
Tick Immunobiology and Extracellular Traps: An Integrative Vision to Control of Vectors

 ,
,  ,
, 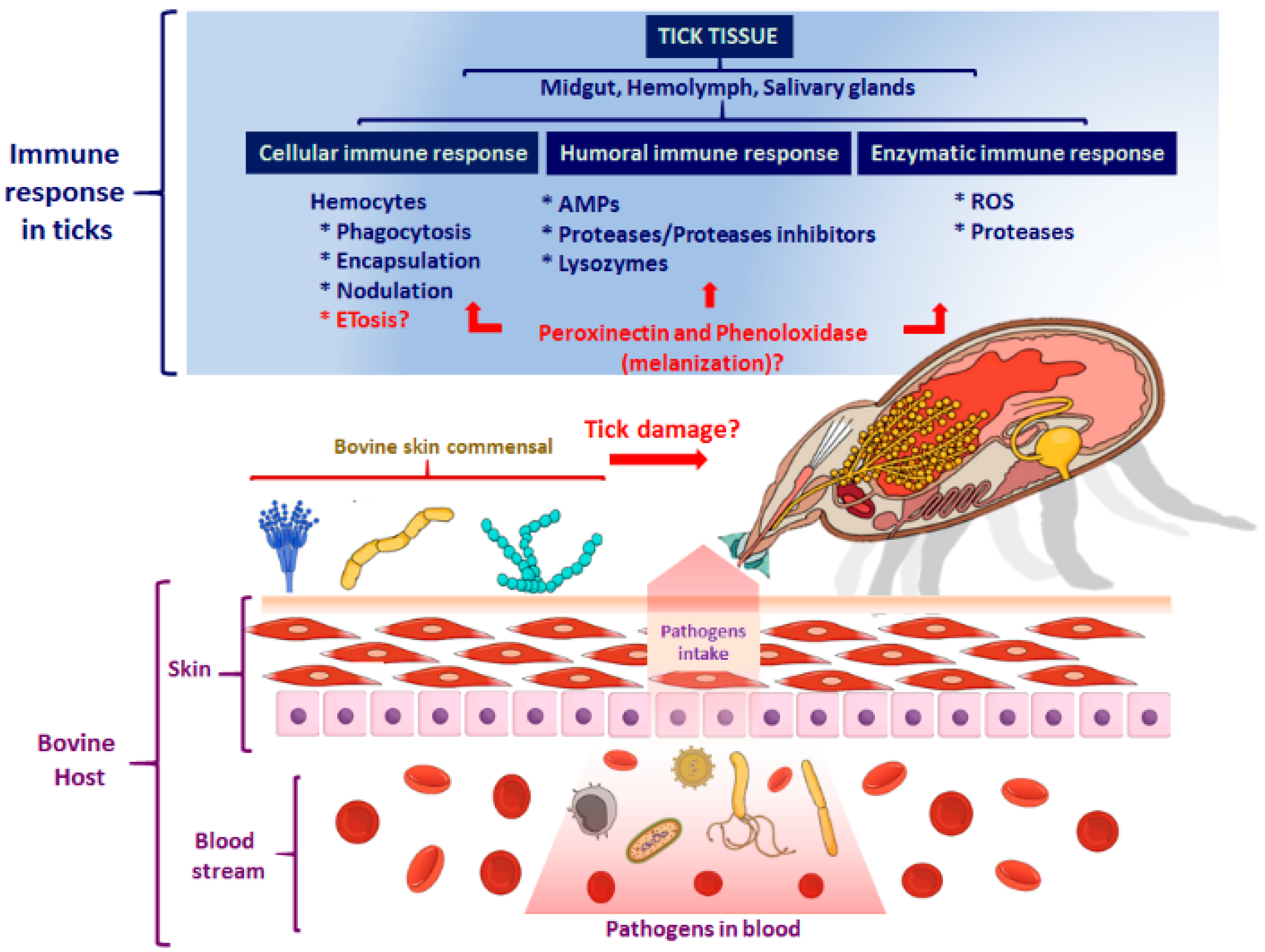{kind=link}
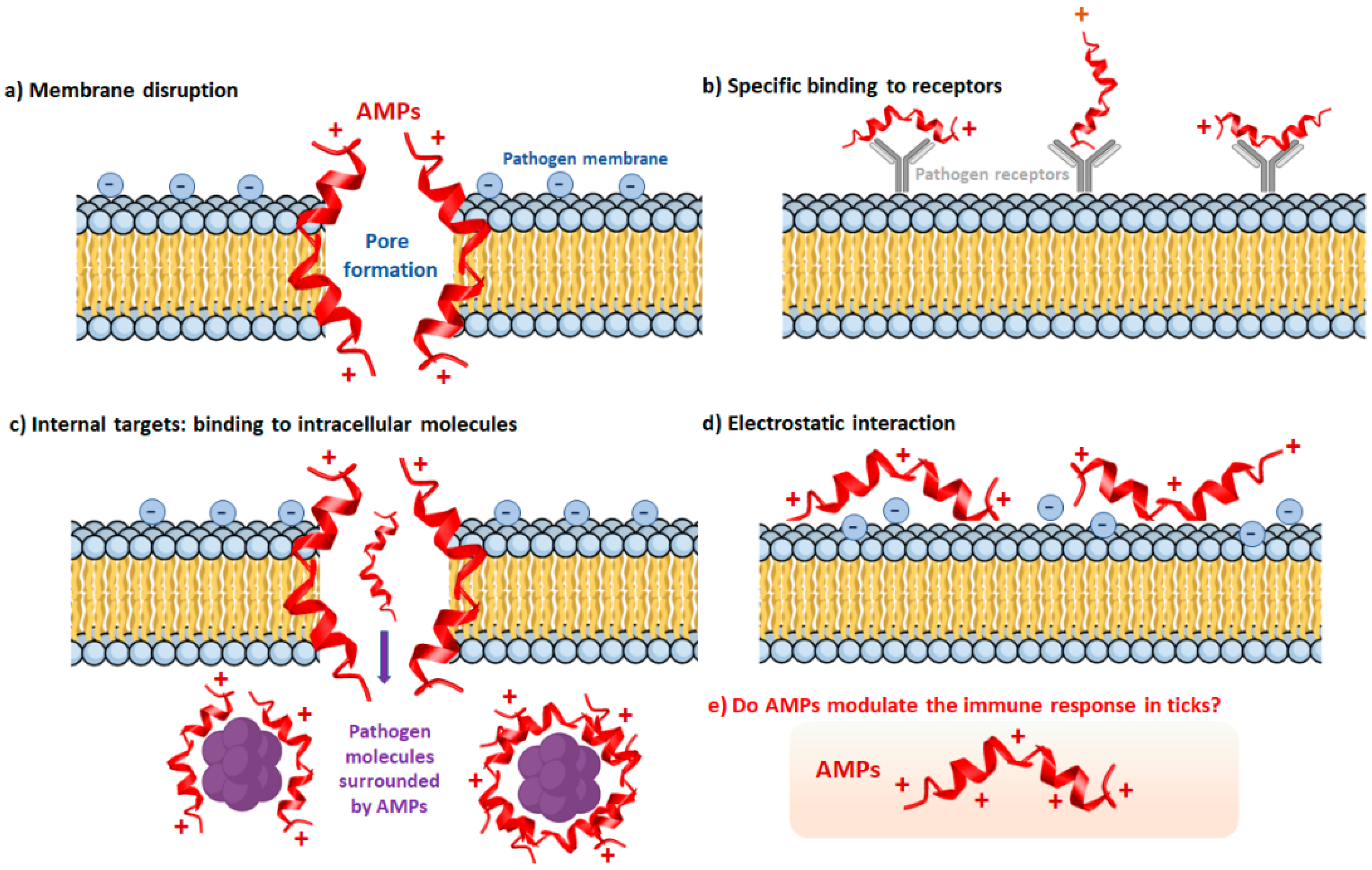{kind=link}
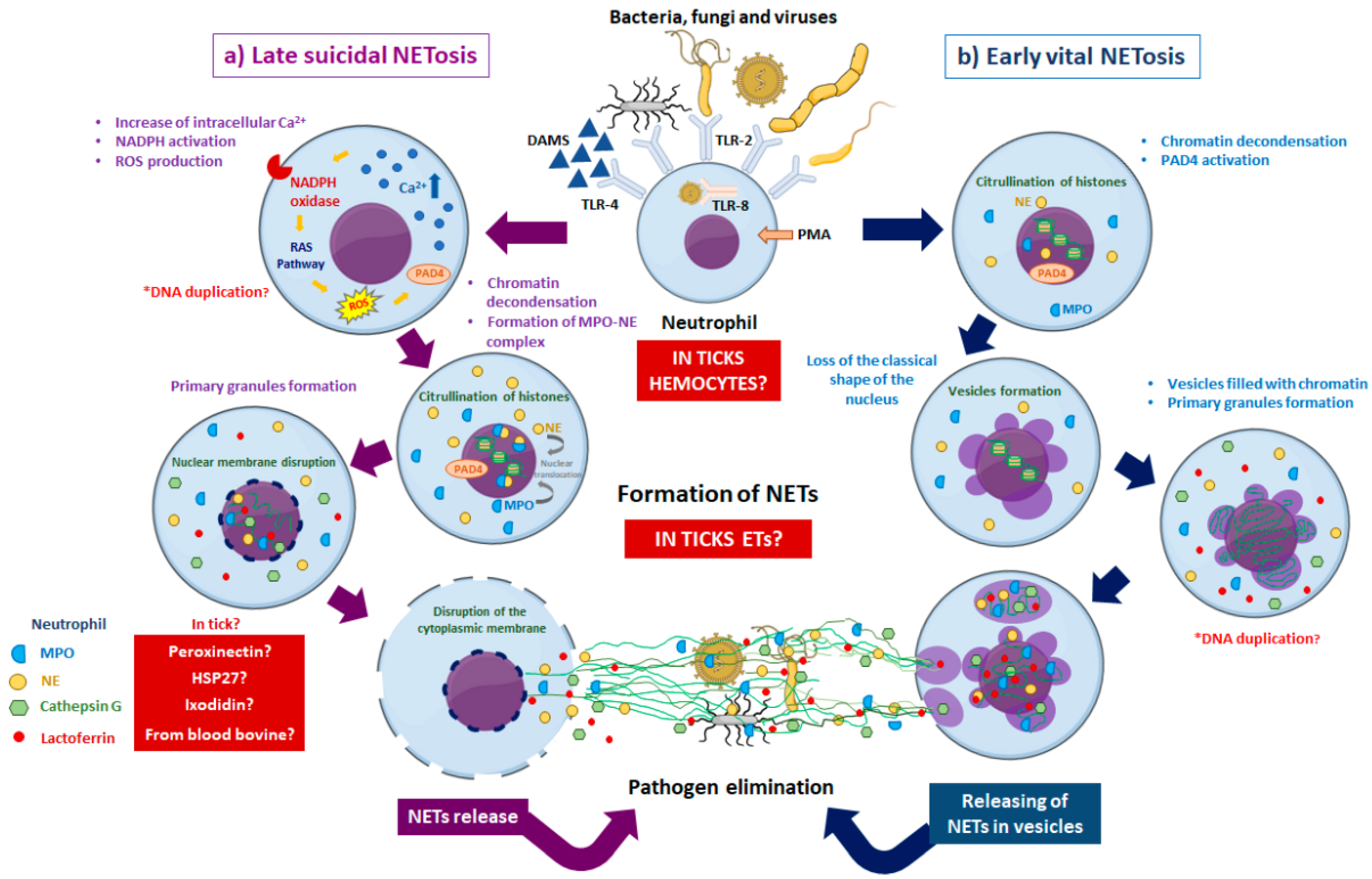{kind=link}
Abstract
:1. Introduction
2. Physical Barriers and Tissues
3. The Tick Immunobiology
3.1. Tick Signaling Pathways in Immune Response
3.1.1. Toll Pathway
3.1.2. Immune Deficiency (IMD) Pathway
3.1.3. Janus Kinase/Signal Transducer and Activator of Transcription (JAK/STAT) Pathway
3.1.4. RNA Interference (RNAi) Pathway
3.2. The Cellular Immune Response
4. The Humoral Immune Response of Ticks
The Tick Repertoire of Humoral Factors
5. Reactive Oxygen Species and Oxidative Stress
6. Do Ticks Have Extracellular Trap Formation?
7. A Comparison of Known, Proposed, and Potential Molecules: Towards an Integrative Vision to Vector Control
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ristic, M. Anaplasmosis. In Diseases of Cattle in the Tropics Economic and Zoonotic Relevance, 1st ed.; Ristic, M., McIntyre, I., Eds.; Martinus Nijhoff: The Hague, The Netherlands, 1981; pp. 327–344. [Google Scholar]
- Betancurt-Hurtado, O.; Giraldo-Ríos, C. Economic and health impact of the ticks in production animals. In Muhammad Abubakar; Perera, P.K., Ed.; Economic and health impact of the ticks in production animals, ticks and tick-borne pathogens; InTech Open: London, UK, 2018; Available online: https://www.intechopen.com/books/ticks-and-tick-borne-pathogens/economic-and-health-impact-of-the-ticks-in-production-animals (accessed on 6 September 2021).
- Tabor, A.E.; Ali, A.; Rehman, G.; Garcia, G.R.; Zangirolamo, A.F.; Malardo, T.; Jonsson, N.N. Cattle tick Rhipicephalus microplus-host interface: A review of resistant and susceptible host responses. Front. Cell. Infect. Microbiol. 2017, 7, 506. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Azhahianambi, P.; Yadav, M.P. Upcoming and future strategies of tick control: A review. J. Vector Borne Dis. 2007, 44, 79–89. [Google Scholar] [PubMed]
- Maritz, C.; Louw, A.; Gothe, R.; Neitz, A. Neuropathogenic properties of Argas (Persicargas) walkerae larval homogenates. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 128, 233–239. [Google Scholar] [CrossRef]
- Hajdušek, O.; Šíma, R.; Ayllón, N.; Jalovecká, M.; Perner, J.; De La Fuente, J.; Kopáček, P. Interaction of the tick immune system with transmitted pathogens. Front. Cell. Infect. Microbiol. 2013, 3, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Kunz, S.; Kemp, D. Insecticides and acaricides: Resistance and environmental impact. Rev. Sci. Tech. 1994, 13, 1249–1286. [Google Scholar] [CrossRef] [PubMed]
- Abbas, R.Z.; Zaman, M.A.; Colwell, D.; Gilleard, J.; Iqbal, Z. Acaricide resistance in cattle ticks and approaches to its management: The state of play. Vet. Parasitol. 2014, 203, 6–20. [Google Scholar] [CrossRef]
- George, J.E.; Pound, J.M.; Davey, R.B. Chemical control of ticks on cattle and the resistance of these parasites to acaricides. Parasitology 2004, 129, S353–S366. [Google Scholar] [CrossRef] [PubMed]
- Kopácek, P.; Hajdusek, O.; Buresová, V.; Daffre, S. Tick innate immunity. Adv. Exp. Med. Biol. 2010, 708, 137–162. [Google Scholar]
- Hackman, R.H. Structure and function in tick cuticle. Annu. Rev. Entomol. 1982, 27, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Kariu, T.; Smith, A.; Yang, X.; Pal, U. A chitin deacetylase-like protein is a predominant constituent of tick peritrophic membrane that influences the persistence of Lyme disease pathogens within the vector. PLoS ONE 2013, 8, e78376. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Mueller, C.R.; Fuchs, J.F.; McElroy, K.; Wessely, V.; Higgs, S.; Christensen, B.M. Evaluation of the function of a type I peritrophic matrix as a physical barrier for midgut epithelium invasion by mosquito-borne pathogens in Aedes aegypti. Vector-Borne Zoonotic Dis. 2008, 8, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.-L.; Cheng, T.-Y. A survey of proteins in midgut contents of the tick, Haemaphysalis flava, by proteome and transcriptome analysis. Exp. Appl. Acarol. 2020, 80, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Francischetti, I.M.B.; Sa-Nunes, A.; Mans, B.J.; Santos, I.M.; Ribeiro, J.M.C. The role of saliva in tick feeding. Front. Biosci. 2009, 14, 2051–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Dai, J.; Zhao, Y.O.; Narasimhan, S.; Yang, Y.; Zhang, L.; Fikrig, E. Ixodes scapularis JAK-STAT pathway regulates tick antimicrobial peptides, thereby controlling the agent of human granulocytic anaplasmosis. J. Infect. Dis. 2012, 206, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- Kvell, K.; Cooper, E.; Engelmann, P.; Bovari, J.; Németh, P. Blurring borders: Innate immunity with adaptive features. Clin. Dev. Immunol. 2007, 83671. [Google Scholar] [CrossRef] [Green Version]
- Fogaça, A.C.; Sousa, G.; Pavanelo, D.B.; Esteves, E.; Martins, L.A.; Urbanova, V.; Kopacek, P.; Daffre, S. Tick immune system: What is known, the interconnections, the gaps, and the challenges. Front. Immunol. 2021, 12, 119. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, F.; Miller, R.; Rousseau, M.-E.; Sunkara, S.; Quackenbush, J.; Lee, Y.; Nene, V. BmiGI: A database of cDNAs expressed in Boophilus microplus, the tropical/southern cattle tick. Insect Biochem. Mol. Biol. 2005, 35, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Megy, K.; Emrich, S.J.; Lawson, D.; Campbell, D.; Dialynas, E.; Hughes, D.S.; Koscielny, G.; Louis, C.; MacCallum, R.; Redmond, S.; et al. VectorBase: Improvements to a bioinformatics resource for invertebrate vector genomics. Nucleic Acids Res. 2012, 40, D729–D734. [Google Scholar] [CrossRef]
- Gulia-Nuss, M.; Nuss, A.; Meyer, J.M.; Sonenshine, D.E.; Roe, R.M.; Waterhouse, R.M.; Sattelle, D.B.; De La Fuente, J.; Ribeiro, J.; Megy, K.; et al. Genomic insights into the Ixodes scapularis tick vector of Lyme disease. Nat. Commun. 2016, 7, 10507. [Google Scholar] [CrossRef] [Green Version]
- Rosa, R.D.; Peixoto, J.; Mesquita, R.D.; Kalil, S.P.; Pohl, P.C.; Braz, G.R.; Fogaça, A.; Daffre, S. Exploring the immune signalling pathway-related genes of the cattle tick Rhipicephalus microplus: From molecular characterization to transcriptional profile upon microbial challenge. Dev. Comp. Immunol. 2016, 59, 1–14. [Google Scholar] [CrossRef]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti Toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef]
- Lemaitre, B. The road to Toll. Nat. Rev. Immunol. 2004, 4, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Kleino, A.; Silverman, N. The Drosophila IMD pathway in the activation of the humoral immune response. Dev. Comp. Immunol. 2014, 42, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, W.J.; Jiggins, F.M. Comparative genomics reveals the origins and diversity of arthropod immune systems. Mol. Biol. Evol. 2015, 32, 2111–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narasimhan, S.; Schuijt, T.J.; Abraham, N.M.; Rajeevan, N.; Coumou, J.; Graham, M.; Robson, A.; Wu, M.-J.; Daffre, S.; Hovius, J.W.; et al. Modulation of the tick gut milieu by a secreted tick protein favors Borrelia burgdorferi colonization. Nat. Commun. 2017, 8, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tafesh-Edwards, G.; Eleftherianos, I. JNK signaling in Drosophila immunity and homeostasis. Immunol. Lett. 2020, 226, 7–11. [Google Scholar] [CrossRef]
- Shaw, D.K.; Wang, X.; Brown, L.J.; Chávez, A.S.O.; Reif, K.E.; Smith, A.A.; Scott, A.J.; McClure, E.E.; Boradia, V.M.; Hammond, H.L.; et al. Infection-derived lipids elicit an immune deficiency circuit in arthropods. Nat. Commun. 2017, 8, 14401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dostert, C.; Jouanguy, E.; Irving, P.; Troxler, L.; Galiana-Arnoux, D.; Hetru, C.; Hoffmann, J.A.; Imler, J.-L. The Jak-STAT signaling pathway is required but not sufficient for the antiviral response of Drosophila. Nat. Immunol. 2005, 6, 946–953. [Google Scholar] [CrossRef]
- Shuai, K.; Ziemiecki, A.; Wilks, A.F.; Harpur, A.G.; Sadowski, H.B.; Gilman, M.Z.; Darnell, J.E. Polypeptide signaling to the nucleus through tyrosine phosphorylation of Jak and Stat proteins. Nat. Cell Biol. 1993, 366, 580–583. [Google Scholar] [CrossRef]
- Capelli-Peixoto, J.; Carvalho, D.D.; Johnson, W.C.; Scoles, G.A.; Fogaça, A.C.; Daffre, S.; Ueti, M.W. The transcription factor Relish controls Anaplasma marginale infection in the bovine tick Rhipicephalus microplus. Dev. Comp. Immunol. 2017, 74, 32–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlikow, M.; Goic, B.; Saleh, M.-C. RNAi and antiviral defense in Drosophila: Setting up a systemic immune response. Dev. Comp. Immunol. 2014, 42, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnettler, E.; Tykalová, H.; Watson, M.; Sharma, M.; Sterken, M.; Obbard, D.; Lewis, S.H.; McFarlane, M.; Bell-Sakyi, L.; Barry, G.; et al. Induction and suppression of tick cell antiviral RNAi responses by tick-borne flaviviruses. Nucleic Acids Res. 2014, 42, 9436–9446. [Google Scholar] [CrossRef]
- Blair, C.D. Mosquito RNAi is the major innate immune pathway controlling arbovirus infection and transmission. Future Microbiol. 2011, 6, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asgari, S. Role of microRNAs in arbovirus/vector interactions. Viruses 2014, 6, 3514–3534. [Google Scholar] [CrossRef] [PubMed]
- Budachetri, K.; Karim, S. An insight into the functional role of thioredoxin reductase, a selenoprotein, in maintaining normal native microbiota in the Gulf Coast tick (Amblyomma maculatum). Insect Mol. Biol. 2015, 24, 570–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narasimhan, S.; Sukumaran, B.; Bozdogan, U.; Thomas, V.; Liang, X.; DePonte, K.; Marcantonio, N.; Koski, R.A.; Anderson, J.F.; Kantor, F.; et al. A tick antioxidant facilitates the Lyme disease agent’s successful migration from the mammalian host to the arthropod vector. Cell Host Microbe 2007, 2, 7–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradkar, P.N.; Duchemin, J.-B.; Voysey, R.; Walker, P.J. Dicer-2-dependent activation of Culex vago occurs via the TRAF-Rel2 signaling pathway. PLoS Negl. Trop. Dis. 2014, 8, e2823. [Google Scholar] [CrossRef] [Green Version]
- Lässer, C. Exosomes in diagnostic and therapeutic applications: Biomarker, vaccine and RNA interference delivery vehicle. Expert Opin. Biol. Ther. 2015, 15, 103–117. [Google Scholar] [CrossRef]
- Sonenshine, D.E.; Hynes, W.L. Molecular characterization and related aspects of the innate immune response in ticks. Front. Biosci. 2008, 1, 7046–7063. [Google Scholar] [CrossRef] [Green Version]
- Strand, M.R. The insect cellular immune response. Insect Sci. 2008, 15, 1–14. [Google Scholar] [CrossRef]
- Urbanová, V.; Šíma, R.; Sauman, I.; Hajdusek, O.; Kopáček, P. Thioester-containing proteins of the tick Ixodes ricinus: Gene expression, response to microbial challenge and their role in phagocytosis of the yeast Candida albicans. Dev. Comp. Immunol. 2015, 48, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Eggenberger, L.R.; Lamoreaux, W.J.; Coons, L.B. Hemocytic encapsulation of implants in the tick Dermacentor variabilis. Exp. Appl. Acarol. 1990, 9, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Ceraul, S.M.; Sonenshine, D.E.; Hynes, W. Resistance of the tick Dermacentor variabilis (Acari: Ixodidae) following challenge with the bacterium Escherichia coli (Enterobacteriales: Enterobacteriaceae). J. Med. Entomol. 2002, 39, 376–383. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Fogaça, A.C.; Lorenzini, D.M.; Kaku, L.M.; Esteves, E.; Bulet, P.; Daffre, S. Cysteine-rich antimicrobial peptides of the cattle tick Boophilus microplus: Isolation, structural characterization and tissue expression profile. Dev. Comp. Immunol. 2004, 28, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D. Innate Immunity in Ticks: A review. J. Acarol. Soc. Jpn. 2006, 15, 109–127. [Google Scholar] [CrossRef]
- Cabezas-Cruz, A.; Tonk, M.; Bleackley, M.R.; Valdés, J.J.; Barrero, R.A.; Hernández-Jarguín, A.; Moutailler, S.; Vilcinskas, A.; Richard-Forget, F.; Anderson, M.A.; et al. Antibacterial and antifungal activity of defensins from the Australian paralysis tick, Ixodes holocyclus. Ticks Tick-Borne Dis. 2019, 10, 101269. [Google Scholar] [CrossRef]
- Nakajima, Y.; Naters-Yasui, A.V.D.G.V.; Taylor, D.; Yamakawa, M. Antibacterial peptide defensin is involved in midgut immunity of the soft tick, Ornithodoros moubata. Insect Mol. Biol. 2002, 11, 611–618. [Google Scholar] [CrossRef]
- Janeway, C.A.J.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [Green Version]
- Jiravanichpaisal, P.; Lee, B.L.; Söderhäll, K. Cell-mediated immunity in arthropods: Hematopoiesis, coagulation, melanization and opsonization. Immunobiology 2006, 211, 213–236. [Google Scholar] [CrossRef]
- Valenzuela, J.G.; Francischetti, I.M.B.; Pham, V.M.; Garfield, M.K.; Mather, T.N.; Ribeiro, J.M.C. Exploring the sialome of the tick Ixodes scapularis. J. Exp. Biol. 2002, 205, 2843–2864. [Google Scholar] [CrossRef]
- Blisnick, A.A.; Foulon, T.; Bonnet, S.I. Serine protease inhibitors in ticks: An overview of their role in tick biology and tick-borne pathogen transmission. Front. Cell. Infect. Microbiol. 2017, 7, 199. [Google Scholar] [CrossRef] [Green Version]
- Saravanan, T.; Weise, C.; Sojka, D.; Kopáček, P. Molecular cloning, structure and bait region splice variants of α2-macroglobulin from the soft tick Ornithodoros moubata. Insect Biochem. Mol. Biol. 2003, 33, 841–851. [Google Scholar] [CrossRef]
- Pereira, L.S.; Oliveira, P.; Barja-Fidalgo, C.; Daffre, S. Production of reactive oxygen species by hemocytes from the cattle tick Boophilus microplus. Exp. Parasitol. 2001, 99, 66–72. [Google Scholar] [CrossRef]
- Aguilar-Díaz, H.; Esquivel-Velázquez, M.; Quiroz-Castañeda, R.E.; Miranda-Miranda, E.; Conde-Baeye, R.J.P.; Cobaxín-Cárdenas, M.; Ostoa-Saloma, P.; Cossio-Bayugar, R. Comparative hemolymph proteomic and enzymatic analyses of two strains of Rhipicephalus (Boophilus) microplus ticks resistant and susceptible to ixodicides. BioMed Res. Int. 2018, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabadin, G.A.; Xavier, M.A.; Da Silva Vaz, I. Control of redox homeostasis in tick blood feeding. Acta Sci. Vet. 2019, 47, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, E.P.; Talactac, M.R.; Fujisaki, K.; Tanaka, T. The case for oxidative stress molecule involvement in the tick-pathogen interactions -an omics approach. Dev. Comp. Immunol. 2019, 100, 103409. [Google Scholar] [CrossRef] [PubMed]
- Villar, M.; Ayllón, N.; Alberdi, P.; Moreno, A.; Moreno, M.; Tobes, R.; Mateos-Hernández, L.; Weisheit, S.; Bell-Sakyi, L.; de la Fuente, J. Integrated metabolomics, transcriptomics and proteomics identifies metabolic pathways affected by Anaplasma phagocytophilum infection in tick cells. Mol. Cell. Proteom. 2015, 14, 3154–3172. [Google Scholar] [CrossRef] [Green Version]
- Busby, A.T.; Ayllón, N.; Kocan, K.M.; Blouin, E.F.; DE LA Fuente, G.; Galindo, R.C.; Villar, M. Expression of heat shock proteins and subolesin affects stress responses, Anaplasma phagocytophilum infection and questing behaviour in the tick, Ixodes scapularis. Med. Vet. Entomol. 2012, 26, 92–102. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente, J.; Blouin, E.F.; Manzano-Roman, R.; Naranjo, V.; Almazán, C.; de la Lastra, J.M.P.; Zivkovic, Z.; Jongejan, F.; Kocan, K.M. Functional genomic studies of tick cells in response to infection with the cattle pathogen, Anaplasma marginale. Genomics 2007, 90, 712–722. [Google Scholar] [CrossRef] [Green Version]
- De Vito, P. The sodium/hydrogen exchanger: A possible mediator of immunity. Cell Immunol. 2006, 240, 69–85. [Google Scholar] [CrossRef]
- Kocan, K.M.; Zivkovic, Z.; Blouin, E.F.; Naranjo, V.; Almazán, C.; Mitra, R.; De La Fuente, J. Silencing of genes involved in Anaplasma marginale-tick interactions affects the pathogen developmental cycle in Dermacentor variabilis. BMC Dev. Biol. 2009, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Zychlinsky, A. Neutrophil extracellular traps: Is immunity the second function of chromatin? J. Cell Biol. 2012, 198, 773–783. [Google Scholar] [CrossRef] [Green Version]
- Romero, A.; Novoa, B.; Figueras, A. Extracellular traps (ETosis) can be activated through NADPH-dependent and -independent mechanisms in bivalve mollusks. Dev. Comp. Immunol. 2020, 106, 103585. [Google Scholar] [CrossRef] [PubMed]
- De Bont, C.M.; Koopman, W.J.H.; Boelens, W.C.; Pruijn, G.J.M. Stimulus-dependent chromatin dynamics, citrullination, calcium signalling and ROS production during NET formation. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865 Pt A, 1621–1629. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Díaz-Godínez, C.; Carrero, J.C. The state of art of neutrophil extracellular traps in protozoan and helminthic infections. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Hoppenbrouwers, T.; Autar, A.S.A.; Sultan, A.R.; Abraham, T.E.; Van Cappellen, W.A.; Houtsmuller, A.B.; Van Wamel, W.J.B.; Van Beusekom, H.M.M.; Van Neck, J.W.; De Maat, M.P.M. In vitro induction of NETosis: Comprehensive live imaging comparison and systematic review. PLoS ONE 2017, 12, e0176472. [Google Scholar] [CrossRef] [Green Version]
- Azzouz, D.; Khan, M.A.; Sweezey, N.; Palaniyar, N. Two-in-one: UV radiation simultaneously induces apoptosis and NETosis. Cell Death Discov. 2018, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Robb, C.T.; Dyrynda, E.A.; Gray, R.; Rossi, A.G.; Smith, V.J. Invertebrate extracellular phagocyte traps show that chromatin is an ancient defence weapon. Nat. Commun. 2014, 5, 4627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, G. NETosis; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2019; p. 200. [Google Scholar]
- Von Köckritz-Blickwede, M.; Goldmann, O.; Thulin, P.; Heinemann, K.; Norrby-Teglund, A.; Rohde, M.; Medina, E. Phagocytosis-independent antimicrobial activity of mast cells by means of extracellular trap formation. Blood 2008, 111, 3070–3080. [Google Scholar] [CrossRef]
- Yousefi, S.; Gold, J.A.; Andina, N.; Lee, J.J.; Kelly, A.M.; Kozlowski, E.; Schmid, I.; Straumann, A.; Reichenbach, J.; Gleich, G.J.; et al. Catapult-like release of mitochondrial DNA by eosinophils contributes to antibacterial defense. Nat. Med. 2008, 14, 949–953. [Google Scholar] [CrossRef]
- Poirier, A.C.; Schmitt, P.; Rosa, R.D.; Vanhove, A.S.; Kieffer-Jaquinod, S.; Rubio, T.P.; Charrière, G.M.; Destoumieux-Garzón, D. Antimicrobial histones and DNA traps in invertebrate immunity: Evidences in Crassostrea gigas. J. Biol. Chem. 2014, 289, 24821–24831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, A.; Brogden, G.; Von Köckritz-Blickwede, M. Extracellular Traps: An ancient weapon of multiple kingdoms. Biology 2020, 9, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homa, J. Earthworm coelomocyte extracellular traps: Structural and functional similarities with neutrophil NETs. Cell Tissue Res. 2018, 371, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Koiwai, K.; Alenton, R.R.R.; Kondo, H.; Hirono, I. Extracellular trap formation in kuruma shrimp (Marsupenaeus japonicus) hemocytes is coupled with c-type lysozyme. Fish Shellfish Immunol. 2016, 52, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.; Lind, M.; Holmblad, T.; Thornqvist, P.; Soderhall, K. Peroxinectin, a novel cell adhesion protein from crayfish blood. Biochem. Biophys. Res. Commun. 1995, 216, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Thörnqvist, P.-O.; Johansson, M.W.; Söderhäll, K. Opsonic activity of cell adhesion proteins and β-1,3-glucan binding proteins from two crustaceans. Dev. Comp. Immunol. 1994, 18, 3–12. [Google Scholar] [CrossRef]
- Ng, T.H.; Chang, S.-H.; Wu, M.-H.; Wang, H.-C. Shrimp hemocytes release extracellular traps that kill bacteria. Dev. Comp. Immunol. 2013, 41, 644–651. [Google Scholar] [CrossRef]
- Kalil, S.P.; Da Rosa, R.D.; Capelli-Peixoto, J.; Pohl, P.C.; De Oliveira, P.L.; Fogaça, A.C.; Daffre, S. Immune-related redox metabolism of embryonic cells of the tick Rhipicephalus microplus (BME26) in response to infection with Anaplasma marginale. Parasites Vectors 2017, 10, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klouwens, M.J.; Trentelman, J.J.A.; Wagemakers, A.; Ersoz, J.I.; Bins, A.D.; Hovius, J.W. Tick-tattoo: DNA vaccination against Borrelia burgdorferi or Ixodes scapularis tick proteins. Front. Immunol. 2021, 12, 138. [Google Scholar] [CrossRef] [PubMed]
- Willadsen, P. Vaccination against Ticks and the Control of Ticks and Tick-Borne Disease; Springer: Dordrecht, The Netherlands, 2005; Available online: http://inis.iaea.org/search/search.aspx?orig_q=RN:37064659 (accessed on 9 September 2021).
- Mitchell, R.D., III; Sonenshine, D.E.; Pérez de León, A.A. Vitellogenin receptor as a target for tick control: A Mini-Review. Front. Physiol. 2019, 10, 618. [Google Scholar] [CrossRef]
- Galay, R.L.; Miyata, T.; Umemiya-Shirafuji, R.; Maeda, H.; Kusakisako, K.; Tsuji, N.; Mochizuki, M.; Fujisaki, K.; Tanaka, T. Evaluation and comparison of the potential of two ferritins as anti-tick vaccines against Haemaphysalis longicornis. Parasites Vectors 2014, 7, 482. [Google Scholar] [CrossRef]
- Zhou, J.; Ueda, M.; Umemiya, R.; Battsetseg, B.; Boldbaatar, D.; Xuan, X.; Fujisaki, K. A secreted cystatin from the tick Haemaphysalis longicornis and its distinct expression patterns in relation to innate immunity. Insect Biochem. Mol. Biol. 2006, 36, 527–535. [Google Scholar] [CrossRef]
- Parizi, L.F.; Rangel, C.K.; Sabadin, G.A.; Saggin, B.F.; Kiio, I.; Xavier, M.A.; Matos, R.D.S.; Camargo-Mathias, M.I.; Seixas, A.; Konnai, S.; et al. Rhipicephalus microplus cystatin as a potential cross-protective tick vaccine against Rhipicephalus appendiculatus. Ticks Tick-Borne Dis. 2020, 11, 101378. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, N.; Miyoshi, T.; Battsetseg, B.; Matsuo, T.; Xuan, X.; Fujisaki, K. A Cysteine protease is critical for Babesia spp. transmission in Haemaphysalis ticks. PLoS Pathog. 2008, 4, e1000062. [Google Scholar] [CrossRef] [Green Version]
- Galay, R.L.; Aung, K.M.; Umemiya-Shirafuji, R.; Maeda, H.; Matsuo, T.; Kawaguchi, H.; Miyoshi, N.; Suzuki, H.; Xuan, X.; Mochizuki, M.; et al. Multiple ferritins are vital to successful blood feeding and reproduction of the hard tick Haemaphysalis longicornis. J. Exp. Biol. 2013, 216 Pt 10, 1905–1915. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, C.; McMaster, W.R.; Girard, D.; Descoteaux, A. Leishmania donovani promastigotes evade the antimicrobial activity of neutrophil extracellular traps. J. Immunol. 2010, 185, 4319–4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, B.M.; Fisher, B.J.; Kraskauskas, D.; Farkas, D.; Brophy, D.F.; Fowler, A.A.; Natarajan, R. Vitamin C: A novel regulator of neutrophil extracellular trap formation. Nutrients 2013, 5, 3131–3150. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Camarillo, S.D.; Quiroz-Castañeda, R.E.; Aguilar-Díaz, H.; Vara-Pastrana, J.E.; Pescador-Pérez, D.; Amaro-Estrada, I.; Martínez-Ocampo, F. Immunoinformatic analysis to identify proteins to be used as potential targets to control bovine Anaplasmosis. Int. J. Microbiol. 2020, 2020, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.H.G.; Contet, A.; Krueger, K. Arthropod innate immune systems and vector-borne diseases. Biochemistry 2017, 56, 907–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar-Díaz, H.; Quiroz-Castañeda, R.E.; Salazar-Morales, K.; Cossío-Bayúgar, R.; Miranda-Miranda, E. Tick Immunobiology and Extracellular Traps: An Integrative Vision to Control of Vectors. Pathogens 2021, 10, 1511. https://doi.org/10.3390/pathogens10111511
Aguilar-Díaz H, Quiroz-Castañeda RE, Salazar-Morales K, Cossío-Bayúgar R, Miranda-Miranda E. Tick Immunobiology and Extracellular Traps: An Integrative Vision to Control of Vectors. Pathogens. 2021; 10(11):1511. https://doi.org/10.3390/pathogens10111511
Chicago/Turabian StyleAguilar-Díaz, Hugo, Rosa Estela Quiroz-Castañeda, Karina Salazar-Morales, Raquel Cossío-Bayúgar, and Estefan Miranda-Miranda. 2021. "Tick Immunobiology and Extracellular Traps: An Integrative Vision to Control of Vectors" Pathogens 10, no. 11: 1511. https://doi.org/10.3390/pathogens10111511
APA StyleAguilar-Díaz, H., Quiroz-Castañeda, R. E., Salazar-Morales, K., Cossío-Bayúgar, R., & Miranda-Miranda, E. (2021). Tick Immunobiology and Extracellular Traps: An Integrative Vision to Control of Vectors. Pathogens, 10(11), 1511. https://doi.org/10.3390/pathogens10111511