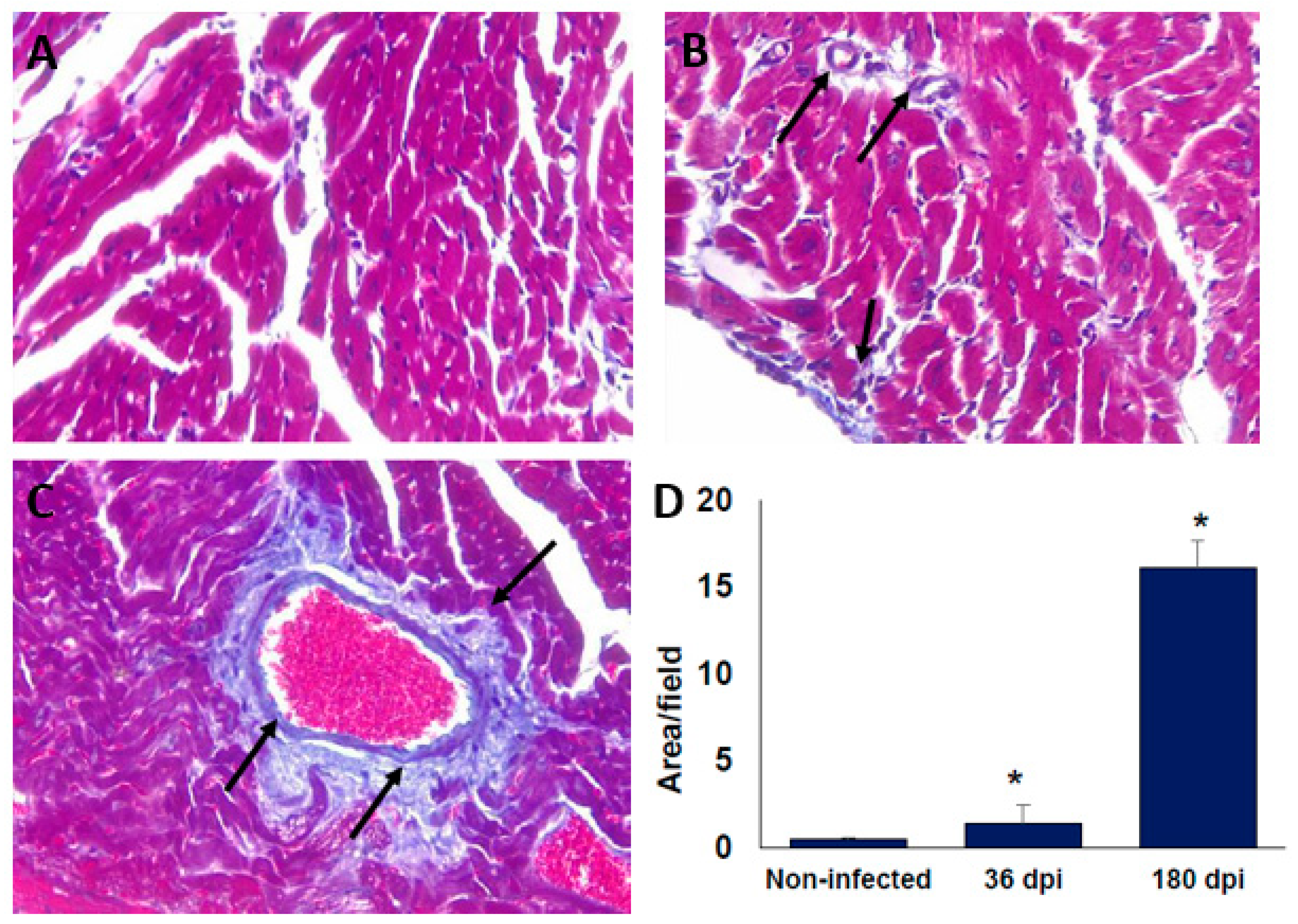
2.3. Histological Findings in the Heart
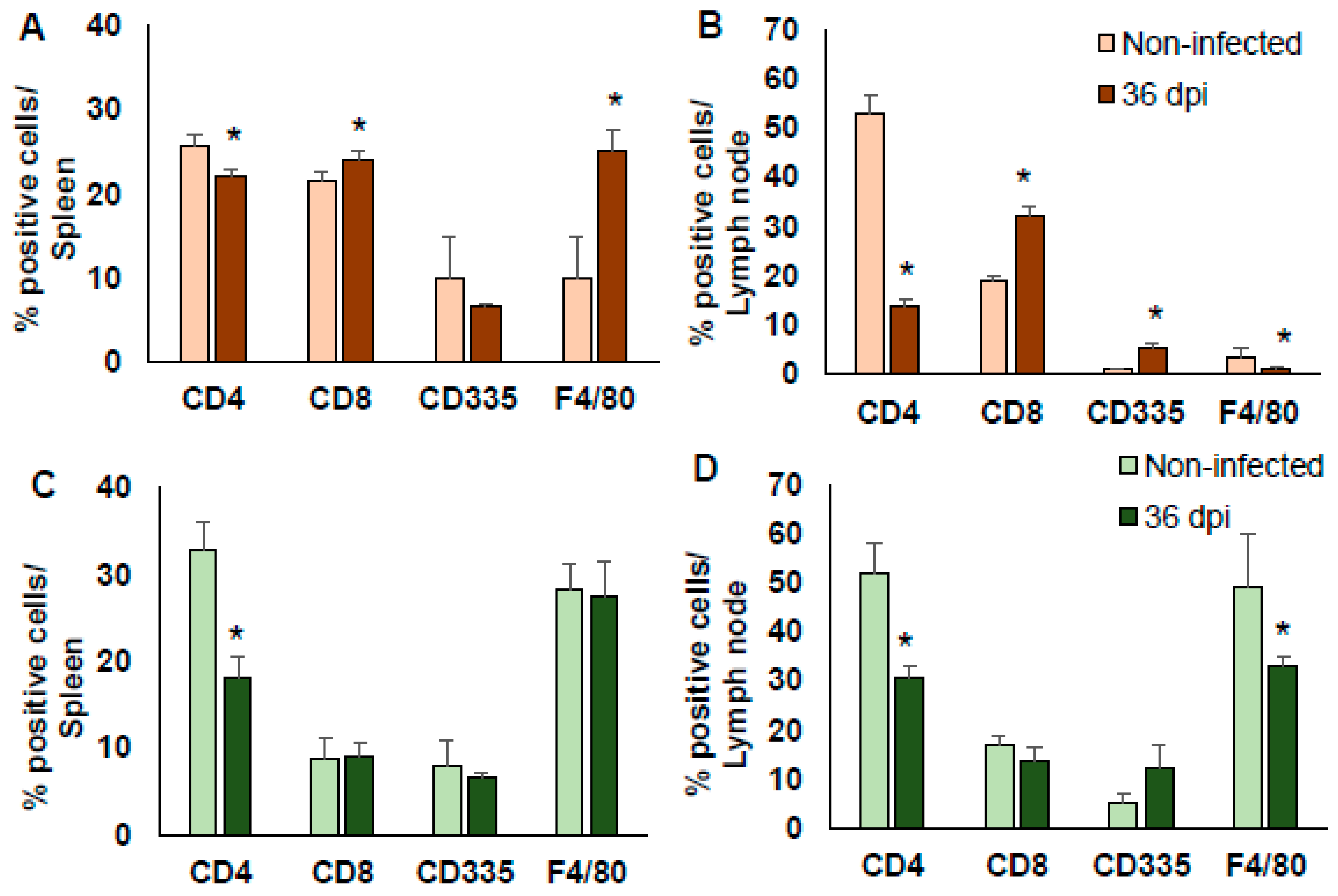
2.4. Cell Populations in Spleen and Lymph Nodes
2.5. Characterisation of Peritoneal Exudate Cells (PECs) after T. cruzi Infection
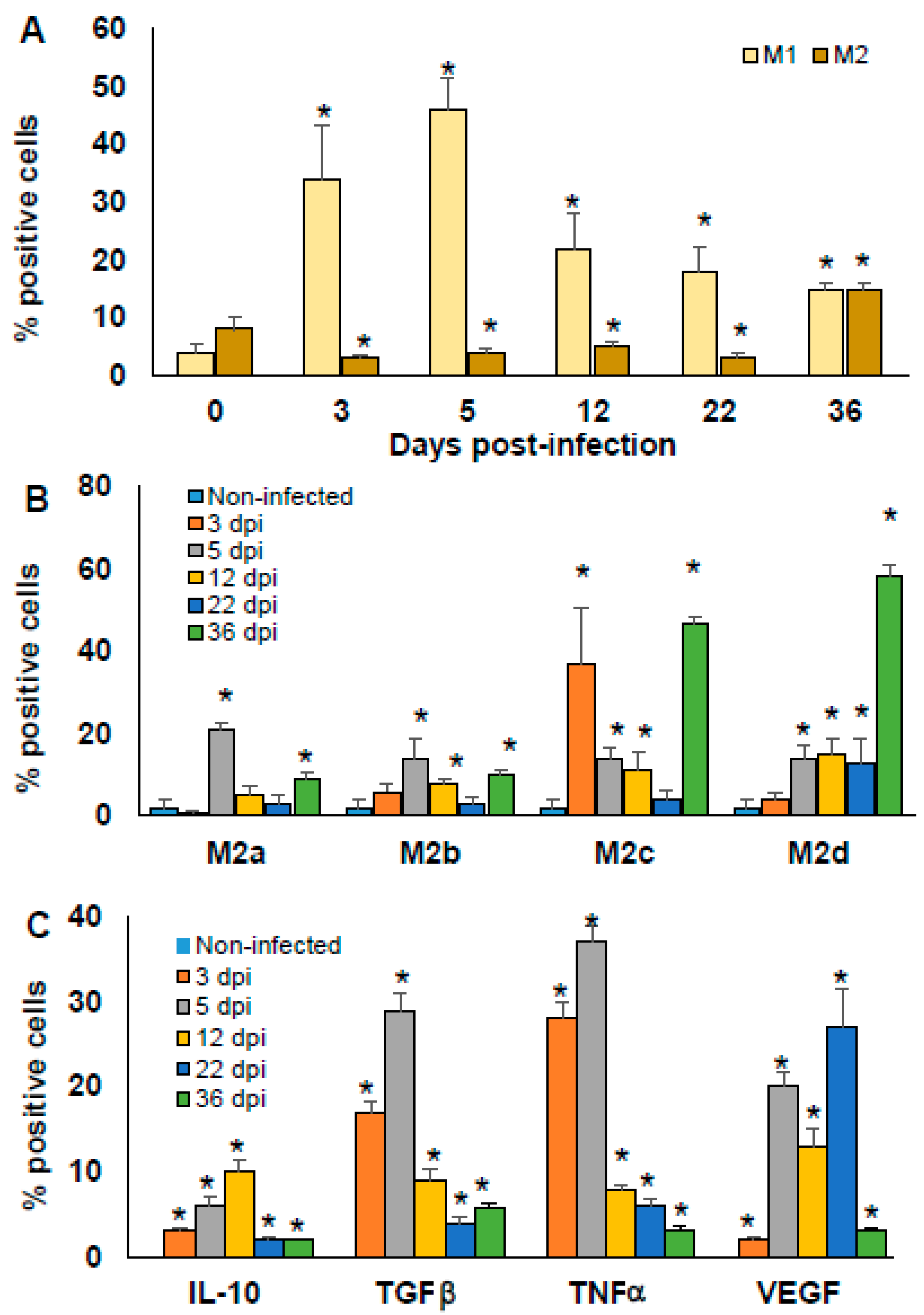
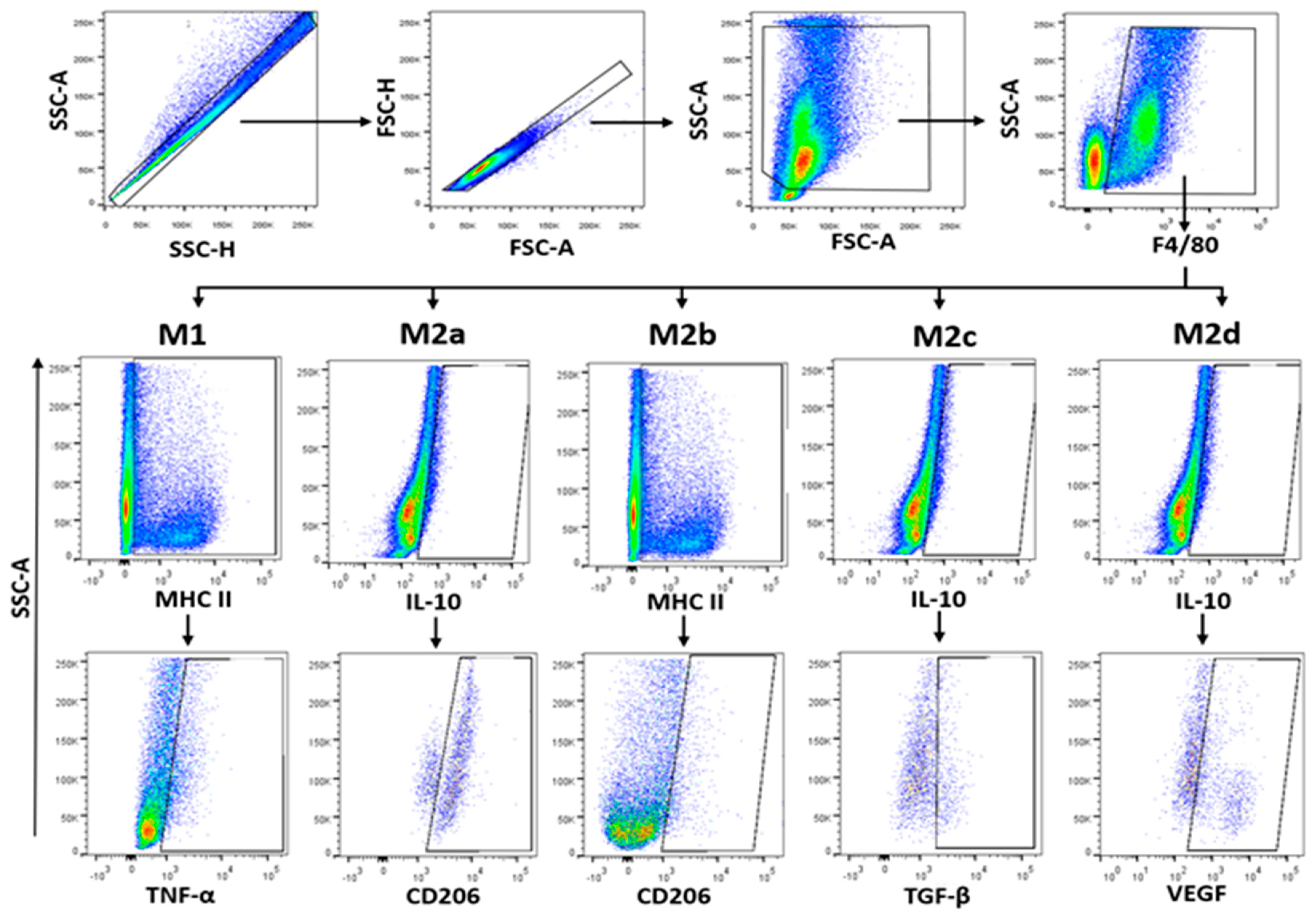
2.6. Macrophages Polarisation Induced by T. cruzi Infection
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Ethics Statement
4.3. Trypanosoma cruzi Strain
4.4. Experimental Groups and Infection Challenge
4.5. Histopathology
4.6. Single-Cell Preparation from Spleen and Inguinal Lymph Nodes
4.7. Cell Isolation from the Peritoneal Cavity
4.8. Flow Cytometry Staining
4.9. FACS Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hotez, P.J.; Molyneux, D.H.; Fenwick, A.; Kumaresan, J.; Sachs, S.E.; Sachs, J.D.; Savioli, L. Control of neglected tropical diseases. N. Engl. J. Med. 2007, 357, 1018–1027. [Google Scholar] [CrossRef] [Green Version]
- Mena-Marín, A.L.; Zeledón, R.; Morales, J.A.; Pereira, M.; Urbina, A. Sex influence on the susceptibility of Swiss mice to Trypanosoma cruzi [Influencia del sexo en la susceptibilidad de ratones Swiss a Trypanosoma cruzi]. Boletín de Malariogía y Salud Ambiental 2012, LII, 233–244. [Google Scholar]
- PAHO/WHO. Pan American Health Organization. Chagas Disease. Available online: https://www.paho.org/en/topics/chagas-disease (accessed on 6 May 2021).
- WHO. World Health Organization. Chagas Disease (American Trypanosomiasis). Available online: https://www.who.int/health-topics/chagas-disease#tab=tab_1 (accessed on 6 May 2021).
- Bern, C.; Martin, D.L.; Gilman, R.H. Acute and congenital Chagas disease. Adv. Parasitol. 2011, 75, 19–47. [Google Scholar]
- Lopes, M.F.; Freire-de-Lima, C.G.; DosReis, G.A. The macrophage haunted by cell ghosts: A pathogen grows. Immunol. Today 2000, 21, 489–494. [Google Scholar] [CrossRef]
- Rassi, A., Jr.; Rassi, A.; Marin-Neto, J.A. Chagas Disease. In Neglected Tropical Diseases—Latin America and the Caribbean; Neglected Tropical Diseases; Franco-Paredes, C., Santos-Preciado, J., Eds.; Springer: Vienna, Austria, 2015; pp. 45–71. [Google Scholar]
- Vieira, J.L.; Távora, F.R.F.; Sobral, M.G.V.; Vasconcelos, G.G.; Almeida, G.P.L.; Fernandes, J.R.; da Escóssia Marinho, L.L.; de Mendonça Trompieri, D.F.; Neto, J.D.D.S.; Mejia, J.A.C. Chagas cardiomyopathy in Latin America review. Curr. Cardiol. Rep. 2019, 21, 1–8. [Google Scholar] [CrossRef]
- Mills, R.M. Chagas Disease. Epidemiology and Barriers to Treatment. Am. J. Med. 2020, 133, 1262–1265. [Google Scholar] [CrossRef]
- Leon, C.M.; Montilla, M.; Vanegas, R.; Castillo, M.; Parra, E.; Ramirez, J.D. Murine models susceptibility to distinct Trypanosoma cruzi I genotypes infection. Parasitology 2017, 144, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Micucci, L.R.; Bazan, P.C.; Fauro, R.; Baez, A.; Lo Presti, M.S.; Triquel, M.F.; Paglini-Oliva, P.A.; Rivarola, H.W. [Importance of host sex in the development of Trypanosoma cruzi infection]. Rev. Fac. Cien. Med. Univ. Nac. Cordoba 2010, 67, 73–76. [Google Scholar]
- Rodriguez, T.; Pacheco-Fernandez, T.; Vazquez-Mendoza, A.; Nieto-Yanez, O.; Juarez-Avelar, I.; Reyes, J.L.; Terrazas, L.I.; Rodriguez-Sosa, M. MGL1 Receptor Plays a Key Role in the Control of T. cruzi Infection by Increasing Macrophage Activation through Modulation of ERK1/2, c-Jun, NF-kappaB and NLRP3 Pathways. Cells 2020, 9, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, A.; de Dios Ruiz-Rosado, J.; Terrazas, L.I.; Juarez, I.; Gomez-Garcia, L.; Calleja, E.; Camacho, G.; Chavez, A.; Romero, M.; Rodriguez, T.; et al. Mouse macrophage galactose-type lectin (mMGL) is critical for host resistance against Trypanosoma cruzi infection. Int. J. Biol. Sci. 2014, 10, 909–920. [Google Scholar] [CrossRef] [Green Version]
- Golgher, D.; Gazzinelli, R.T. Innate and acquired immunity in the pathogenesis of Chagas disease. Autoimmunity 2004, 37, 399–409. [Google Scholar] [CrossRef]
- Reyes, J.L.; Terrazas, L.I.; Espinoza, B.; Cruz-Robles, D.; Soto, V.; Rivera-Montoya, I.; Gomez-Garcia, L.; Snider, H.; Satoskar, A.R.; Rodriguez-Sosa, M. Macrophage migration inhibitory factor contributes to host defense against acute Trypanosoma cruzi infection. Infect. Immun. 2006, 74, 3170–3179. [Google Scholar] [CrossRef] [Green Version]
- Terrazas, C.A.; Huitron, E.; Vazquez, A.; Juarez, I.; Camacho, G.M.; Calleja, E.A.; Rodriguez-Sosa, M. MIF synergizes with Trypanosoma cruzi antigens to promote efficient dendritic cell maturation and IL-12 production via p38 MAPK. Int. J. Biol. Sci. 2011, 7, 1298–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, M.B.; Silva-Mota, K.N.; Lima, R.S.; Bellintani, M.C.; Pontes-de-Carvalho, L.; Ribeiro-dos-Santos, R. Modulation of chagasic cardiomyopathy by interleukin-4: Dissociation between inflammation and tissue parasitism. Am. J. Pathol. 2001, 159, 703–709. [Google Scholar] [CrossRef]
- da Silva, M.V.; de Almeida, V.L.; de Oliveira, W.D.; Matos Cascudo, N.C.; de Oliveira, P.G.; da Silva, C.A.; da Silva, A.C.S.; Dos Reis Monteiro, M.L.G.; Correa, R.R.M.; Oliveira, M.A.P.; et al. Upregulation of Cardiac IL-10 and Downregulation of IFN-gamma in Balb/c IL-4(−/−) in Acute Chagasic Myocarditis due to Colombian Strain of Trypanosoma cruzi. Mediat. Inflamm. 2018, 2018, 3421897. [Google Scholar] [CrossRef] [Green Version]
- Rios, L.E.; Vazquez-Chagoyan, J.C.; Pacheco, A.O.; Zago, M.P.; Garg, N.J. Immunity and vaccine development efforts against Trypanosoma cruzi. Acta Trop. 2019, 200, 105168. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, C.J.; Leibovich, S.J. Regulation of Macrophage Polarization and Wound Healing. Adv. Wound Care (New Rochelle) 2012, 1, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Tomiotto-Pellissier, F.; Bortoleti, B.; Assolini, J.P.; Goncalves, M.D.; Carloto, A.C.M.; Miranda-Sapla, M.M.; Conchon-Costa, I.; Bordignon, J.; Pavanelli, W.R. Macrophage Polarization in Leishmaniasis: Broadening Horizons. Front. Immunol. 2018, 9, 2529. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Cao, Q.; Zheng, D.; Sun, Y.; Wang, C.; Yu, X.; Wang, Y.; Lee, V.W.; Zheng, G.; Tan, T.K.; et al. Discrete functions of M2a and M2c macrophage subsets determine their relative efficacy in treating chronic kidney disease. Kidney Int. 2013, 84, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Zanluqui, N.G.; Lovo-Martins, M.I.; Malvezi, A.D.; Panis, C.; da Silva, R.V.; Tatakihara, V.L.H.; Felipe, I.; Martins-Pinge, M.C.; Wowk, P.F.; Pinge-Filho, P. Concanavalin-A stimulates IL-17 and nitric oxide production and induces macrophage polarization and resistance to Trypanosoma cruzi infection. Life Sci. 2020, 258, 118137. [Google Scholar] [CrossRef] [PubMed]
- Roffe, E.; Rothfuchs, A.G.; Santiago, H.C.; Marino, A.P.; Ribeiro-Gomes, F.L.; Eckhaus, M.; Antonelli, L.R.; Murphy, P.M. IL-10 limits parasite burden and protects against fatal myocarditis in a mouse model of Trypanosoma cruzi infection. J. Immunol. 2012, 188, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Fraternale, A.; Brundu, S.; Magnani, M. Polarization and repolarization of macrophages. J. Clin. Cell Immunol. 2015, 6, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Reyes, J.L.; Espinoza-Jimenez, A.F.; Gonzalez, M.I.; Verdin, L.; Terrazas, L.I. Taenia crassiceps infection abrogates experimental autoimmune encephalomyelitis. Cell. Immunol. 2011, 267, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Leal-Sena, J.A.; Dos Santos, J.L.; Dos Santos, T.A.R.; de Andrade, E.M.; de Oliveira Mendes, T.A.; Santana, J.O.; Mineo, T.W.P.; Mineo, J.R.; da Cunha-Junior, J.P.; Pirovani, C.P. Toxoplasma gondii antigen SAG2A differentially modulates IL-1beta expression in resistant and susceptible murine peritoneal cells. Appl. Microbiol. Biotechnol. 2018, 102, 2235–2249. [Google Scholar] [CrossRef]
- Espinoza, B.; Solorzano-Dominguez, N.; Vizcaino-Castillo, A.; Martinez, I.; Elias-Lopez, A.L.; Rodriguez-Martinez, J.A. Gastrointestinal infection with Mexican TcI Trypanosoma cruzi strains: Different degrees of colonization and diverse immune responses. Int. J. Biol. Sci. 2011, 7, 1357–1370. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, B.; Rico, T.; Sosa, S.; Oaxaca, E.; Vizcaino-Castillo, A.; Caballero, M.L.; Martinez, I. Mexican Trypanosoma cruzi (TCI) Strains with different degrees of virulence induce diverse humoral and cellular immune responses in a murine experimental infection model. J. Biomed. Biotechnol. 2010, 2010, 890672. [Google Scholar] [CrossRef] [Green Version]
- da Costa, T.A.; Silva, M.V.; Mendes, M.T.; Carvalho-Costa, T.M.; Batista, L.R.; Lages-Silva, E.; Rodrigues, V.; Oliveira, C.J.; Ramirez, L.E. Immunomodulation by Trypanosoma cruzi: Toward understanding the association of dendritic cells with infecting TcI and TcII populations. J. Immunol. Res. 2014, 2014, 962047. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, C.G.; Carvalho Costa, T.M.; Desiderio, C.S.; Ferreira, P.T.M.; Silva, M.O.; Hernandez, C.G.; Santos, M.M.; Trevisan, R.O.; Bovi, W.G.; Rodrigues, V.; et al. Trypanosoma cruzi Mexican Strains Differentially Modulate Surface Markers and Cytokine Production in Bone Marrow-Derived Dendritic Cells from C57BL/6 and BALB/c Mice. Mediat. Inflamm. 2019, 2019, 7214798. [Google Scholar] [CrossRef]
- Ruiz, R.C.; Favoreto, S., Jr.; Dorta, M.L.; Oshiro, M.E.; Ferreira, A.T.; Manque, P.M.; Yoshida, N. Infectivity of Trypanosoma cruzi strains is associated with differential expression of surface glycoproteins with differential Ca2+ signalling activity. Biochem. J. 1998, 330 Pt 1, 505–511. [Google Scholar] [CrossRef]
- Ferreira, B.L.; Ferreira, E.R.; de Brito, M.V.; Salu, B.R.; Oliva, M.L.V.; Mortara, R.A.; Orikaza, C.M. BALB/c and C57BL/6 Mice Cytokine Responses to Trypanosoma cruzi Infection Are Independent of Parasite Strain Infectivity. Front. Microbiol. 2018, 9, 553. [Google Scholar] [CrossRef]
- Huynh, L.; Kusnadi, A.; Park, S.H.; Murata, K.; Park-Min, K.H.; Ivashkiv, L.B. Opposing regulation of the late phase TNF response by mTORC1-IL-10 signaling and hypoxia in human macrophages. Sci. Rep. 2016, 6, 31959. [Google Scholar] [CrossRef] [Green Version]
- Wanidworanun, C.; Strober, W. Predominant role of tumor necrosis factor-alpha in human monocyte IL-10 synthesis. J. Immunol. 1993, 151, 6853–6861. [Google Scholar] [PubMed]
- Alba Soto, C.D.; Solana, M.E.; Poncini, C.V.; Pino-Martinez, A.M.; Tekiel, V.; Gonzalez-Cappa, S.M. Dendritic cells devoid of IL-10 induce protective immunity against the protozoan parasite Trypanosoma cruzi. Vaccine 2010, 28, 7407–7413. [Google Scholar] [CrossRef] [PubMed]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef]
- Cerban, F.M.; Stempin, C.C.; Volpini, X.; Carrera Silva, E.A.; Gea, S.; Motran, C.C. Signaling pathways that regulate Trypanosoma cruzi infection and immune response. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165707. [Google Scholar] [CrossRef] [PubMed]
- Noel, W.; Raes, G.; Hassanzadeh Ghassabeh, G.; De Baetselier, P.; Beschin, A. Alternatively activated macrophages during parasite infections. Trends Parasitol. 2004, 20, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Cabral-Piccin, M.P.; Guillermo, L.V.; Vellozo, N.S.; Filardy, A.A.; Pereira-Marques, S.T.; Rigoni, T.S.; Pereira-Manfro, W.F.; DosReis, G.A.; Lopes, M.F. Apoptotic CD8 T-lymphocytes disable macrophage-mediated immunity to Trypanosoma cruzi infection. Cell Death Dis 2016, 7, e2232. [Google Scholar] [CrossRef] [Green Version]
- Ponce, N.E.; Sanmarco, L.M.; Eberhardt, N.; Garcia, M.C.; Rivarola, H.W.; Cano, R.C.; Aoki, M.P. CD73 Inhibition Shifts Cardiac Macrophage Polarization toward a Microbicidal Phenotype and Ameliorates the Outcome of Experimental Chagas Cardiomyopathy. J. Immunol. 2016, 197, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Rojas Marquez, J.D.; Ana, Y.; Baigorri, R.E.; Stempin, C.C.; Cerban, F.M. Mammalian Target of Rapamycin Inhibition in Trypanosoma cruzi-Infected Macrophages Leads to an Intracellular Profile That Is Detrimental for Infection. Front. Immunol. 2018, 9, 313. [Google Scholar] [CrossRef] [Green Version]
- Monteón, V.M.; Furuzawa-Carballeda, J.; Alejandre-Aguilar, R.; Aranda-Fraustro, A.; Rosales-Encina, J.L.; Reyes, P.A. American trypanosomosis: In situand generalized features of parasitism and inflammation kinetics in a murine model. Exp. Parasitol. 1996, 83, 267–274. [Google Scholar] [CrossRef]
- Gil-Jaramillo, N.; Motta, F.N.; Favali, C.B.; Bastos, I.; Santana, J.M. Dendritic Cells: A Double-Edged Sword in Immune Responses during Chagas Disease. Front. Microbiol. 2016, 7, 1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-Buelvas, D.M.; Estrada-Muñiz, E.; Rodríguez-Sosa, M.; Shibayama, M.; Vega, L. Increased heart fibrosis and acute infection in a murine Chagas disease model associated with organophosphorus pesticide metabolite exposure. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tomioka, H.; Tatano, Y.; Maw, W.W.; Sano, C.; Kanehiro, Y.; Shimizu, T. Characteristics of suppressor macrophages induced by mycobacterial and protozoal infections in relation to alternatively activated M2 macrophages. Clin. Dev. Immunol. 2012, 2012, 635451. [Google Scholar] [CrossRef] [PubMed]
- Haloul, M.; Oliveira, E.R.A.; Kader, M.; Wells, J.Z.; Tominello, T.R.; El Andaloussi, A.; Yates, C.C.; Ismail, N. mTORC1-mediated polarization of M1 macrophages and their accumulation in the liver correlate with immunopathology in fatal ehrlichiosis. Sci. Rep. 2019, 9, 14050. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BALB/c | C57BL/6 | |||
|---|---|---|---|---|
| Organ 1 | Non-infected | 36 dpi 2 | Non infected | 36 dpi |
| Spleen | 0.096 ± 0.01 | 2.97 ± 0.21 * | 0.098 ± 0.03 | 2.40 ± 0.50 * |
| Liver | 0.87 ± 0.13 | 7.32 ± 0.19 * | 0.97 ± 0.15 | 6.30 ± 0.20 * |
| Heart | 0.126 ± 0.01 | 0.66 ± 0.06 | 0.14 ± 0.03 | 0.60 ± 0.10 |
| Lymph node | 0.092 ± 0.14 | 0.06 ± 0.00 * | 0.098 ± 0.24 | 0.06 ± 0.01 * |
| Body weight (g) | 17.38 ± 0.94 | 17.56 ± 0.15 | 18.43 ± 0.29 | 18.70 ± 0.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina-Buelvas, D.M.; Rodríguez-Sosa, M.; Vega, L. Characterisation of Macrophage Polarisation in Mice Infected with Ninoa Strain of Trypanosoma cruzi. Pathogens 2021, 10, 1444. https://doi.org/10.3390/pathogens10111444
Medina-Buelvas DM, Rodríguez-Sosa M, Vega L. Characterisation of Macrophage Polarisation in Mice Infected with Ninoa Strain of Trypanosoma cruzi. Pathogens. 2021; 10(11):1444. https://doi.org/10.3390/pathogens10111444
Chicago/Turabian StyleMedina-Buelvas, Dunia M., Miriam Rodríguez-Sosa, and Libia Vega. 2021. "Characterisation of Macrophage Polarisation in Mice Infected with Ninoa Strain of Trypanosoma cruzi" Pathogens 10, no. 11: 1444. https://doi.org/10.3390/pathogens10111444
APA StyleMedina-Buelvas, D. M., Rodríguez-Sosa, M., & Vega, L. (2021). Characterisation of Macrophage Polarisation in Mice Infected with Ninoa Strain of Trypanosoma cruzi. Pathogens, 10(11), 1444. https://doi.org/10.3390/pathogens10111444